September
,1987 Acta
Phytotax
. Geobot .275
ヒ ル
ギ 科 の 範 疇 と 類 縁
戸
部
博*Hiroshi
ToBE : The Rhizophoraceae : circumscription and relationg .hips
ヒ ル ギ科の 属 (後述)は, それ ぞ れの分 布域 はいろいろに偏 る が, 全 体 と しては熱 帯 全 域に 分 布し て い る。 日木で はヒ ル ギ科 と言え ば琉球 列島 (西表 島)に分布するヤ エ ヤマ ヒル ギの 他,
オ ヒ ル ギや , メ ヒル ギ な ど, マ ング ロ ーブ植 物の代 表 的な例と して 知 られて い る。ヒル ギ科を
マ ン グロ ーブ植 物で代 表させ て し まう傾 向は日本ば かりでな く世 界で も
同様で, そ のた め ヒ ル ギ科の これまで の研 究の 多 くがマ ン グロ ーブ植 物につ い て 行 わ れて きて お り
, 科の大 部 分を占
め る内 陸 性の植物につ い ては, ほ と ん ど研究 さ れて いな か っ た。し か しマ ングロ ーブ
植 物は ,
ヒル ギ科の 中で は進 化の末 端にある植 物 群である た め, これを研 究 して もヒル ギ科を 正 しく理 解 する には 程 遠い もの で あっ た。 こ の た め, 我 々 は今 まで ヒル ギ 科につ い て, その 特 徴 も定 義 も あい まい に しか 理 解 して いな か っ た と言え る。 同 様に, ど ん な属や 種 が ヒ ル ギ科 と してま と め られ, それ らが 他の どん な植 物群と類縁 を もつ の か, 明 確には何 も 分か っ て いな か っ た. こ の よ うな背 景か ら, 数 年 前か ら, 私 自 身を含む 8人の研 究 者の協 同に よっ て い ろいろ な角 度か らヒル ギ科が研 究され, ヒル ギ科の実 態 を 明 らか に し てきた。 それ らの成 果は, 1986年
8
月ア メ リカ植 物 学 会におい て ヒ ル ギ科の シ ン ポ ジ ウム で発 表さ れ
( Amer
.J
.Bot
.,73 (
5)
:746
−748
[Abst .]), 近 くAnnals of the Missouri BotanicalGarden
にま とめて印 刷される予 定になっ て い る。そ こ で , こ こ で発 表され る予定の私 自身の を含め た幾つ かの論 文 と, こ れと は別に発 表 した幾つ かの論 文や 現在 投 稿中の論文( ToBE
& RAvEN , 1987a,b
, c)
とを 併せ,ヒ ル ギ科と は どの ような植物 群か, 主 と して比較 形 態 学 的な観 点 か ら紹 介 したい。 最 近は N本
で も, ヒル ギ科に関 心 を持つ 研究 者 も少な くな い が, ヒル ギ科の系 統 群と し て の “ま とま り”
と, その 類 縁につ い て の , 現 在 まで の情 報と考え方 を大ま かに で も理 解して も らえ たら, 今 後 一層深 く研 究し て い く
際の手 助 けに な る かも知 れない 。
ヒ ル ギ 科の構成と類 縁に関 す る 歴史 的 概観
始めに, ヒ ル ギ科が どの ような属や種 か ら構 成 され, それ ら がヒ ル ギ科の 中でどのよ うに 亜 科や連にまと め られて きた か見て み よ う。
そ もそもヒル ギ科
( Rhizophoraceae
)は, LINNEAus(
1753)
によ っ て作 ら れ た Rhizophora 一属 か ら 出 発 し, BRowN(
1814 )が Rhizophora , Bruguicra ,Ccr
三〇psか ら成る科 と して建てた もの で あ る。しか し, LINNEAus の
Rhizophora
