EFFECT OF HEAT TREATMENT AND D-GLUCOSAMINE ON
T H
E GERMINATION OF BACILLUS SUBTILIS SPORES.
Koichi OKUTANI and Chihiro NAKAGAWA
Introduction
It has been known that there were many substances which acted a s germinating agents for bacterial spores after heat treatment.") However, i t has not yet been clear whether the heat treatment of spores increases the germination rate or not.
D-glucosamine was found to be a n initiator for the germiation by NELSON e t a1.(2) They also reported that D-glucosamine possibly acted a s an energy source for early stages of the germination.
The present experiment was attempted to analyze quantitatively the effect of D-gluco- samine a s an initiator on the germination of spores.
Materials and Methods
Bacterial strain: Bacillus subtilzs (Marburg 168,thy,- try-) was used for all the studies reported here. ( 3 )
Preparation of spores: Spores were harvested after 48 hrs' incubation a t 32 OC in the sporulation medium of SCHAEFFER. ( 4 )
For maintaining the strain a nutrient agar slant was used.
For the experiments a loopful of the cells grown on a nutrient agar slant was suspended in 10 ml of a sterilyzed distilled water and heated in a water bath a t 80 "C for 20 min. An aliquot of this suspension (1 ml) was inoculated into 10 ml of SCHAEFFER'S medium and incubated a t 32 "C on a shaker for 12 hrs. An aliquot of this culture ( 1 ml) was transferred into 100 ml of the fresh medium and incubated a t 32 OC for 12 hrs. And then finally an aliquot of this culture (1 ml) was again transferred into 100 ml of the fresh medium and incubated a t 32°C for 48 hrs.
The cells were harvested by centrifugation for 20 min a t 900 x g in a refrigerated centrifuge. The pellet was resuspended in distilled water (O.D. =1.5 a t 560 rnp with a Leitz spectrophotometer) and treated with 200 wg/ml of crystalline egg white lysozyme (Sigma) a t 37OC for 30 min, and then 1% sodium lauryl sulfate (SLS). Spores were washed five times with distilled water and kept in distilled water a t 4OC.
The spore preparations showed complete absence of vegetative cells and all of the forms present were refractile spores as revealed by phase-contrast microscopy.
Germination of spores: Germination was carried out by shaking spores a t 37OC in various media.
Results
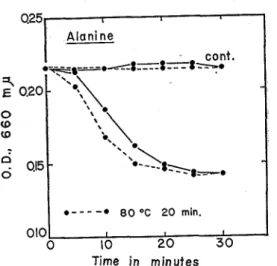
The effect of L-alanine on the germination: The spores which were heated or not heated were incubated in L-alanine solution prepared with distilled water, and the changes in optical density during incubation were observed (Fig. 1)
.
As shown in Fig. 1, during the first 15 min O.D. in L-alanine media decreased sharply. The controls without L-alanine, on the other hand, retained their initial O.D. until 30 min. The germinatin of spores was slightly stimulated by the heat treatment. In the case of L-alanine induced germination, during incubation of the first 15 min, heated spores showed a little more decrease in 0.D than not heated spores.
The patterns of the germination of spores in the incubation medium prepared with M/100 Tris-HC1 buffer (pH6.8) are shown in Fig. 2.
As shown in Fig 2, during the first 15 min, 0. D. decreased sharply also in this medi- um. In the control media without L-alanine 0. D. decreased gradually until 30 min. This result is somewhat different from that illustrated in Fig. 1.
In I.-alanine t Tris buffer systems, the germination was a little more stimulated by heat treatment a s compared with L-alanine f water systems.
0.25 1 t
Alani ne
Fig. 1. Germination induced by L-alanine in distilled water.
The reaction system contained spore susp- ension and 45 mM L-alanine. The control medium contained only spore suspension. An aliquot of the spore suspension was heated for 20 min at 80°C in water. Another aliquot of the suspension was incubated without heat treatment. 010 Ala.
