近畿大学学術情報リポジトリ
20
0
0
全文
(2) Doctoral. Two maintaining. distinct. Dissertation. factors,. nuclear constitutive. promoter. angiotensinogen. Seijiro. Graduate. School. First. Department. of. RBF-. activity. of. 1, human. gene. Shimada. of Medicine,. Course. Sp 1 and. of. Internal. November. Kinki. University. Medicine. (Director 1995. : Prof.. Ryo. Katori).
(3) Two. distinct. nuclear. maintaining. factors,. constitutive. Sp 1. promoter. activity. angiotensinogen. Seijiro. First. RBF-. 1 ,. of. human. gene. Shimada. Department. of Internal. Kinki University (Director. and. Medicine,. School of Medicine. : Prof.. Ryo. Katori). ABSTRACT Angiotensinogen, blood into. pressure. the protein. precursor. and electrolyte. the circulation.. The. balance,. its own expression. regulation. human. of the. promoter various. region lengths. transiently. transfected. nucleotide. positions. full extent. of parental. using. this segment. complexes either. could. experiment tensinogen. maintaining. has not. plasmids. gene. been. that contain. reported.. promoter. activity.. the formation. of DNA/HepG. by competing. recognition. GC box-binding. factor,. regulating that. the constitutive. mobility. region. the CAT activity. suggest. promoter. that. activity. gene is characterized. (CAT). half. words. : Human. Angiotensinogen,. Gene. at. contain. These. those. of. characterized. (Rb) gene. An in vivo competition from. the cotransfected. angio-. RBF- 1 binding sites or containing. Sp 1 and RBF- 1 play an important of the human. regulation,. ( EMSA ). sequences. derived. as the second. --1—. located. complexes.. angiotensinogen. target. Sp 1 , RBF-. for RBF- 1 beside. 1 , HepG. role in. gene and. gene.. Key. In. assay. or RBF- 1 , a recently. containing. with. as much as the. shift. that. its. gene.. 2 nuclear factor. the human retinoblastoma. is reduced by a competitor. sites. These findings. to mediate. An electrophoretic. been. We analyzed. the 5 '-flanking. acetyltransferase. -344 and -313 by was shown. has. mice, but the transcriptional. HepG 2 cells, the only 32-bp DNA region. suggested. the angiotensinogen. angiotensinogen. human hepatoma. identified. promoter. Sp 1 binding. gene. of. by the liver and secreted. chloramphenicol. be dissociated. also. of the human. regulator. to the bacterial. between. factor. produced. in the liver of transgenic. angiotensinogen. Sp 1 , a widely expressed. GC box-binding. region. by utilizing the reporter fused. II which is a critical. is constitutively. 5 '-flanking. found to regulate. of angiotensin. 2. cells. that. the Rb.
(4) Introduction The renin-angiotensin fluid. volume. system. balance. angiotensin. and. electrolyte. II. Angiotensinogen,. liver and secreted. Genetic linkage that. transcriptional. transgenic. angiotensinogen hormones,7-9. studies. these. revealed. example,. a. direct. glucocorticoid Two. the basal. distinct. of an. pressure,. octapeptide. II, is produced cleavage. gene inheriting. by. occurs. the. by renin. high blood pressure. of the angiotensinogen Fukamizu. of which. gene is involved. et a16 have generated carrying. transgenic. either the human renin. is constitutively. to different. cells differentiation,". at the transcription. between. elements. between. in response. cytokine,'. one underlies. adipogenic. expressed. in the. are. the. for. the regulation. rat. activation. of. angiotensinogen. transcription. or in vivo, Several. mouse.. For. element. and. of the rat angiotensinogen mouse. However, less information. of gene. and. enhancer. the induction. of the human. in vitro. inflammation.'. interleukin-I-inducible. necessary. machinery. and. level, in the. the acute-phase. differentiation.'. states. is available gene,. and. angiotensinogen concerning. and the molecular. the pathogenesis. of hyper-. is unclear.. As the first. step to understand. gene, we examined expression.. For. human. identified partially. (CAT). contributes. contain. binding regulating. and EMSA. with. in human. DNA segment. using. These. suggested. in the control. transient. various. hepatoma. transfection. lengths. linked. positions. promoter. activity.. could be dissociated. binding. factor. using. chloramphenicol. ( HepG 2 ). Then. -313 that. An electrophoretic. mobility. of DNA/HepG. 2 nuclear. recognition. sequences (GC-box). 1 (RBF- 1 ), a novel GC-box binding. gene. Further. that. RBF- 1 play an. we. -344 to. multiple GGGCGG sequences. (Rb). —2—. of its constitutive. to. between. by competing. retinoblastoma Sp 1 and. angiotensinogen. experiments. cell line HepG 2. at nucleotide. Sp 1 , a widely expressed. or retinoblastoma. the human. participate. of the human. this DNA region identified the formation. complexes. those of either. factor,. that. to the angiotensinogen. (EMSA). complexes.. promoter. gene. the upstream. factors. regulation. we have performed. angiotensinogen. shift assay. the molecular. the regulatory. this purpose,. acetyltransferase. that. is regulated. interaction. transcriptional. relationship. factor. proteolytic. lines of animals. gene, the latter. regulations. responsive. mRNA during. the. successive. Recently, separate. angiotensinogen. such as steroid. tension. action. blood. liver.'. In addition,. gene.". the. of angiotensin. mechanism. of hypertension.'. mice by cross-mating. gene or human. precursor. where. in regulating. through. of the human angiotensinogen. the basal. hypertensive. a unique. actions. enzyme.'. analyses. in the pathogenesis. different. homeostasis. into the circulation,. and angiotensin-converting. proposed. plays many. in vivo competition important. role. factor. experiments. in maintaining. the.
