0 •
Profile of olfactory
tyrosine hydroxylase-expressing neurons in the
bulb of prokineticin type 2 receptor-deficient
mice during embryonic development
Atsuko Kubo, Mamoru Nagano, Mitsugu Chihiro Yamazaki, Toshio Terashima'
Sujino, Koh-hei Masumoto, and Yasufumi Shigeyoshi Department of Anatomy and Neurobiology, Kinki University Faculty of
Osakasayama, Osaka 589-8511, Japan
'Division of Anatomy and Developmental Neurobiology, Department of Cell Physiology, Kobe University Graduate School of Medicine,
7-5-1 Kusunoki-cho, Chuo-ku, Kobe 650-0017, Japan
Medicine,
Biology and
Abstract
Dopamine neurons in the olfactory bulb play essential roles in the maintenance of olfactory function. The maturation of these neurons involves mitosis, cell migration and the tran- scriptional regulation of specific neurotransmit- ters and is an intensively studied process. In our present study, we investigated the embryonic development of tyrosine-hydroxylase (TH)-ex- pressing neurons in the main olfactory bulb (MOB) of Pkr2-'- mice that lack axonal infil- tration from the olfactory neurons (ON) and wild-type littermates (WT) at E16.5, E18.5 and PO using immunohistochemical techniques.
Even at E16.5, we observed two populations of neurons in the MOB of WT mice, one scattered in the primitive glomerular layer and another population of TH-positive neurons in the surface region of the primitive granular cell layer. The number of TH-positive neurons had increased
markedly by P0. Pkr2-'- mouse embryos and neonates showed disorganization of the layered structure, mainly due to the loss of glomerular layer in the olfactory system. The number of TH-immunoreactive cells per slice was marked- ly lower at PO but not at E16.5 and E18.5.
Interestingly, even in Pkr2-1- mice, TH-im- munoreactive neurons were still present in the periphery of the MOB. Hence, it is highly probable that the lack of Pkr2-i- does not affect
the migration of TH-containing neurons from the subventricular zone to the periphery of the MOB during embryonic development but strong- ly impairs the migration of TH-expressing neur- ons or TH expression in the MOB around birth.
Key words : olfactory bulb, tyrosine hydrox- ylase, prokineticin, periglomerular cell, embryo, dopamine, tufted cell
Introduction
The olfactory bulb (OB) is the most rostral region of the brain and is generally divided into two parts. The main OB (MOB) occupies a large part of the OB and receives projections from the olfactory neurons, receiving most of the odor signals, whilst the accessory OB receives projec- tions from the vomeronasal organ and plays the
principal role in sensing pheromones. The MOB has the appearance of a layered structure com- prising a glomerular cell layer, external plexifor- m layer, mitral cell layer, internal plexiform layer, and internal granular cell layer, from the surface to the center.'
Dopaminergic neurons in the MOB play cru- cial roles in the maintenance of olfactory func- tion.2'3 In the matured MOB, dopamine-contain-
Received August 29, 2011 ; Accepted November 15, 2011
25
ing neurons exist almost exclusively in the glomerular layer.4'5 Two types of neurons, perig- lomerular and tufted neurons, are present in the glomerular layer and contain dopamine as a neurotransmitter.4 Tyrosine hydroxylase (TH) is one of the rate-limiting enzymes in dopamine synthesis and is regarded as a marker of dopamine-producing neurons in the MOB.6 TH expression in the MOB depends on axonal inner- vation from olfactory neurons. Deafferentation causes widespread downregulation of TH tran- scription in the neurons in the glomerular layer, suggesting that afferent projections from the olfactory nerve are essential drivers of TH expres- sion37-" Although TH expression in the MOB is strongly influenced by projections from olfac- tory neurons, it remains to be investigated whether all of the neurons expressing TH are dependent on innervation from the olfactory neurons.