+
M/100 Tris."W
.-
- -
- 0 8 0 OC 20 min. I I 0 10 20 3 0Time i n minutes Time i n minutes
2
Q2;--q-$--*
Fig .2. Germination induced by L-alanine in Tr is-HC1 buffer
.
The reaction syat:m contained spore snspen- sion, 45 mM L-alanine and M/100 Tris-HC1 buffer (pH 6.8). The control system comp- osed of spore suspension and M/100 Tris- HCl buffer (pH 6.8). Spores were treated as in Fig. 1. 0 (0 (D C 0 6 015- 010
The effect of D-glucosamine on the germination : Spores were incubated in D-glucosa - mine solution, and 0. D. changes during incubation were observed (Fig.3).
As shown in Fig. 3, during incubation for 30 min 0. D. in glucosamine media decreased gradually. While, controls without added D-glucosamine retained their initial O.D. until 30 min. Heat activated spores showed less decrease in O.D. than unheated spores. Patt-
y
-
cont.-
\;---..
\
*,
*-GI&,
,,,in.- \ * '%*\$ky----*
I I 0 10 20 30erns of the germination of spores in the glucosamine media prepared with M/100 Tris-HC1 buffer (pH6.8) are shown in Fig. 4.
As shown in Fig 4, in the case of D-glucosamine $ Tris buffer systems O.D. of spore
suspension with or without heat treatment retained their initial values until 30 min, whe- reas O.D. of the suspensions without glucosamine decreased gradually until 30 min. These results suggest that D-glucosamine slightly inhibited the germination of spores in M/100 Tris-HC1 buffer (pH6.8) solution.
l
a - - - , - 0 80°C 2 0 min.Time in minutes
Fig. 3. Germination induced by D-glucosamine in water.
The reaction system contained spore susp- ension and 40 mM D-glucosamine-HC1. The control system contained only spore suspen- sion. Spores were treated as in Fig.1.
0 25 1 I
M/IO Citrate
QIQQ
0 I0 2 0 30
Time in minutes
Fig.5. Effect of pH values on germination. The reaction system contained spore suspen- sion and MI10 citrate buffer. The pH values f o the reaction systems were indicated in the figures. 0.25 I t I G A .t M A 0 0 T r i s .
I
I
.-.--
-
-*
80°C 20 min.I
0101
t I I I 0 10 20 30 ' Time i n minutesFig. 4. Germination induced by D-glucosamine in Tris HC1 buffer.
The reaction system contained spore snsp- ension, 40 mM D-glucosamine-HC1 and M/100 Tris-HCI buffer (pH6.8). The control syst- em contained spore suspension and M/100 Tris-HC1 bnf f er (pH6
.a).
Spores were trea- ted as in Fig. 1The effect of pH on the germination: The 0. D. changes of the spore suspensions in M/10 citr ate buffer duriug incubation are shown in Fig .5. In all the cases, no germinating agents were added.
As shown in Fig. 5, the gradual decre- ases of 0 .D. were observed in all the med
-
ia tested. The rate of the decreases of O.D. under variouspH values tested were not conspicuousDiscussion
In L-alanine medium, heating of spores a t 80°C for 20 min stimulated the germination rate of Baczllus subtzlzs (Marburg 168, thy-, try-).
KADOTA et a l S t s ) studied the fate of heat activation of spores (80°C, 20 min) prior to germination of Bacillus subtzlzs (Marburg, ATCC 6051). Their results indicated that heat treatment of spores in L alanine mediurn delayed the germination of spores freshly harv-
ested, but remarkably activated the germination of spores stored a t O°C for more than 66 days.
Though contradictory results were obtained in the present experiments, the present res- ults are not compared directly with those of previous experiments, because the materials and methods employed in the present experiments were somewhat different from those in the previous studies.
In order to maintain the constant pH value during the germination of spores, M/100 Tris-HCl buffer (pH6.8) were used. The L-alanine germination system in M/100 Tris- HCI buffer showed no significant differences in the germination patterns of spores, com- pared with those in systems without Tris-HC1 buffer.