(5) constitutive. promoter. activity. of the human. angiotensinogen. Materials. gene.. and methods. Materials Restriction. enzymes. Biolabos ; Takara HindIII. linker. Ci/mmol). and. DNA-modifying. Shuzo Co. Ltd, Osaka,. were obtained. was purchased. /mmol). was from DuPont/. Plasmid. constructions. The human. 44) SmaI/HindIII. fragment,. to make. BglII/Smal. (Figure. Products,. 357-bp (-312 to. derived. ( -344 to -313). Competitive. plasmids. previously.''. and DM 8.5 cat, respectively.. DNA fragment. from. into. of pc-Sp 1 and pc-RBF- 1 were constructed. copies of Sp 1 and RBF- 1 binding. elements. (3000. (50-60 mCi. ( CAT). hybrid. -516 to + 44). A 389-bp (-344 to + fragment,. and. 341-bp. sites of pUCSV 0. A 172-bp (-516 to -345 ). into the BglII site of DM 8'. by inserting. DM 7 cat. BglII and. [ r -52p] ATP. positions. into the BglII/HindIII. -32-bpDM 7 . + 32-bpDM 9 was made. England. USA.. + 44 ) Eco81I/HindIII. cat to constract. Japan.. acetyltransferase. from DM 7 cat was inserted. derived. Japan.. New. [14C] Chloramphenicol. Boston,. as described. were subcloned. from. Co. Ltd, Osaka,. 1) : DM 7 cat (nucleotide. were constructed. DM 7.8 cat, DM 8 'cat,. DNA fragment. Tokyo.. promoter-chloramphenicol. (-296 to +44) BlnI/ HincIIII fragment CAT'. Japan,. NEN Research. as follows. purchased. Shuzo Co. Ltd, Osaka,. from Amersham. and DM 9 cat (-243 to +44). were. Japan ; Toyobo. from Takara. angiotensinogen. genes were constructed. enzymes. the. the 32-bp SmaI/Eco81I BglII. site. of DM 9 cat.. by fusing the eight. into the Smal site of pUC 19, respectively.. tandem.
(6) Fig.. 1. Plasmid constructions. Schematic representation of the human angiotensinogen promoter-CAT hybrid genes. Restriction endonuclease cleavage site is indicated as follows: Bg, BglII; Sm, Smal; Ec, Eco81I; Bl, Blni; Hi, HindIII. TATA box is located. Cell culture, Human Tokyo,. transient. hepatoma Japan). One day before dishes.. amino. containing. bovine serum. by Gorman. (Gibco. essential. Laboratories,. medium. (Nissui,. New York,. 1 X 106 cells for HepG 2 cells were plated. was changed. 3 hours. 3 ug of CsCI-purified. for. in minimum. USA ). acids.. 6 hours,. washed. prior. et al." The extracts. by centrifugation.. to transfection.. DNA was added. were prepared. on 60-mm plastic. A calcium. to the cultured. with PBS( —), and incubated. They were collected and their extracts. was removed. -31 to -24.. and CAT assay. 10% fetal. transfection,. cells were incubated. described. positions. cells (HepG 2) were maintained. The cell medium. coprecipitate. medium.. transfection. containing. and non essential. at nucleotide. concentration. —4—. cell line. The. for 36 hours in fresh. by freezing. were heated at 60°C for 10 min, and. The protein. phosphate. was determined. and thawing. as. the precipitate by the Bio-Rad.
(7) protein. assay. kit. The reaction. mixture. contained. mCi / mmol ) of [ "C ] chloramphenicol, technology. was dried,. at 37°C for. and acetylated. derivatives. chloroform/methanol measured. (95 : 5 , v/v). by. an. image. experiments. were performed. Preparation. of nuclear. Nuclear. extracts. ation. was. was removed. using. of ["C] chloramphenicol Tokyo,. Japan ) . All. five times.. protocol. Hela cells and glioblastoma. of Dignam. et al.lB The final protein. T98G concentr. shift assay. using. competitor. (approximately. 0.5 mM dithiothreitol, loaded. and the double-stranded. G-50 column.. (Pharmacia). containing. Five aug of nuclear 5. label. extracts. were. min in the presence. with the end-labeled. or. oligonucleotide reaction. 10 mM Tris-HC1 (pH 7.5), 30 mM KC1, 10% glycerol,. and 0.5 mM phenylmethylsulfonylfluoride. onto a 4 % nondenaturing. made in 0.5 x TBE that. (100. kinase. Unincorporated. on ice for. DNA and then incubated. DNA probes. 1 X 10' cpm, 0.1 to 0.2 ng) at 20°C for 20 min. The binding. out in a solution. was directly. on a Sephadex. 2 itt g of poly (dI-dC). of unlabeled. was carried. were annealed. r-32p] ATP and T4 polynucleotide. by gel filtration. with. performed. a modified. oligonucleotides. ng) were end-labeled. probe. ratios. chloramphenicol. chromatography. ( BAS 2000 ; Fujix,. HepG 2 cells, cervical carcinoma. using. mobility. Single-stranded. absence. at least. system. layer. The solution. 5 to 15 mg/ml.. Electrophoretic. incubated. The results of conversion. analyzer. The labeled. thin. Bio-. extracts. from. cells were prepared. by ascending. LKB. of 150 it 1. The mixture. with cold ethyl acetate.. in 20 ,ul of ethyl acetate.. were separated. ( Pharmacia. in a final volume. 1 hour and then extracted. and the pellet redissolved. pH 7.8, 0.2 g Ci (50-60. acetyl-coenzymeA. Inc.), and 20 or 40 ,ag of cell extract. was incubated. were. 4 mM. 300mM Tris-HC1,. polyacrylamide. had been pre-electrophoresed. The reaction. mixture (20 ,u1). gels containing. 4 % glycerol. for 20 min. After electrophoresis. was. at 130V for 210 min or 225 min at 4 °C, the gels were dried and autoradiographed. with an intensifying. screen.. Oligonucleotides Oligonucleotide oligonucleotide city, CA) competition. DNA. was. DNA synthesizer,. as described analysis. AGG- 3' described. synthesized. on. and purified. by the manufacturer.. were as follows: by Landschulz. a. on OPC columns Sources. the C/EBP. et al.;'. MilliGen /. (Applied. Cyclone" Biosystems,. of the oligonucleotide. binding. site. the HNF- 1 binding. —5—. Bioresearch. plus Foster. sequences. for. 5 '-AATTCAATTGGGCAATC site (nucleotide. positions. -103.