The prokineticins (PKs), PK1 (also known as EG-VEGF) and PK2 (also denoted Bv8), are secreted bioactive proteins." Two endogenous prokineticin receptors, termed Pkrl and Pkr2, are members of G-protein coupled receptors that initiate signal transduction by PKs.12-14 Pkrl shows preferential distribution in the peripheral tissues, whereas Pkr2 expression is mostly con- fined to the central nervous system. From previ- ous analysis using receptor transfected mam- malian cell lines, PK2 binds prokineticin rece- ptors (PKR) with higher affinity than PK1, suggesting that PK2 is a stronger agonist of the PK/PKR system undre physiological condi- tion.12-14 Pkr2-deficient mutant (Pkr2-1-) ex- hibited hypoplastic OBs and hypogonadism due to the lack of GnRH neurons in the hypoth- alamus,16'16 whereas Pkrl -f-exhibited no appar- ent morphological abnormality.'6 Pkr2-i- mice fail to form afferent projections from olfactory neurons to their targets in the MOB and therefore lack a glomerulus in which axon terminals of olfactory neurons and dendrites of mitral cells form synapses.17'18 In our present study, we analyzed the embryonic development of TH-ex- pressing neurons in the MOB of wild-type (WT) mice. In addition we compared the OB of Pkr2 mutant mice and control WT littermates to inves- tigate whether loss of axonal infiltration of olfactory neurons affects TH expression in the OB.
Experimental Procedure
Experimental animals
The generation of Pkr2-'- mice is described by Matsumoto et al." Originally, the background of the mice was C57/BL but, with the low sur- vival ratio of neonates, the genetic background
was changed. The colony used in the present study was established by 11 backcrosses with Jcl : ICR mice (Clea Japan, Tokyo). The resulting strain was subsequently maintained by inter- breeding for at least 10 generations. Genotypes were determined using an established PCR method. For comparison, embryos and neonates of WT and mutant mice (n= 3) at E16.5, E18.5 and PO were collected. This study was performed in compliance with the Rules and Regulations of the Animal Care and Use Committee of Kinki
University School of Medicine, and adhered to the Guide for the Care and Use of Laboratory
Animals, Kinki University School of Medicine.
Tissue preparation
The day on which a check plug was evident was counted as embryonic day 0.5 (E0.5) and the day of birth was defined as postnatal 0 (P0).
Pregnant mice were deeply anesthetized using ether and the embryos were removed by cesarean section. E16.5 and E18.5 fetuses were fixed in 0.
1 M PB, pH 7.4, containing 4% paraformalde- hyde (PFA) at 4°C for 24 h. PO neonatal mice were anesthetized by hypothermia and intracar- di ally perfused with 4% PFA. The brains were then removed and immersed in the same fixative for 24 h at 4°C. Subsequently, embryonic and neonatal mice were blocked in the coronal plane and embedded in paraffin. The paraffin-embed- ded brains were sectioned at 4 ,u m thickness in the frontal plane of the entire OB and mounted
on silane-coated slides.
Immunohistochemistry
The specificity of the anti-TH antibodies used in the experiments has been described previous- ly.16 As the cerebral cortex of Pkr2-1- mice does not have any apparent deformities, we deter- mined one plane by using the position of the rostral tip of the frontal lobe. The cutting plane is symmetrical in the right-to-left axis, vertical to the basal plane of the mouse brain, and is in tangential contact with the rostral tip of the
frontal lobe (Fig. 1).
Sections were deparaffinized in xylene and rehydrated in an ethanol series. For antigen retrieval, the slides were immersed in citrate
26
Fig. 1 Section used for comparison. To compare the posi- tion of the OB, we defined one plane The cutting plane is vertical to the basal plane of the mouse brain and is in tangential contact with the rostral tip of the frontal lobe.
buffer (pH 6) and heated in a microwave oven for 15 min.20'2' The slides were then bleached for 30 min at 4°C with 50% methanol containing 1.
5% H202 to block endogenous peroxidase activ- ity. The sections were next incubated with a primary antibody against TH (MAB318 ; Merck Millipore, Germany) at 4°C overnight (diluted 1 : 2000 in PBS containing 0.3% Triton X-100).
After rinsing with PBS, the sections were in- cubated in biotinylated anti-mouse IgG (Vectas- tain ABC kit ; Vector Laboratories, Burlingame, CA, USA) diluted 1 : 500 in PBS at room tem- perature for 1 h, washed with PBS and incubated in avidin-biotin complex (Vectastain ABC kit) diluted 1 : 500 in PBS at room temperature for 1 h. After rinsing with PBS, the sections were washed with 0.05 M Tris-HC1 buffer, pH 7.4 and treated with 0.035% diaminobenzidine (DAB) and 0.05 M Tris-HC1 buffer in the presence of 0.003% hydrogen peroxide for 10-15 min at room temperature. After the DAB reaction, the sam- ples were rinsed with 0.05 M Tris-HC1 buffer, stained with hematoxylin and dehydrated with a graded series of ethanol, immersed in xylene, and embedded in Entellan (Merck, Darmstadt, Ger- many).