8種の う ち, 現在 も正 当な種と して残 っ ているの は唯一種
・
R
・mangle ,だ けで他はBruguicra
や Kandelia な どの ヒル ギ科の他のマ ン* 京 都 大学教 養部生物 学教室
Vo1
. XXXV 皿Acta
Phytotax
.Geobot
.276
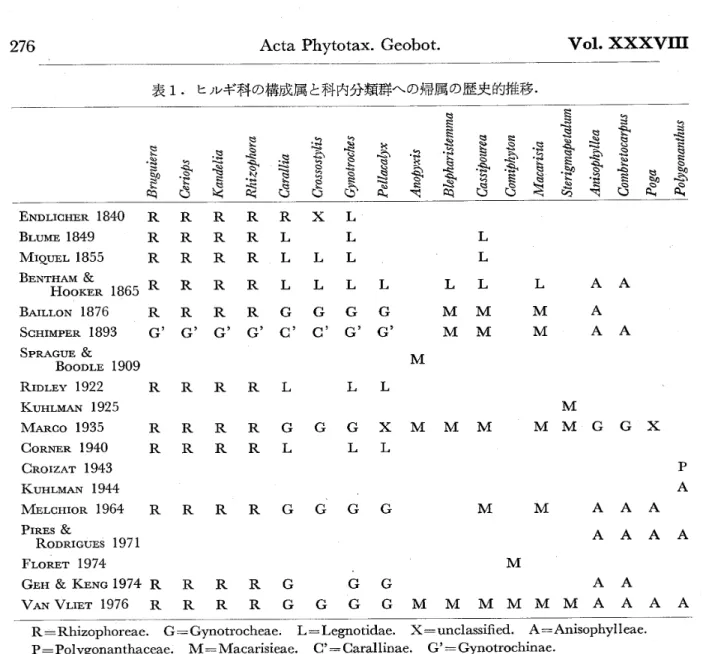
表 1.ヒ ル ギ科の構 成 属と科 内 分 類 群へ の帰属 の歴 史 的 推 移
§ ミ
簔 唱 霜O 吻
高
ミ
ミ鶚 冖 S§
竃O 竜
お§
δ
曽
ミN
爵魯
叫
黛 団
四
§ ミ
鳶
魯
ミ 欝
蠢 お
q騨
轟L
竜噛
羮
皿一
黌 ミ
爵魯 ミ
δ.
§
§ 魯
題
δ
電
§ ミ ミ 麓 喫竜
§Q
鬟
§
超融 ゐ
§
ミ 魅 へ 竈 ミ
ミ 巷匿
台
訟
丶
急 旨逡
06
ミ
越
ミ
δ
ミq
哩
SN
ミ 叱 滋吋
愚 咤§
碁
黛
δ 讐§
ぎ
箱
PA
A
A ENDLIcHERI840 R R R R R X I.
BLUME l849 R R R R L L L
MIQuELI855 RRRRLLL L
BE瞬T
翻 盞。 1865RRRR ・ …
LL
舳 LエoNl876 RRRRGGGG MM
SGHIMPER189S G ’ G ’ G ’ G, G , α G’ G ,
MM SPRAGUE &
M
BoOD エE l909
RIDLEY1922 RRRRL LL
KUHLMAN l925
MARao1935 RRRRGGGXMMM
CORNERI940 RRRRL LL
CROIZAT l943 KUIILMAN l944
MELcH 匸oR }964 R R R R G G G G PIRES&
RODRIGUES 1971 F正ORET l974
GEH & KENGl974RRRRG GG
L AA M A M AA
M
.MMGGX
M M AAA
AAA
M
AA
VANVHETI976RRRRGGGGMMMMMMAAA
R=Rhizophoreae . G =Gynotrocheae. L− Legnotidae. X =unclassi 丘ed , A=Anisophy11cae. P=Polygonanthaceae . M 篇Macarisieae,α 一〔】arallinae . G ’=Gynotrochinae.
グ ロ _ブの属 や
Sonneratiaceae
のSonneratia
やMyrsinaceae
の Aegiceros な ど他の 科の マン グロ ーブの属 に移されて い る。 その 後, ENDLIGHER
( 1840
),BLuME
(1849)
, BENTHAM &HooKER
( 1865 )
,BAILLoN ( 1876
)そ し てSGHIMPER
(1893 ), RIDLEY( 1922 )
等に よ っ て既 成の属や新 属が ヒ ル ギ科に加え られ, M ELaHIOR
(
1964 )に 至 っ て, 不 完全 な が ら今日我々 がヒル ギ 科につ い て抱 く全 様の よ う なもの が見ら れ るこ とに な る。
M
肌 CHIOR (1964)
の 分 類は 基図 1.花 弁の形 態.“Anisophyllcac” の花 弁 はヒ ルギ科の花弁と良 く似て い る.時には, 花 弁の 細 裂 片 先
端に腺 (9)を持つ , 左 :Anisophyllca laurina R・BR・ex SABINE (1)・Thomas 4522・MO ); 中 央 ;
Blepharistemma membranif ()lia(MI皀.)DING 且 ou (Moniat s.n. in 1984, MO ); 右 :Ceriops tage1
(PERR.)G . B. RoB .(Hemaez (]A 29249, CAHUP ).スヶ一ルは 1mm .