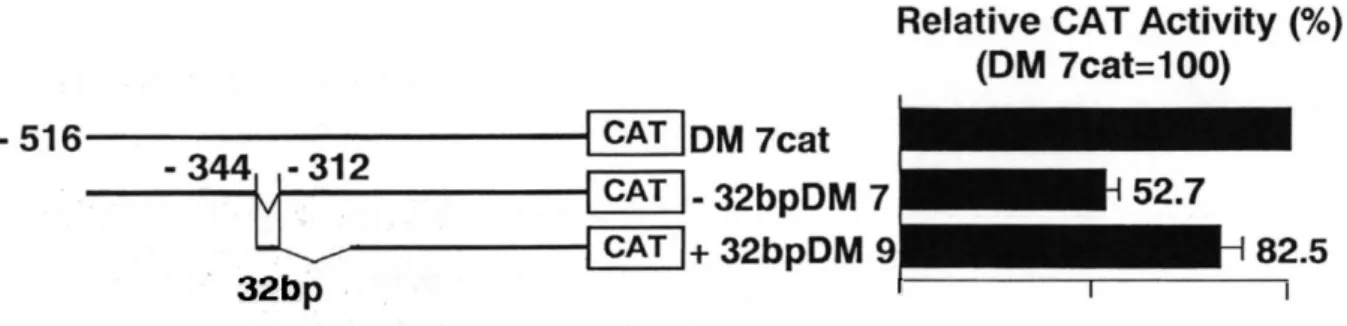
(8) to. -74) ,. 5 '-AACCAAACTGTCAAATATTAACTAAAGGGA-. promoter,'. the HNF- 3 binding site. AG- 3 ' of the mouse transthyretin. promoter,'. GCTGCCGCGGGCGGAAGTTCGAoligonucleotides. containing. (-111 to -85),. GATGAGTCAGCCGGATC-. the. rat. the RBF- 1 binding (Sakai. site (Rb-Sp. sites for transcription. 3 ' were purchased. 1 -wt),. 5 '-A. et al., 1991).22 Double-stranded factors. AP- 1 , 5 '-CTAGT. 3 ', NF- 1 , 5 '-ATTTTGGCTTGAAGCCAATATG-. GATCGATCGGGGCGGGGCGATC-. /9 -fibrinogen. 5 '-GTTGACTAAGTCAATAATCAGAATC. 3' of the Rb gene. the recognition. 3 ' of. from Stratagene. 3 ' and Sp 1 , 5 '(GELSHIFT'. KIT La. Jolla, CA). The 32-bp DNA fragment by the polymerase primer. chain. (-344 to -313) used in EMSA was generated reaction. (-353 to -306),. (PCR). from. using a pair of oligonucleotide. 5 '-ACAgatctGGGGCTGGGTCAGAAG-. 3. and. the DM 7 cat. primers.. The distal. the proximal. primer. ( -306 to 353) , 5 '-GTGGgatccTGAGGCCAACCACC-. 3 ', were synthesized. with a BglII rest-. riction. letter),. The PCR products. site (small. were isolated. letters). from. and a BamHI site (small. 5 % polyacrylamide. respectively.. gels and used for EMSA.. Results Deletion. analysis. Fukamizu human. angiotensinogen. mutation,. To identify. region. significantly DM 7 cat).. reduced Further. The CAT activity region-CAT. promoter. activity. to direct. the upstream. the expression. DNA. the CAT reporter. plasmids. containing. various. angiotensinogen. hand,. positions. deletions. -516 to -345 slightly decreased from. to 50.9% (as compared. (data. -344 to -313 contributed. of the human. angiotensinogen. These to gene.. findings. gene in. the deletion. lengths. of the. 5 '-. in Figure. the promoter. with the parental change. activity. suggested. half as much. as. plasmid,. in CAT activity.. equal level to that found in the 1.3-kb. not shown).. of the. -344 to -313 ( DM 8 'cat). up to -243 did not give a significant. of DM 7 cat was nearly. for. gene into HepG 2 cells. As shown. the only 32-bp DNA deletion activity. region. of the CAT reporter responsible. the promoter. from. that. initially. hybrid construct. bp DNA segment. demonstrated. DNA region. from nucleotide. On the other. promoter. the upstream. of the human. 2 , the deletion. have. gene is able. we constructed. to 83%.. angiotensinogen. and his colleagues. HepG 2 cells.'. flanking. of the human. that the. 5 '-flanking only the 32-. full. extent. of.
(9) Fig.. 2 5 '-Deletion analysis of DM 7 fragment (-516 to +44) of the human angiotensinogen promoter. The TATA box is located at nucleotide positions -31 to -24 and the transcriptional start site is indicated by + 1 . Transfection was performed using 3 lig of reporter plasmid DNA/dish. Cells were harvested 36 h after transfection and aliquots of al extract containing equal amount of total protein (20 or 40 itg) were used for CAT assay. The CAT activity of DM 7 cat is designated as 100 and each value of CAT activity represents the mean ± S.E. for six independent experiments.. To test the functional. importance. tensinogen. promoter. introduced. into HepG 2 cells. The reporter. constructed. activity,. of the 32-bp DNA fragment. by internally. deleting. bp to DM 9 cat, respectively. dramatically and +32-bp. reduced. plasmids,. The CAT activity. to 52.7%, which was. to the level of DM7.8cat. 32-bp DNA element. plays functional. angiotensinogen. gene.. as shown -32-bp. the 32-bp from DM7cat. DM 9 was able to activate. was the similar. the human. the CAT constructs. directed. (-344 to -313) in the angio-. in Figure DM 7 and. 3 were. transiently. + 32-bp DM 9 , were. and by 5' terminally. adding. 32-. from. the former. reporter. gene. was. the level similar. to DM8'cat. activity. (Fig.. 2 ),. the DM 9 cat promoter activity. (Fig.. roles in mediating. activity. up to 82.5%,. 2 ). These findings the constitutive. suggested. promoter. which. that. the. activity. of.