Statistical analysis
The cell size and distribution patterns were determined using ImageJ analysis software. To compare cell size, we measured the length of the short axis of the cells. The number of TH-im- munoreactive neurons per slice was counted manually under a microscope. Results are ex- pressed as the mean ± SEM. Differences between groups were examined for statistical significance using two-way ANOVA (analysis of variance) and individual values were analyzed for significant differences using one-way ANOVA Tukey's post hoc test.
Results
Analysis of developing olfactory structures in mice using hematoxylin and eosin stained sec-
tions
At E16.5, the MOB in WT mouse embryos shows evidence of layered structures. In the MOB, the olfactory nerve layer, consisting of olfactory neuron axons, is already established at this stage (Fig. 2A). The external plexiform
layer and mitral cell layer are also recognized, although they are in an immature state.22 The glomerular layer and external plexiform layer are not well separated by E16.5 so the outer most
po1iLl'. •
•
( • *: •
• . ••:‘ 143i=
10;14 • ••:-
i;" •
00111
A: • • 1164
•
• •••:, • sH,‘
' MV
P
•
AP!A:
08.1
":.
'4Nhottl'ff,
't1:: •
'EfNL•+`EG~
.1:
.;•^•;•••••••:,
• ‘& • ;1' 2 • ;
ISO
Fig. 2 Tf
•• •
6•
ti3W8itt*ri.gt
ktslitMik,
1.0
•
• .•
•
a - k • 1
• • k^
1.1.
•
, ^:!‘
Z1C^
•:•. ^•••
6E5:44 tt.
•‘'
••••••.:
-,,,,k,.- • •a•.' • • .
.,• , ...,,,•:I: -, 7. •
..,.!: 4. lil: :: :: ,,i ,.1.1. % .... .: :: : . tt: ;:i'Ll .!.‘ "C:11 i .1.41_ :: ' ... :5; ;.: .44 ... z 1,... I ..::^:. . ..u; . ; I : f l'....f; :. 1 i ..I i i;'... : ,,. . - A.:: ,. . • : s .‘ ' 11 :1; : : : •.• .:.. : : ..' . . , . .: : ; ' .' ;: .1,t.t . . ..,: :1 1 .:1.* ..;; I :, ..• t....' 47 I. 1 :.,: .•:: ;* 6 : ::I 41: : : :: 4.1
*"....• "Tio,loric .., :::,..-fv.,:itv "••••,,ff.4 ,„ It:',.••,..• • • ,l,,,s
i.57,..t.' '. V•*';'''... . .(1 ;,.. %J.% 8%4.4 ' ' - 4•,,. -,-- "....1.
1
...tr.,: .E.-f,..,:t&k....r.i., • ...-• -4 .... -t-,
... ..ft.,,,._. •_:Fr ..-
• .
it
••
•,,
• •• :;.::
t•-E
•;14 os.!•••••: lot*
. '
;1;•et•.:(2' • n
•
••^,, V i•
r. •
Aq-rg.g.
witt. Au.
41141440 otrio
. •••-•
?4,
40,1 i
The OB of control wild-type littermates (WT) and Pkr2-i- mouse embryos. Coronal sections of the E16.5 (A, D), E18.5 (B, E) and PO (C, F) OB of WT (A-C) and Pkr2-1- (D-F) mice. Scale bars, 100 pm.
pONL, primitive layer of the olfactory nerve fibers ; pGL/pEPL, primitive glomerular layer and external plexiform ; pMiL, primitive mitral cell layer ; IZ, intermediate zone ; SZ, subependymal zone ; ONL, layer of the olfactory nerve fibers ; GL/EPL, glomer- ular layer and external plexiform layer ; MiL, mitral cell layer ; IGrL, internal granular layer ; GL, glomerular layer ; EPL, external plexiform layer.