図2 .(】assipourea gummifiua
TuL
. var . vcrticiHata (N. E. BR.)j
. LEwis (Tit. Mfiller S558, SRGH )の 子房 壁の縦 断 切片.放射 方 向に伸 びた細胞か ら成る一層の樹 脂 細 胞 層 (1)が観察さ れ る.ov は 胚珠,ス
ケール は 10
μm .
図3 ,Sterigmapctalu皿 heteroxoxum S[[EYERMARu [ & LIEsNER の シュ ートの横 断切片.
托葉 (st)上に
colleter (co)を持つ .スケール は 0・5 mm ・
図4 .Carallia brachiata(LouR .)MERR .の胚 珠の縦断切 片.ヒ ル ギ 科 (Rhizophoreae は除 く)に特徴 的
な endothelium (=・ integumentary tapetum )(end )が 観 察される.ス ヶ一ル は 20岬 .
図5.Blepharistemma membranif ()lia(Sasidaran s.n. in l986, MO )の果 実 と種 子 (果 皮の一部は取 り除い てあ る ).種 子に はaril (ar)が付い て い る.ス ケール は lmm .
September
,1987 Acta
Phytotax
.Geobot
. 277278 Acta Phytotax . Geobot .
Vo1
. XXXV 皿1本 的には BAILLON
(
1876)の それ と あ ま り変わ っ て いな い が, その 後の ヒル ギ科の 分類 学や 比 較 形態学の 研究の 骨 格と もな り, 同時 に そ れ らの 研 究 の 文持 も 受けて きて い る (例 ,VAN VLIET
,1976
一材 解 剖 )。ヒル ギ科の細 分 類とそ の構 成 属につ い て は, VAN VLIET(
1976>
が 比較的 分か りやす くレ ビ ュ ーして い る,表 1 に は,VAN VLIET に な らい、それ を 少 し改 変して,
これ まで の ヒル ギ科の属 の歴 史 的 推 移を示 し た。 それに よれ ば,
VAN
VLIET
(1976
)の時 点で , ヒル ギ科は 18属か ら成 り, 4 つ の 連 に ま とめ ら れて い る。即ち, (1)Rhizophoreae
(Bruguicra,
Ceriops
,Kandelia
,Rhizephora
), (2
)Gynotrocheae
(Garallia
,Crossostylis
,Gynotrochcs , Pcllacalyx
)
,(
3) Macarisieae (Anopyxis
,Blepharistcmma
,Cassipourea
,Macarisia
,Sterigmapetalum
), (4)Anisophylleae (Anisophyllea, Gombretocarpus , Poga , Polygonanthus)
, であ る。こ の う ち,Rhizophoreae
に まと め られる 4 属 だけがマ ン グロ ーブ 植 物で , ほかの /4属は 内陵 性 植物であ る。しかし一方,とりわけ 最近 にな っ て , 上 記の 属 を含む ヒ ル ギ科は, 異質な群の 集ま りで ある らしい と指 摘 されて きて いた (CRoNQuls ・r,1981, 1983; ToBE & RAvEN , 1983 )。 特に,
Anisophylleac
につ いて は, 他の群 とのギ ャ ッ プ が 指摘さ れ, ヒ ル ギ科の中で 亜科 Anisoph − ylleoideae と して他 の 全 属 か ら 成る亜科Rhizophylleoideae
か ら 区 別 さ れ る か (THoRNE
,1983
), ヒ ル ギ 科 から除か れて独 立 科 Anisophylleaceae と して 扱わ れ る傾 向にあ る (AIRYS
且Aw , 1966;CRoNQulsT
,1981
, 1983;DAHLGREN , 1983 )。次 に, ヒル ギ科の 類 縁につ い て見 る と, ヒ ル ギ科は, 伝統 的に, 曖昧に定 義さ れ た フ トモ モ 目の 中に い れ られ て,
Combretaceae
あるい は Lythraccae と近 縁 と考え られて き た (例 ScHIMpER ,1893
;MELcHloR
, 1964;TAKHATAJAN , 1980)
。 し か し,フ トモ モ 目の研究 が進む と,知 ら れ る限 りの ヒル ギ科の特徴は フ トモ モ 目の そ れと一致 し ない こ と が明ら かにな る。