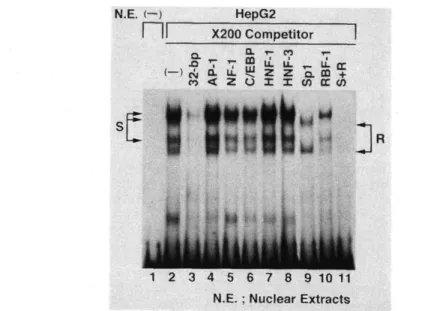
(10) 32b Fig.. 3 Effect of the 32-bp DNA element on the promoter activity. CAT assay was performed and relative CAT activities were determined as described in the legend of Figure 2 . The value of CAT activity represents the mean ± S.E. for six independent experiments.. Identification. of transcription. To identify nuclear nuclear. extracts. probe.. Incubation. factor. binding. complexes nuclear. DNA fragment. factors. factors. prepared. complexes. binding. from. HepG 2 cells,. associated. with. sequence-specific. 4 , lanes. 4 Identification. using. with the nuclear. we performed. the end-labeled. extracts. the five major interaction. since the binding competed (Fig.. to the 32-bp DNA element. that bind to the 32-bp DNA segment,. of this probe. represented. factors,. Fig.. 1. DNA fragment. produced bands. between. with an increasing. the. multiple. ( Fig.. excess. DNA. 2 ). probe. factors. that. bind to the 32-bp DNA element. (-344 to -313). EMSA and competition analyses were performed. Nuclear extracts from HepG 2 cells ( 5 gg) except for lane 1 were incubated with 1 X 10' cpm (0.2ng) of 'P-labeled DNA probe. In the competition assay, 5 -, 10-, 25-, 50- and 200-fold molar excess of the unlabeled element (lanes 3 to 7) were added to the reaction mixture.. —8—. These and. of the unlabeled. 3 to 7 ).. of nuclear. as a. DNA-nuclear. 4 , lane. labeled. molar. EMSA with. DNA.
(11) Inspection. of DNA sequences. bp segment examine. contains. whether. a competition. the putative. their consensus. experiment. complexes. was. containing. the binding. S; Fig.. 6 , lane. of the angiotensinogen binding. partially. inhibited. could prevent. a. 200-fold. sites for constitutively. 9 ). The formation. excess of the oligonucleotides. box binding. RBF- 1 (complex. factor. binding. transcription. (lane factors,. 5 , respectively recognition. 11). No other. sites for typical to inhibit. the formation. activities. could also. be detected. HeLa cells and glioblastoma. the. sequences. various. liver-enriched. 3 failed. second the. the binding. unlabeled. of complexes with. nuclear. T98G cells (Fig.. such as those. these. transcription. 5 ). To formation,. and. fourth. oligonucleotides. was partially. complex. competed of widely. formation. oligonucleotides. factors,. C/EBP,. prevented. sites for novel GC-. R; lane 10). However, both elements consensus. (Fig.. the 32-. GC-box binding factor Sp 1 (complex. containing. AP- 1 and NF- 1 , prevented. ) . Furthermore,. of. of the third and fifth complexes. by the same molar factor. excess. that. complex. of the first,. molar. expressed. Spl. the observed. out. The formation. by. region revealed. sites for AP- 1 , NF- 1 , and. sequences. was carried. upstream. out these expressed. (lanes. 4 and. containing. HNF- 1 , and HNF-. S and. R (lanes. 6. to. 8 ). Such. extracts. prepared. from. cervical. binding. carcinoma. 7 ).. ADA. Abbreviations: K, T/G; M, A/C; N,A/C/G/T; R,A/G Fig.. 5 The recognition DNA fragments for the known transcriptional factors and nucleotide sequences of 32-bp fragment. Small letters indicate the restriction enzyme sites. Putative recognition sequences for known eukaryotic transcription factors, AP- 1 , Sp 1 and NF- 1 were compared with the sequences of the 32-bp element. Different sequences are shown by open squares. K, T/G; M, A/C; N, A/C/G/T; R, A/G.. --9—. the.
(12) Fig.. 6 EMSA and competition analyses using the 32-bp element and the recognition DNA fragments for known transcription factors. Binding reactions using a 32-bp fragment were performed as in the legend of Figure 4 . In the competition assay, 200-fold molar excess of unlabeled 32-bp fragment (lane 3 ), 200-fold molar excess of unlabeled various double-stranded oligonucleotides containing the binding sites for known transcription factors (lanes 4 to 11, AP- 1 , NF- 1 , C/EBP, HNF- 1 , HNF- 3 , Sp 1 , RBF- 1 ), and 200fold molar excess of unlabeled Sp 1 and RBF- 1 (total 400-fold molar, lanel2) were added to the reaction mixture. Arrows point to specific DNA-nuclear factor complexes S and R.. Fig.. 7 EMSA revealed ubiquitous DNA binding activity to the 32-bp element. Nuclear extract from HepG 2 (lane 2 , 3 and 4 , 5 lug), HeLa (lane 5 , 6 and 7 , 5 it g) , T98G (lane 8 , 9 and 10, 5 Atg) were incubated with 1 X 10' cpm (0.2 ng) of VP-labeled DNA probe. Binding reactions were performed as in the legend of Figure 4 . In the competition assay, 200-fold molar excess of unlabeled 32-bp fragment (lane 3 , 6 and 9) and 200-fold molar excess of unlabeled double-stranded oligonucleotides containing the binding sites for C/EBP (lane 4 , 7 and 10) were added to the reaction mixture, Arrows point to specific DNA-nuclear factor complexes S and R. — 10 —.