Iv.... •. •••k- :;',.:".:4-•••••• -.• ••Zr. • ':'-.•- ''''-• 2-2
... . , -.--".4.,^••..), ' --,:'..- ....•.:‘;:,,I.*".`)•::;;...c:.-- ,..:4•• z t • - • ..• It t %..:,-.• -6 . • •• 1- 41.ii..•^•'•4.,...:?....ir .:...1 - '',,: v ,..z.,: .44: r ... hi. :;'•;:•3 :•;'11%,:t
..:Arti,r:".• ::-..r,%.; A, ., : • I, A,-f.:..-...,..
, ,?.?,..„..,,,..,.-- .- 7 • • . fe .,(6.1c;A"81:, . '41? 7- ',
• ;•••''..
•*:, .1•\'''' %.1."i4Si.. ' ' ' .q.-•
44,4,' :',711i.'is: •• ." 'tWi•••:4.$41;*'''''' • '11' - .-•-",:,.;t-A...- .,,4...ffi",,,,,
1:4;e1;.f',...4WAW, . • ,ir.t.gAZV":t:‘ '' • •.-f ,, -s4'..' 8 -:
.1;„,..4.• .iii*:, .-s,...--;'''F;.', ',:131' ,..,...":..;5:,:.••,,,• ,....,•'---411't.:; le. . .: f.: •''pi.-,,I - .-..^-!• , , ,. , 4.11i.•:-..:I .P.:;::.ii:i7..`77; : . . • -, '. ., , , - ..., ,:.N.: 3,s-.;.7.--ii... Itil
;:r ..: 2 :kr,z. ..:t-diwz%..t.. . -,.--...•.e''....A496. ,•40:-..;047.4,A27-.3..t., ,z.v.a•l.R. _,,'..Z..• Y•:.;-. .5r-:,ZsT '..*;;'-A:V;i-Iki k -.4 ',. 7.. '..
,Pt.:-::'!!;':•*,;::1'.:.ifiito..4ii.13frl\s- Yti ... , . 47,. ., - ..8,:'....,„ • t mir
, . 1-. , .1::-:,... 4.,-..!q..tY.• -4,...t .L.;,...A. s- -
• i.. *. ‘,i" f.#14:074--;i6tv i..T.a.4%<1: "4 7s .„..:*:..,....;.;.,..„.-ti.f,--..'.3110 ...v . ,.:, `511..^:.,,i.,,,,,.it .
,;2---.1-.1klie:•0444n-'V^-
,.,...,,,,,L,
lb.%:,?..A40;,:.. Atikitc:; • 47. .-ti,,,„:
#. .,,O' „ A *;$:'sq;.in.• ;V:.1":1;;;VAW::
1..:',' ./11;1J...f• .: .41' '..'kirlZaig
It., 1: A'>',.."':". .;1.7,.,;.•;•-:-.,..- *: II:
‘• • . i.'"*... ., -A. , i.,•,i1" • 4.t.--.0' -A- -,•A't .:.:.-..,,., -114,::74,,, , - t
..t....;•::::44_4*...., ,..,.:7•2-tv..,,:.A...,'Pvt:zien.1:.'4 _ t • ..-....-4,11,,,i,:.- ..e;riis;:-,;:i•,-,..,f,i.i.;,.., ,:"..,',44. t'igeRe...440.41 t....c.,:..far 4.-ii=1,4,77.„5:7.:-44.1' PO .,•;,..*t1/41,4, fk..1
)[ ds -.t y pceo rl iot ht earim saeteetsi 0( nWs To) f atnhde
27
layer in OB should be named the primitive glomerular and external plexiform layer, as de- scribed previously.23 In addition, the internal plexiform layer and internal granular layer are also not well divided and the layer is referred to as the intermediate zone.22 In the central area, there is a subependymal zone distinguished by its typical radial cell arrangement that has been reported to contain proliferating neurons.24
At E18.5 of WT (Fig. 2B), the appearance of an established mitral cell layer was the most striking observation. Mitral cells are distinguish- able from other cells by their larger size and were found to be arranged in a line. Due to the formation of the mitral cell layer, the glomer- ular/external plexiform layer became distin- guishable by E18.5.
At PO in WT mice (Fig. 2C), all of the layers in the adult brain besides the internal plexiform layer were distinguishable ; however, the struc- ture of the glomerulus was found to be immature as we did not observe the spherical shapes that are typical of this structure and the internal plexiform layer was not discernible at this stage.