The present data suggested that the germination in the systems without M/100 Tris-HC1 buffer (pH6.8) was stimulated by D-glucosamine. But in these systems, 40 mM of D-glucosamine made the germination mixture to pH 4.5, therefore, the germination pattern in D-glucosamine medium might be due to the low pH value in the germination medium. The fact that in the glucosamine medium prepared with M/100 Tris-HC1 buffer (pH6.8), no germination took place within 30 min may confirm this presumption.
In spite of these data, there is no enough reason to disagree with the possibility that D-glucosamine acts as a germinating agent. NELSON et a1. c 2 ) pointed out that some synthetic reactions may occur relatively early in the germination of spores and these reactions require energy. They discussed the possibility of D-glucosamine induced germ- inations and the utilization of exogenous glucosamine for energy source.
However, the pathway of D-glucosamine metabolism in Bacillus subtilzs during germin
-
ation has not been known.Summary
D-glucosamine was found to act a s a germinating agent for Bacillus subtilis spores. The heat treatment (80°C, 20 min) had a stimulatory effect on the germination of spo- res in L-alanine medium,'and inhibitory effect on the germination in D-glucosamine med- ium.
In germination media bufferized with M/100 Tris-HC1 (pH6.8) L-alanine acted as a germinating substance. However, D-glucosamine did not act as an initiator for the ger- mination. In this case the heat treatment of spores had no effect on the germination.
Some relationships were found between concentration of D-glucosamine and pH values of germination media.
Acknowledgment
The authors wish to express their thanks to Professor Dr. Hajime KADOTA of the Rese- arch Institute for Food Science, Kyoto University, for his valuable discussions and sug- gestion.
References
(1) NELSON D .L. and KORNBERG A. : J . Bzol. Chem., 245 1128-1136(1970). (2) NELSON D.L. and KORNBERG A.: J.Bzol.Chem, 245, 1146-1155(1970)
(3) TERANO H , TANOOKA H. and KADOTA H . : Bzochem Biophys Res Comm , 3 7 , 66-71 (1969). (4) TAKAHASHI I. : J Bacte~zol , 8 9 , 294-298 (1965).
(5) KADOTA H., SHICKI Y .and ISHIDA Y. : Bull. Jap Soc
.
S C Z Fzsh , 3 3 , 758-762(1967).Bacillus subtilis
EF
8%%K
~
I
M
Thn@&@E
2
D--9')v3.3.
Z'JD#$%
@&B-
+)Il=FSD - y j V 3 9
<
YfI$@@@3 D $%?$%j&$J2 L T D.YFm&g
P z j d ~ ~ 6 ~ & 3 I . " , 6 k & K @ ~ & ~ D%?$%j&%Jki
k
zj tkfi;@~aGik~c.r/\zjL-77.sYD.Y~Ht:li;@Lk&~b@W%fi& = k o & s @ T M ,
+jH
t: L -2 Baczllus subtilzs (Marburg, 168, thf,trf)%mb\ft,D - - y ) V 3 9
:
YM, ~ 6 7 5 4 C t D ~ ~ D $ % ~ & @ % ~ - % Z 2 d : ~ 8 & b h 7 F i o LfInLja;dIb, L - 7 7b h ~ @ ? & J 3 6 & l & $ i j & @ K a,;sB?$D@@B?jW;z& b h y , %KpH&%%D $5 zj t k &ii8& 6;kko
-3, D - y j v ~ 9 ? Ya$k,M/lOOTris-HCl buffer (pH 623) @-CM%%%j@#J L TD%%&ZZ& b h ? ,
~3Dfin#%%j@@%%%~~K~Gs%E%%&L&b~Z2fi::2&5;hk0 D - Y P ~ Y
s
'/(40mM)7fd&@&, 37'CDljijG&?i$&$TMpH4.5&%f c 2 ; 5 : 3 & b h , ~~KBB@LT@@~~D%S.Y~HK%~~PR$GYR%DPH.W~D~~
%K9b\ T Z , 3 DR3J &fi ja; 9 k o