(13) These. findings. suggested. that. tensinogen. gene might be, at least. we termed. the DNA element. Sp 1 /RBF-. 1 binding. Relationship. between. To examine we performed extracts. was inhibited the formation by. promoter. regulated. between. of binding activity the end-labeled. with increasing of complex. findings. human. suggested. amounts. in this. of complex. of the competitor. condition. complex). ( Fig.. sites prevented. Thus,. seemed. the complex. 4. element,. to be somewhat and. formation. whereas increased. 5 ) . By contrast,. the. of R at 100-fold molar. binding. properties. N.E.; Nuclear Extracts 8 Relationship of Sp1 and RBF-1 to the hASR. Competition experiments of EMSA using the hASR were performed as in the legend of Figure 2 . In competition assay, 200-fold molar excess of unlabeled hASR (lane 3 ), 100- and 400-fold molar excess of unlabeled Sp 1 fragment (lanes 4 and 5 , respectively), 100- and 400-fold molar excess of unlabeled RBF- 1 fragment (lanes 6 and 7 , respectively) and 400-fold molar excess of unlabeled C/EBP fragment. — 11 —. complex). excess (lanes 6 and 7 , respectively).. Sp 1 and RBF- 1 have differential. (lane 8) were added to the reaction mixture. DNA-nuclear factor complexes S and R.. to hASR,. with the nuclear. S (Sp 1 binding. for Sp 1 binding. 8 , lanes. the complex S at 400-fold molar that. as the probe. hASR.. Fig.. angio-. angiotensinogen. between Sp 1 and RBF- 1 proteins. hASR fragment. R (RBF- 1 binding. for RBF- 1 binding. These. human. by Sp 1 and RBF- 1 proteins.. -344 to -313 hASR,. from HepG 2 cells. The formation. and dissociated. of the. Sp 1 and RBF- 1. EMSA using. excess. activity. element.. the Sp 1 competitor. competitor. in part,. located. the relationship. prepared. the constitutive. Arrows. point to specific. to the.
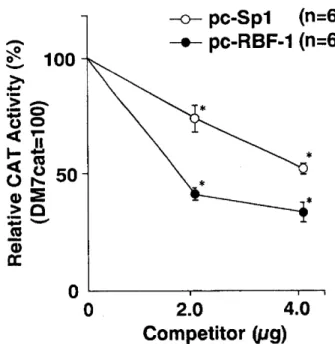
(14) Functional. analysis. of hASR and its binding. We next performed the factors, sequences. in vivo competition. Sp 1 and RBF- 1 , as positive. CAT activities. were analyzed. amounts. RBF- 1 was greater plasmid.. Therefore,. recognition. regulators.. Eight tandem. angiotensinogen. 9. (Fig.. showed. that. than that. of pc-Sp 1 (p<0.01). these functional. in vivo assays. for constitutively contribute. copies of the recognition named. of transfected. pc-Sp 1 and pc-. DNA was normalized. the CAT activity gradually Moreover,. at the concentration suggested. that. decreased. the competitive. activity. nuclear of the. gene.. In vivo competition analysis of the hASR. Competitive plasmids containing eight tandem copies of the Sp 1 fragment (pc-Sp 1) or RBF- 1 fragment (pc-RBF- 1) were used in this experiment. The reporter plasmid of the angiotensinogen promoter-CAT chimeric construct, DM 7 cat (2 jig), was transiently cotransfected with 2 or 4 gg of pc-Sp 1 or pc-RBF- 1 into HepG 2 cells. Total amounts of DNA were adjusted to 6 lig by pUC19. CAT assay was performed as described in the legend of Figure 2 . The CAT activity of DM 7 cat, cotransfected with 4 jig of pUC19, is designated as 100 and each value of CAT activity represents the mean ± S.E. for five independent experiment. * p< 0.01 vs the CAT activity of DM 7 cat cotransfected with 4 jig of pUC19.. — 12 —. with. effect of pc-. hASR is important. of the promoter. by. of 2 jig competitor. active Sp 1 and RBF- 1 , and that both. to the maintenance. roles of both. with DM 7 cat into HepG 2 cells, and the. 9 ). The amount. findings. the functional. into the pUC19 plasmid,. of pc-Sp 1 or pc-RBF- 1 (p<0.01).. sequences. could differentially. Fig.. to confirm. Each plasmid was cotransfected. pUC19 to 6 /Lg. The present increasing. experiments. for Sp 1 and RBF- 1 were inserted. RBF- 1 , respectively.. factors. as the factors human.
(15) Discussion Fukamizu. et al'. and localized. isolated. the promoter. their transgenic. DNA. positions gene. nuclear. that. involved. in. factors,. maintain. transcription. regulates. parental. that. First,. activity. binds. was. DM 7 cat. decreased. significant. reduction. of the promoter. one was first. 32-bp hASR. This observation the -32-bpDM 7 promoter. combination recovered. of hASR. competitors.. effects. activity. the exact of. study,. the. to mediate. derived. activity. This observation. factor. from two lines. the full extent. CAT analyses.. from the DNA/HepG. 2 nuclear. (-516 to -345) (DM7.8cat) no more. of. Second, extracts. mutant. cells,. 51% as compared. with that. DM 8 'cat. removed produces. promoter. from. with. CAT activity. CAT activity. in the promoter. activity. may reflect the differential. analyses. that. (53%) similar in the. ( 44 %). titration. binding affinity. was. of either. in HepG 2 cells showed at the lower. the. only the. reconstituted. in the DM7.8 cat. The sequential. of the in vivo competition. lacking. mutation. + 32-bpDM 9. the lowest. from the parental. 17 % in HepG 2. with the results of further. the CAT activity. DM9cat. than. to approximately. in the deletion. is consistent. on the reduction. common. sequences.. detectable. by means. the well-. binding. results. as much as. that. angiotensinogen. one of the most. and in vivo competition. with hASR internally. with. half. human. (nucleotide. Sp 1 is probably. This conclusion. cis-. we found. of the human. activity.. in the. the hASR was used as the labeled probe was dissociated. to the levels (82.5% ) found. differential. Rb gene.'. in CAT activity. and that. Sp 1 or RBF- 1 binding. promoter. operate. RBF- 1 is a novel GC-box. of the 172-bp sequences. promoter. DM8'cat. genes.". shown. with both Sp 1 and RBF- 1 recognition. of the parental. However,. to the GC-box sequences,. complexes. in EMSA when. While the 5 ' deletion. promoter.. sequences. promoter. by the deletion. of the five shifted. (S and R) detected. DNA sequences. In the present. in the upstream. of the human. the hASR. promoter. from. functional. DNA library. region of the gene. In addition,. upstream. constitutive. of eukaryotic. the expression. the formation. to that. 5 '-flanking. these. its constitutive. factor. DNA elements. of evidence.. the. from the genomic. Sp 1 and RBF- 1 , bind to only an 32-bp DNA region. -344 to -313; hASR) located. regulatory. that. gene. gene have not yet been characterized.. and partially. known. to the 1.3-kb. mice as a constitutively. elements. angiotensinogen two distinct. activity. angiotensinogen. study 6. " demonstrated. liver of transgenic acting. the human. the. concentration. of. of Sp 1 and RBF-. 1 to hASR. Of the five gel-shifted. complexes. generated. nuclear. extracts,. the three. activity. competes. with the Sp 1 consensus. correspond. S complexes. to the 95- and 105-kDa. when the hASR was incubated. appear. to contain. Sp 1 because. GC box sequences.. Sp 1 variants. — 13—. described. this DNA binding. The S complexes previously.'". with HepG 2. most. likely. The appearance.