こ の事 実が積 極 的に受 け止 め ら れ て, 最近で は, ヒ ル ギ科は ミ ズ キ 目 (THoRNE , 1968, 1983; VAN VLIET , 1976 一材 解 剖),あるい は単型 の ヒル ギ 目に移 す提 案が な さ れて い る
( CRoNQulsT
,1981
;DAHLGREN , 1983)。 し か し, 前 述の よ うに , マ ン グロ ーブ植 物を除け ば, ヒル ギ科は,
この 時 点ま で は ま だ ほ とん ど研 究されて いな か っ たため, 他の どの 科 或は群と近 縁 か は全 く議 論されて いない。
ヒ ルギ科は 異質な群の集 合か
上 述した 4連18属につ い て, 全 属 に基づ いた こ れまでの比 較 研 究 (主と して ,比 較 解 剖に よ る)か らの結 論は,“
Anisophylleae
”の 4 属 は 明 ら かに他の全 属 と多 くの重 要な点で異な り,ヒ ル ギ科に馴 染ま ない , とい うこ と で あ る。 例えば,
“Anisophylleae ” では, ヒル ギ科の 他の 属と異な り,師 部 細 胞の 色素休 含有 物が
S
−type(
ヒル ギ科で はユ ニ ークなPVc
−typc )(BEHNKE ,1984
),葉に アル ミニ ュ ゥム を蓄 積 する( CHENERY
, 1948;KuKAcHKA
&MILLER
,1980
), 無 托 葉の葉 と 互 生葉序を持つ , 合生子房 の 花 柱が 離 れて い る (GEH & KENG , 1974 ), 胚珠が endothelium を発達 させ ない ,内珠 皮が 2細 胞層 (ヒ ル ギ科で は, 多層), 成熟 種 子 は 胚 乳 を 欠 く, 種 皮 がfibrous
exotegmen を欠い て い る (以 上, ToBE & RAvEN , 1987a, b, d)。 仮 に, こ れらの 相 違 点を無 視 し た場 合, ‘‘Anisophylleae
” を含む ヒ ル ギ科は, 著し く捉 えど こ ろ の な い 集 合 体に な るこ とは疑いな い。上 にあげた 重要な相違点に も関 わ らず,
[CAnisophylleae ”
は花の形 態の点で は, ヒル ギ科の 他の属 と実に興 味 深い一
致を示 す。 即 ち, ‘‘
Anisophylleac
”( Polygonanthus
を 除 く)の 花 弁がSeptember
, 1987Acta
Phytotax . Geobot .279
細か く裂片 化 し (ToBE & RAvEN , 1987c
)
, 同 様 の こ と が ヒル ギ 科の他の 属 (Rhizophora ,(〕rossostylis を除 く
)
で も見 ら れ る (図 1)。更に,Anisophyllea
の 種とCeriops
の種は花 弁 裂 片の先 端 に腺 (gland )を持つ 点で も一致 する. ‘cAnisophylleae ”が ヒ 丿レギ科と共 に扱 わ れ
て き たの は, 花 弁の 形態の 類 似を含む, こ の様な花の基本構造の 類似の ため か も知 れ ない 。 花 弁の裂 片 化は,外に幾つ かの科 (例え ば,Elaeocarpaceae ,
Malvaceae
,Euphorbiaceae
)にも観 察されてい る。更 に,材 解 剖の研 究 も また,
“
Anisophylleae
,’と ヒル ギ科のGynotrocheae
(
特に,Carallia )
との 類 似を指摘 して い る(
VAN VLIET , 1976 )。 し か し, Gynotrocheae は,そ れ自身,ヒル ギ科の中で も特殊 化 した群で あ り, ‘‘
Anisophylleae
” とGynotrochcac
との材 構 造の類似 は お そらく convergence によ る ものと 思わ れる。
‘‘
Anisophylleae
” は, cmbryology の 形 質 の 比 較 に よ れ ば, フ トモ モ 目と殆 ど一致 す る(
ToBE & RAvEN , 1987a)
。即ち, (1)タペ ート細胞 がglandular
, (2)胚珠は crassinucellate ,(3)内珠 皮が 2 細胞 層, (4)珠孔 が 内 ・外の 二 枚の珠 皮で 形 成
, (5) 反足細 胞は短 命, (6) 胚 乳 形 成は nuclear 型 , (7 ) 種 子は無胚乳, (8) 花.粉は送 粉 時 2細 胞 期, の 8つ の特 徴の組 合 せ は, フ トモ モ 目の embryology の 定 義と して与え ら れ た が
( ToBE
&RAvEN
,1983
, 1984 ),‘ ‘
Anisophyllcae
” は こ の条件 を満たすの で ある.し か し, ‘‘Anisophylleae
” は, フ トモ モ 目の定 義とも言 える vestured pits!