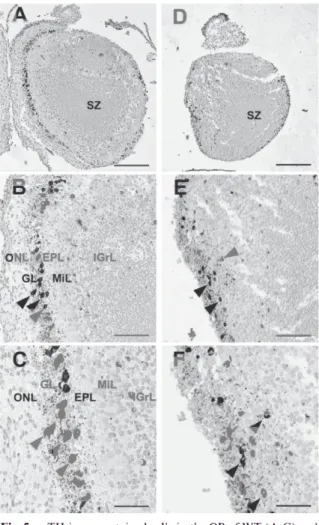
In the MOB of the Pkr2-1- mouse embryo, the layered structure including the olfactory nerve layer was lost at all of the developmental stages examined ; however, a cell-dense area surround- ing the ventricle and a gradient of cell density from the center to the periphery was observed throughout development in the mouse (Fig. 2D- F). At all of the ages, the subependymal zone, distinguished by its typical radial cell arrange- ment, was also observed in both Pkr2-1- as well as in WT mice (Fig. 2A, B, D, E ; Fig. 5A, D).
Cell density was high in the central region, and low in the periphery of the MOB.
Analysis of TH-positive neurons in the OB of wild-type littermates and Pkr2-i- mice
In the MOB of WT mice at E16.5, TH-im- munopositive cells were observed. These cells were generally divided into two populations (Fig. 3A-C). Strongly stained TH-immunor- eactive neurons were found to be scattered in the primitive glomerular and external plexiform layer. Moreover, a few weakly stained TH-im- munoreactive neurons were detected in a scat- tered pattern in the intermediate zone. In Pkr2-1- mutant mice (Fig. 3D-F), TH-immunor- eactive neurons were scattered from the core to the surface, with the exception of the subepen- dymal zone.
At E18.5 in WT mouse embryos, strongly
/if firli,V.i.irr,g'M 1/1,1 : -4
, , ' -12,,-...1c ,...' ,r tifs,...,.ii•,,,, 04tiirt"4 1•4.:
•' , ,:,;).?1r,,':'"-.i'll, '. :,4'..iit'O' .%;:!,"`{:;'''f'''' V. ''' ','.':5:V•:.:4 Y% .
,"t
t--;.T.;',:r."..4.6J:
,."'!?:•' _11,` '''.`, Tlif t 4,:;,,,...
,11'i,zi•i':. ' ',' ;.:,''',.' ."' r';qr , .01' '''s
:54! ,,,,::" -••• '''',1 ' 'IV i.
v.,: •
,1•'.'fr Pk.
' •-C-:,...'''' Y'L :?'''-;:V.,':.:3:;1 '`, .1 •V•1'.
_ A' e.,... . • ...• , t .1., :k
„to
...,...,..,...,...,„„„, ...,,_... .,,,,..... ...., ...„._).„4::0,,,,_ . ...„..„‘„ .
..s.„...,,,
.,..„
„4...::;,.::.../.:
.
...774...,, ...:,....,
ov.... ..,..i• ‘-iidg.;-fr:1=:?r3/41ej...14..T ; ,, •oi;;;. ,',..,-/;:,„.:,,,:;.,, ,1'4,,,i', ,• • :,,,„15
AftidiY11,141‘1
, , i',4r,„'„,:-.; ' ,-, • :::t...„', ,.-i% 1'3,,
..-- ,, ,,g,,,,,-;.•,,, zky. , ,,,tk, :705
1.•.,„,,..,-,,,,..,i,i,.,:,,,„....,,v,„„:;..„,..,-,.,,,„,,,,,,, , rj;u45-:.:
,.--A::'
,:st, To•4 .',,-:' ,,t-,-;--;:-,,,, ,'"-eF .`,. k t'•
.,:,,:t..4 -,71,': ,ls-,,--,'.-3A ,,,f,•.%.;,,,, '4
,
'"' ', .-, .."1 ' .2 :','::•T ,i,.... -64440 11,7&:-..,,,'p, ,,,,, , .-.):: ( - "4-,'•.17
"4
01g 7,t10-^
. • 1 • 1..! ; • 41 A:::
...) ;6. . :,!•04? ,7e.j,.,,,,,,,,f.