(16) of such. an. Sp 1 variant. electrophoretic. variants. DNA-dependent result. in EMSA arise. protein. Sp 1 monomer.'. from. kinase.'. from multimerization. has. multiple. of a unique Sp 1 protein. However,. it remains. species. been identified. of the proteins. preferentially. which are. ( positions suggest. with GT boxes.. Sp 2 and Sp 3 appear. Sp 4 is restricted. to the brain.'. -327 to -318).. speculative. by a. can. also. on top of a single DNA-bound Sp 1 itself or closely. characterized. Interestingly,. related have. the hASR contains. extracts. at the moment. including. GT boxes,'. HeLa,. equal. gene. expression,. in our EMSA. However,. and will need to be verified. this. using. to GC. we. correspond in a wide. identification. specific. of. like sequence. of S can also be detected. T98G.. been. whereas. affinity. a GT-box. of Sp 1 family. formation. have. while the distribution. of the three S complexes. the complex. in detail. to GC and. ubiquitously,. In view of the distribution. to Sp 1 , Sp 2 , or Sp 3 because of nuclear. in EMSA. Sp 3 and Sp 4 bind with. to be expressed. that the observed multiple variants. variety. protein. which bind to GC or GT boxes. Sp 2 , Sp 3 , and Sp 4 . Sp 1 binds with high affinity. and GT boxes.. These. by cDNA. to Sp 1 , three. Sp 2 interacts. bands. whether. novel factors. groups.'". of a single. retarded. to be determined. recently. by other. phosphorylation. Furthermore,. bind to the hASR, since several. termed. been reported. differential. proteins. In addition. also. remains. antibodies. for. each. Sp 1 family protein. The formation. of the second. complex. R, which is not. excess. of Sp 1 consensus. GC sequences,. factor. that. to bind to the hASR. Recently,. has the ability. both in monkey. kidney. shown to interact sequence. in the. was efficiently However, containing human. inhibited. thyroid. activity. analysis. responsive. responsive. element. responsive. gene revealed phase. angiotensinogen. that factors. from. element. and regulate. (GRE).'. its upstream 1 and. Sp 1 recognition formation. RBF- 1 binding. region. 2 (H-APF-. responsive. evidence. that,. besides. the. target. for. sequences. for. as the second. element. ( TRE ) and. of the hAG promoter. inspection. contains. the putative. 1 and. H-APF- 2 ).. —14—. sites.. its activity.. sequences. In addition,. of R. of the competitor. DM 7 to DM 9 has shown consensus ( THRE ) , TPA. nuclear. F 9 cells has been. amounts. provides. molar. (RBF- 1) present. existing. containing. gene is characterized. (APRE).15 The other. element. findings. factor. The complex. with increasing. site. Our present. of the region. Rb gene.'. by 400-fold. of an Sp 1 -like. carcinoma. an already. oligonucleotides. did not compete. even. a nuclear. embryonal. of the human. could bind to the promoter. hormone. corticoid. region. the existence. site overlapping. by the competitor. Rb gene, the human. Finally,. acute. 5 '-flanking. the Sp 1 binding. RBF- 1 that. phase. CV 1 cells and in mouse. with a DNA-binding. this binding. suggests. inhibited. have a gluco. of DNA sequences binding. sites. H-APF- 1 and. acute. of the hAG. for. hepatocyte. H-APF- 2 are.
(17) important. for IL- 6 induced C reactive. protein. 288 to -280 which is the APRE consensus of these. regions. transgenic. or both.". hAG mRNA. answering. (described. DNA element. or both. findings raise the question. above) are able to function truly. Further. the experiments required. of the human. that. reported. for, at least. angiotensinogen. involved. here suggested. in part,. gene.. in the maintenance. of many eukaryotic of the. precisely. did not. In the. by LPS or increase. the. whether these consensus. studies. The. that. maintaining. will be needed. present. human. determine. regulatory. promoters. angiotensinogen the molecular. mechanism. gene. relationship. of eukaryotic. hASR is an. characterization. of the angiotensinogen. and enhancers. the. the constitutive. toward. important. promoter. of Sp 1 and. promoter. not only the Sp 1 family but also RBF- 1 play a critical. hARS. activity. the above questions.. In conclusion,. possibly. the constitutive. mRNA was not induced. IL- 6 or glucocorticoid. level in HepG 2 cells." These. between -. by Sp 1 -like and RBF- 1 -like proteins.. the hAG gene, the transgene. Furthermore,. H-APF- 1 maps. site. In our experiments,. to be regulated. mice containing. glucocorticoid. sequences. seemed. gene expression.'. activity RBF- 1. activity. also implies. role in regulating. the activity. along with GC or GT boxes. will be an. excellent. model. between. Sp 1 family. In this respect, DNA. element. to. and RBF- 1 as to the. gene expression.. Acknowledgements We would Internal. like to express. Medicine,. Medicine,. Kinki. for their continuous (University. thanks. to Professor. University ), Dr. Junkichi. University ), Dr. Akiyoshi. Kazuo Murakami. Sagara. Kinki. our. (University. of Tsukuba),. encouragement. of Tsukuba). Ryo Katori. Hama. Fukamizu. ( First. ( University. and Dr. Masatsugu. Department. of. Department. of Internal. of Tsukuba). , Professor. Horiuchi. We also thank Mr. Kazuyuki. for their technical. ( First. (Stanford. Yanai and Mr.. advise and helpful. discussion.. University) Masashi.
