イヌにおけるインターロイキン 1 β 誘導性細胞応答と シグナル伝達
103
0
0
全文
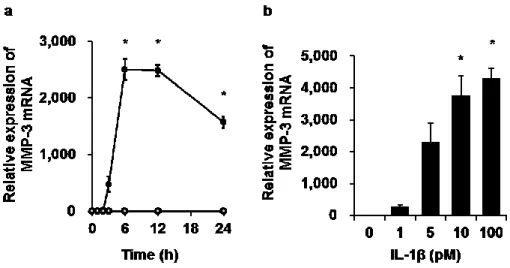
(2)
(3)
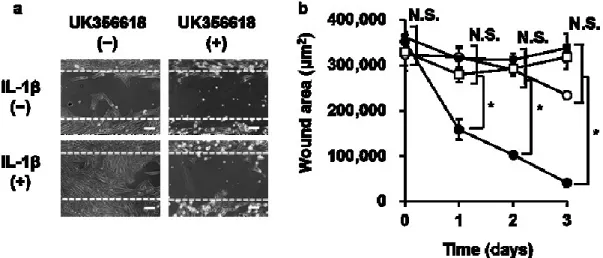
(4)
(5)
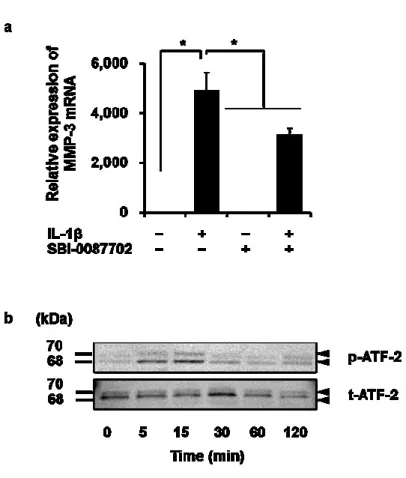
(6)
(7)
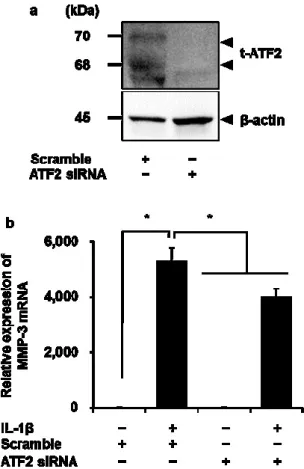
(8)
(9)
(10)
(11)
(12)
(13)
(14)
(15)
(16)
(17)
(18)
(19)
(20)
(21)
(22)
(23)
(24)
(25)
(26)
(27)
(29)
(30)
(31)
(32)
(33)
(34)
(35)
(36)
(37)
(38)
(39)
(40)
(41)
(42)
(43)
(44)
(45)
(46)
(47)
(48)
(49)
図




+7


Outline
関連したドキュメント
However, VPA prevented the morphological changes characteristic for activation and inhibited the expres- sion of collagen type 1 α 1 (COL1A1) and TGF- β 1 in activated LI90 cells
Keywords: homology representation, permutation module, Andre permutations, simsun permutation, tangent and Genocchi
The Beurling-Bj ¨orck space S w , as defined in 2, consists of C ∞ functions such that the functions and their Fourier transform jointly with all their derivatives decay ultrarapidly
As an application, in a neighborhood of a non-degenerate periodic solution a new type of step-dependent, uniquely determined, closed curve is detected for the discrete
Analogs of this theorem were proved by Roitberg for nonregular elliptic boundary- value problems and for general elliptic systems of differential equations, the mod- ified scale of
Then it follows immediately from a suitable version of “Hensel’s Lemma” [cf., e.g., the argument of [4], Lemma 2.1] that S may be obtained, as the notation suggests, as the m A
Definition An embeddable tiled surface is a tiled surface which is actually achieved as the graph of singular leaves of some embedded orientable surface with closed braid
We study the local dimension of the invariant measure for K for special values of β and use the projection to obtain results on the local dimension of the Bernoulli