intraxylary
phloem を共 有 しない。 更に, 厳 密に は, 花の形 態 ・胚の 形態 ・種皮の 形 態 な どで も, 両 者 は 異な る。DAHLGREN ( 1987
)は ‘Anisophylleac ”を
CRoN2ulsT (
1981, 1983)
と同様,独立科Anisophylleaceae
と し てバ ラ 目の 中に移 すの が 妥当 と 判断して い る が, バ ラ目の 中に具 体 的に近 縁 群は見 あた らな い。Anisophylleaceae
の類 縁につ い て は, 現 在ま だ総 意が得ら れて い な い 。
ヒ ル ギ 科 の 定 義
最 近 出 版された分 類の テ キス ト
(
例,CRONQT 〕ls’r, 1991)
で見 られ る シ ノ プシ ス か ら も 分かる ように, ヒル ギ科 (“Anisophylleac” は除 く)の定 義は, 何 十という形質の特…徴の組 合せ に
よっ て与え られて い る。 し か し, その多 様 性の 故に , ヒル ギ科だ けを明確に特 徴づ ける特 徴は 何なの か,ヒル ギ科の 記載を読んで も殆ど 分 か ら ない 。ヒル ギ科が頁に単型 群で ある こ との明
確な根 拠さ え も定かで な い。 花を見 れば, 雄 芯の位 置は子 房上位 ・下 位 ・周 位な ど が あ り
, 果
実を見れ ば, 朔 果 ・液 果, その外マ ン グn 一ブ 植 物に特 有の胎 生 発 芽をする閉 鎖 果
, な ど さ ま ざまである。 解 剖 学 的に も,材 構造
( VAN
VLIET
,1976 )
も embryology( ToBE
& RAvEN ,1987a
,b
,d
;JuNcosA
, 1987 )も, 他の科に比べ て , ヒル ギ科14属が 全体と し て か な りの多 様 性を示す こ と が 分 か っ てい る。し か し,これ ら14属は,子 房壁等に樹脂細胞層 (図 2)を も ち, 托 葉に colleter (図 3)を 持つ , とい う点で際だ っ た一致を見せ る
。 子 房 壁の樹 脂 細 胞 層の存 在は,
JuNcosA
& ToMLI −NsoN (1987 )よ っ て, マ ン グu 一ブ植 物
(
Rhizophoreae )に つ い て初めて 明 らか に さ れ た が, FLoRET (1974)は既に内 陸性のComiphyton
で気 がつ い て い た。 その後・ 私 自 身の観 察によっ て, Pellacalyx (ヒ ル ギ科の 中で もっ と も進 化 し た内 陸 性 植 物 )を除 く全 属に同 様の 樹 脂 細 胞 層が確認 された
(
ToBE & RAvEN , 1987d)
。 colleter は,GILL
&ToMLINSON
(1969)に ょっ て
Rhizophora
mangle に初め て 記録され, そ の 後 LERsTEN & (】uRTIs(
1974)
によっ て組織 学 的 特 徴が明 ら かにさ れ た。 そ れ らによ れ ば, colleter は放 射 方 向に長 く伸 長 した表 皮 細 胞 を持ち, そ こ か ら糖を含む粘 液 性 物 質を分 泌 す るた め, そ れによ っ て鳥 を誘引 し, 虫 害 か ら身
280 Acta
Phytotax
.Geobot
,Vol
.XXXV
皿を 守 っ て い る ら しい,と想 像さ れて い る が 確 か な こ とは分 か っ て い ない。 colleter は,その
後, 全マ ン グロ ーブ植 物
( Rhizophoreae
)に観 察さ れ(
ToMLiNsoN et al.,1979
),さ ら に私 自 身によ っ て利用で きる全 内陸性 植 物 に も 確認 されてい る。 子麓壁 内の樹 脂細胞層 と托 葉 上のcolleter は個々 には希に外の群 にも』
出現する が, こ れ らを同時に共 有 する植 物 群は ヒ ル ギ科14 属以外には 知 ら れて いな い。こ の こ と か ら, ヒル ギ科が単型群であ るこ と は疑いないと 思 わ れ
る。
ヒ ル ギ 科 の 類 縁
ヒル ギ科の類 縁 群を探 すに は, 多 様な ヒ ル ギ 科の 特 徴の中の , 何を基に外の植 物 群と比 較 し た ら良い か判断 し な くて はな らな い 。 即ち, ヒ ル ギ科 の 原 始 的特 徴 (或は, 祖 先 的特徴 )は 何かを明 ら かに し, そ れ に 基 づ い て似た特 徴を持つ 近 縁な植 物 群を探さ な けれ ばな ら ない。