(18) References 1 . Campbell. DJ. Circulating. 2 . Jeunemaitre Hopkins. X, Soubrier PN, Williams. hypertension: 3.. Ward. and tissue. F, Kotelevtsev RR,. Lalouel. Ogasawara. A, Jeunemaitre. M, Suzumori. J-M,. (1987) J Clin Invest 79: 1-6.. RP, Williams. Corvol. CS, Charru. P ( 1992 ) Molecular. A, Hunt SC,. basis. of human. Cell 71: 169-180.. X, Helin. K, Tomoda. Lalouel J-M (1993) A molecular. systems. YV, Lifton. role of angiotensinogen.. K, Hata. Nature. angiotensin. C, Nelson. S, Berrebi. variant. L, Namikawa. S, Sasaki. of angiotensinogen. C, Farrington. M, Corvol. associated. PF,. P, Lifton. RP,. with preeclampsia.. Genet 4: 59-61.. 4 . Caulfield. M, Lavender. Linkage. P, Farrall. M, Munroe. of the angiotensinogen. P, Lawson. gene to essential. M, Turner. hypertension.. P, Clark AJL (1994). N Engl J Med 330: 1629-. 1633. 5 . Fukamizu. A, Sugimura. Kajiwara. N, Yagami. demonstrates human. K, Takimoto K,. sustained. Murakami. increase. S, Fukamizu. in blood. A, Hasegawa. K ( 1991 ) Expression transfected. C, Ganten. Endocrinology. Chimeric. pressure. S, Hatae. renin-angiotensin. of transgenic. T,. system. mice carrying. both. T, Yokoyama. M, Nomura. angiotensinogen. Res Commun. W, Kaking. T, Katsuki. gene. in. M, Murakami. transgenic. mice. and. 180: 1103-1109.. M, Ryffel GU, synthesis. Weimar-ehl. and secretion. T, Hackenthal. E. by steroid hormones.. 130: 3660-3668. AJ, Oparil. gene expression 10. Itoh N, Matsuda. S (1992) Androgen. in hypertensive. 9 . Hong- Brown LQ, Deschepper. LETTERS. ( 1993). of hepatic angiotensinogen. RNA expression. hepatoma. F, Seo M-S, Takahasi. genes. J Biol Chem 268: 11617-11621.. human. D, Hellmann. 8 . Chen Y-F, Naftilan messenger. of the. cells. Biochem Biophys. (1992) Regulation. rats.. CF (1992) Effects. in rat liver, brain, T, Ohtani. and cultured. R, Okamoto. cells is stimulated. dependent. angiotensinogen. Hypertension. 19: 456-463.. of thyroid. hormones. on angiotensinogen. cells. Endocrinology. 130: 1231-1237. H (1989) Angiotensinogen. by B cell stimulatory. factor. and renin. production. by rat. 2/. interleukin-. 6 . FEBS. in angiotensinogen. messenger. RNA in. 244: 6 -10.. 11. Ssy JA, Lynch KR, Peach differentiating 12. Nielsen. K. renin and human angiotensinogen. 6 . Takahashi. 7. Klett. E, Sugiyama. MJ. (1990) Changes. 3 T 3 -F442A adipocytes.. AH, Knudsen. Hypertension. F (1987) Angiotensinogen. 1990; 15: 867-871. is Acute-Phase. Protein. in Man.. Scand. J. Clin Invest 47: 175-178. 13. Ron D, Brasier on interleukin-. AR, Wright KA, Habener JF (1990) The permissive 1 stimulation. of angiotensinogen. — 16 —. gene transcription. role of glucocorticoids is mediated. by an.
(19) interaction 14. Tamura. between K, Tanimoto. DNA Element Adipogenic. Structure. Are Required. for Efficient. Promoter. and Core. Activation. During. Biol Chem 268: 15024-15032.. and expression. of the human angiotensinogen. A, Tanimoto. A (1993) Proximal. Angiotensinogen. S, Seo MS, Tada M, Tanimoto. T, Murakami. K, Uehara. of human. K, Uehara. S, Murakami. K (1990). gene. J Biol Chem 265: 7576-7582.. S, Seo MS, Handa. K (1991) Regulation. M, Sagara. M, Takahashi. renin and angiotensinogen. S, Imai. genes.. Biochem. which. express. Acta 50: 659-663.. Gorman. CM, Moffat. chloramphenicol 18.. K, Fukamizu. A, Takahashi. 16. Fukamizu. Biomed. Mol Cell Biol 10: 4389-4395.. K, Ishii M, Murakami. Differentiation.J. 15. Fukamizu. 17.. inducible enhancers.. Dignam. JD,. polymerase. LF,. Howard. acetyltransferase Lebovitz. RM,. II in a soluble. B ( 1982). Recombinant. in mammalian. Roeder. RG. extract. from. genomes. cells. Mol Cell Biol 2 : 1044-1051. ( 1983 ) Accurate isolated. transcription. mammalian. nuclei.. initiation. Nucleic. by. Acids Res. 11: 1475-1489. 19. Landschulz. WH, Johnson. recombinant 20. Courtois. PF, Adashi. EY, Graves. copy of the gene encoding. G, Morgan. liver-specific. JG, Campbell. nuclear. factor. BJ, McKnight. C/EBP.. LA, Fourel. SL (1988) Isolation. of a. Genes Dev 2: 786-800. G, Crabtree. with the fibrinogen. GR (1987). and al-antitrypsin. Interaction. promoter.. of a. Science 238:. 688-692. 21. Costa. RH, Grayson. function. DR, Darnell. in the regulation. JEJr. (1989) Multiple hepatocyte-enriched. of transthyretin. and al-antitrypsin. nuclear. genes.. factors. Mol Cell Biol 9:. 1415-1425. 22. Sakai. T, Ohtani. mutations. S, Fukamizu. Species-specific phase. 24. Kadonaga. proteins. 25.. Briggs. JT,. Tjian. ( 1991 ) Oncogenic. retinoblastoma. F, Kajiwara. of angiotensinogen. TP. gene. Nature. N, Yagami. mRNA. germ-line 353: 83-86.. K, Murakami. in transgenic. mouse. K (1992) liver during. J Vet Med Sci 54: 367-369. R (1986). Affinity. purification. of sequence-specific. DNA binding. Proc Natl Acad Sci USA 83: 5889-5893. MR.,. Kadonaga. JT,. Bell SP, Tjian. of the promoter-specific. SP, MacDonald. phosphorylation 27. Dawson. A, Sugiyama. induction. reaction.. characterization 26. Jackson. PD, Dryja. in Sp 1 and ATF sites in the human. 23. Takahahsi. acute. N, McGee TL, Robbins. JJ,. transcription. Lees-Miller. of Spl by DNA-dependent. PA, Hofmann. (1988) Sterol-dependent. S. Tjian protein. SL, van der Wethuysen repression. R ( 1986). factor, R (1990). kinase.. DR, Sudhof. of low density. — 17 —. Purification. lipoprotein. and. biochemical. Sp1. Science 234: 47-52. GC box. binding. induced. Cell 63: 155-165. TC, Brown MS, Goldstein receptor. promoter. JL. mediated.
(20) by 16-base. pair sequence. adjacent. to binding. site for transcription. factor. Spl.. J Biol. Chem 263: 3372-3279. 28. Gustafson. TA, Kedes. functional. regions. 29. Sartorelli. of the human. V, Webster. actin gene requires 30.. Pascal. KA, Kedes. cardiac. MyoD1, CArG-box. 31. Kingsley. and mediate C, Winoto. family regulating. activation. promoter.. with. of the cardiac. a-. Genes Dev 4: 1811-1822.. of Sp1 govern. formation. of. proteins:. a novel Sp 1 multigene. Mol Cell Biol 12: 4251-4261.. G (1992) Cloning a family. interact. Genes Dev 5: 1646-1656.. gene expression.. two novel GT box binding proteins:. that. expression. and Spl.. of GT box-binding. M, Suske. protein. Mol Cell Biol 9: 3269-3283.. domains. synergism.. A (1992) Cloning. G, Muller S, Beato. a-actin. binding factor,. transcriptional. T-Cell receptor. of multiple. L (1990) Muscle-specific. E, Tjian R ( 1991) Different. multimers. 32. Hagen. LH ( 1989) Identification. by recognition. of Sp 1 related. site screening. genes.. of. Nucleic Acids Res. 20: 5519-5525. 33. Imataka. H, Sogawa. K, Yasumoto. K, Kikuchi Y, Sasano. Fujii-Kuriyama. Y (1992) Two regulatory. element. a GC box sequence. (BTE),. proteins. K, Kobayashi. that. in the promoter. A, Hayami. bind to the basic region. of the rat. M,. transcription. P-4501A1 gene.. EMBO J 11: 3663-3671. 34. Sogawa. K, Imataka. cloning. and. H, Yamasaki. transcriptional. Y, Kusume H, Abe H, Fujii-Kuriyama. properties. of a novel. GC box-binding. Y (1993). cDNA. protein,. BTEB 2 .. of cDNA. encoding. domain.. Cell 51:. Nucleic Acids Res 21: 1527-1532. 35.. Kadonaga transcription. JT, Carner factor. KR, Masiatz. Spl and functional. FR Tjian analysis. R (1987). Isolation. of the DNA binding. 1079-1090. 36. Majello. factors. B, Arcone R, Toniatti that. interact. C, Ciliberto G (1990) Constitutive. with the human. C-reactive. protein. and IL-6-induced. promoter.. nuclear. EMBO J 9: 457-465..
(21)
図

関連したドキュメント
Then he found that the trapezoidal formula is optimal in each of both function spaces and that the error of the trapezoidal formula approaches zero faster in the function space
The edges terminating in a correspond to the generators, i.e., the south-west cor- ners of the respective Ferrers diagram, whereas the edges originating in a correspond to the
H ernández , Positive and free boundary solutions to singular nonlinear elliptic problems with absorption; An overview and open problems, in: Proceedings of the Variational
Keywords: Convex order ; Fréchet distribution ; Median ; Mittag-Leffler distribution ; Mittag- Leffler function ; Stable distribution ; Stochastic order.. AMS MSC 2010: Primary 60E05
In Section 3, we show that the clique- width is unbounded in any superfactorial class of graphs, and in Section 4, we prove that the clique-width is bounded in any hereditary
Inside this class, we identify a new subclass of Liouvillian integrable systems, under suitable conditions such Liouvillian integrable systems can have at most one limit cycle, and
Then it follows immediately from a suitable version of “Hensel’s Lemma” [cf., e.g., the argument of [4], Lemma 2.1] that S may be obtained, as the notation suggests, as the m A
[Mag3] , Painlev´ e-type differential equations for the recurrence coefficients of semi- classical orthogonal polynomials, J. Zaslavsky , Asymptotic expansions of ratios of