DA 肌 GREN
(
1987)
の 方 法を紹 介 し たい。 そ れによ る と, 近 縁 群を探 す 際の最 も成 功 しそ う な形質 と して,次の 条件 を,同時に満たす 形 質 (状態)を探 すこ と を勧めて い る。 即 ち, (1) 全
ヒ ル ギ科, 又 は, 多 くの ヒ ル ギ科に見 られる形 質 (実 際に は, 形 質 状 態 似 下 同様}), 或は少
な く とも, 全 内 陸性属, 又 は, 多 くの 内 陸 性 属に見 ら れ る形 質。な ぜ な ら, そ うし た形 質はお そ らくヒル ギ科の原 始 的形 質
(
Plesiomorphy)
だ か らで あ る。(2) 高 次 分 類 群
(
superorder )を特 徴づ ける進 化 した形 質 (apomorphy )(被 子 植 物の 中で は原 始 的で は ない 形質)。 (3) 被 子 植 物 として は特 殊で , かつ , 限 られた植物群 (つ ま り, ヒ ル ギ科と その 類縁群) にの み 現れる 形 質。
こ の方 法によっ て得 られた, ヒ ル ギ科及 び その近縁群に共通 の進 化 し た特徴
(
synapomorphy )は, (ユ)endothclium がある (図 4), (2)aril が ある (図 5), (3) 種 皮は exotegmic であ
る, (4)胚が 葉 緑 体 を 持つ , (5) 師部細 胞の色 素 体 含 有 物が
PVc
−type , (5
) 或 種の アル カロ イ ド( pyrrolidine
, tropane ,pyrrolizidine
)を持つ , (6) 幾つ かの embryology の形 質 状 態の 組 合せ を持つ , であ り,そ して, こ れ らの特 徴を ヒ ル ギ科 と共 有 する群と して , 最 も比較 し うる科は,
Elaeocarpaceae
(Malvales
)とCelastraccae ( Celastrales )
で あっ た.E正acocarpaceae は 約10属400種 か ら成る熱 帯 植 物 群で ある。 こ の科は, しばしばTiliaceae
の 中に入 れら れて (HucHINsoN
,
1959
), これ らの科を扱 う者を 混 乱 さ せて き た が, 一般には独立科 と して Ma ]vales の 中で 最 も原 始 的な群と考え られ て き てい る。 DAHLGREN
( 1987
)は,研究の 結 論の 一つ と して
,
Elaeocarpaceac
をMa
ユvales か ら除 外 す る こ と を 主張 して いる。そ の根 拠と して , Elaeocarpaceae は Malvales の特 徴である粘 液 細 胞 ・星 状 毛 ・萼 基 部の蜜 腺 毛 ・或 種の脂 肪 酸を欠 き
, 更に, Malvales に は 見 ら れ ない覆 瓦 状 萼 片 ・雄 蘂基部に蜜 腺 ・
endothelium を示 す, とい う点をあげて い る。 こ の 結論には, 私 も賛成であ る。 Malvalcs の
科の種皮は,放 射方向に伸 長 し た細 胞か ら成 るexotegmen を持つ (お そら く, synapomorphy ) 点で 明確に特徴ず けら れ るが, Elaeocarpaceae の種 皮 は こ の 構 造 を 持た な い か らで あ る
(ToBE & RAvEN
, 正
987d
)。 一方,Elacocarpaceae
とヒル ギ科を 比較 してみ る と,花 部や 栄 養 部の形態 (特に ,細 く裂 片 化 し た花 弁 )・材 解 剖 ・節 解 剖 ・花 粉 ・多 くの embryology の形 質 ・化学成 分など大 部分 の 点で一致, 或は類 似 して い る。 し か し, Elaeocarpaceae は, 厳 密
には, 材 解 剖の細か な点 ・葉 序で異な る上
, 師 部 細 胞の 色 素 体 が蛋 白質の結晶 を含 まない こ と, 子 房 壁 内 樹 脂 細 胞 層 /colleter を 持た な い 点で, 明 ら かに ヒル ギ科と は区 別さ れ る。 こ の
こ と は, Elaeocarpaceae が, ヒル ギ科と近 縁だ が, 独立 した科で ある こ とを示し て い る の で