•Or
•..i.- .7kt...;-.1f,t:'';-t°t'.'
y'tri••.-/•• • •••,• 3d1.4:1 / ':-..: t t,'„14•H*, „€ ,,,r , •,.„6,51,;,,e.tf:,.,„..-/-,I.:ti..„1.7,1,. ,
(.l'n-**,1 i''r 7 i.ily :4.:;i4:f•-•1•:‘. :..:.:-''....:::i.)...?
ittii,;,-,..,_ i, ';',,^':ii • ;",„. .p..,.7, 14,',..iiii;411.;•.• , ,se,?..,,stiot:". '-.1/,!,•::.1'.!_‘,..C‘1:4.1.•...,47a. ,..e. .r.:::
''
J , - i
,,...,"‘,kAfaUk'tur'ALtiuskii,...--.4.'.1
'.{i' .;
,(.;4;,:i',;,1111.•-.;-91 > YP'4,,' r.*1
, "N' •I ;•.' 4 : iiaeic.”- .,, '‘;‘203,i,':^sa, Ult.., fialiY:e4.1tolio., • VIK3,-4---i. 'ib ''',rfb"Y;+' ,',' ..;i''' e '',4- v;'4/4" k -i x
4'ie7-'140t 7 ,
Ditiip -z.- 'i • 'Air ';:,'', '., ,..„. ' ' ,.:
aa~il:' '„1„1 1 Ir ..:1*3 '. .,' ,',.: ', ' iZ• .% 14 . I"
5-
r. pffilL.4 ., ..;,,,' if, • , ,--; • 3, , .0 i1I.:.4„,'3,"•V4.1.3'4(..,','':'''''' ' w -.„ - 0 1
44..r.,4 , , , , \ •iit 0
,:,„ I
,,,i) I' t • II, .11C' .1 . , .',,, ' 6 .. , ' liull, ' ..4_\
Fig. 3 TH staining in the OB of W' F) mice at E16.5 Scale ba
representative immunoposit
rt't-2..,71§tt ft•I
i?.:•••;
7.1.;
1•,e•it".,•••if,',
•
it •
I '
tt'•
••:‘, 41.
• ta•
' 45..ls''' '''.'1',1'=Yt';',',' '1 ",,''''. :14';`,. 11, '''%•,:. ...i.,*/...':''.r.: .:-%''.?,;;;;;4.4.2? '' )'k •AiSair';utAL"aUtliii '',:‘: 4 . . VtioY,:.4 ..*:•YA.':.4.ilii:.Ziali+r '. sd'
0-‘' :0'....j';‘,X.-,:i•;1.9.4.,7.,..70/ ) 1; 7;4'; , "ri,40:1774be?,1..07,:,s•e.';',.4. f' ",,s, 4,;••••••iy:,,..,4
144" %.4,%,;; ;.,,,,di . - 'ft,74. f,,...; --vitar,,.., l' _ Ke••-4. 7..:-.-1"7,,,, $ ,)f. „-i;': , .:" . ill -410*,. .f.';11,0,.,•:,a,,,;,,
,,..",..,,.,-,„.,; .,,,,7e.•,.,,,,*re.
....'" ',17-?..-..;(4.4 -- ,.:1-,; ,, , ,,-:!,.-'7;:r.e.,•/, ,,,,, ; . , ,.,,,, 1 7—r.f.;.4.,...ii ' , . , ;IL l'Ffrf"'•,7„:.t:' ...,.... • -4, „4,,," !,-4-1.,„T, “,..e-40. ‘21.: • i-, c
Or
4:e114140.:4;::.,%, •• ,131, ; ;6;',.'
irpt,,*,::.4. ....s, I. .11t1; -:1.;41/
7 . **'41,- 3,' ? iV ;!•44V,',- i ; '`, .,'^ :141„?4,.4171; r, 41% i-''-'41 . , • •-') .tPlielr,:, ' c' .1 , ' 1,dily '''''' ' cf.../..c._1,!' '4
4".o.,,,,e_. 77 - 'ef
, 3 TH staining in the OB of WT (A-C) and Pkr2-1- (D- F) mice at E16.5 Scale ars, 250,um (A, D) ; 100
Arrowheads indicate
representative immunopositive neurons. Abbrevia-
tions are as listed in Figure 2.
SZ,;5
stained TH-immunopositive neurons were found to be scattered (Fig. 4A-C). Immunoreactive
fibers were barely detectable, but a small number of neurons showed weakly labeled neural fiber extensions. In the internal granular layer, a few moderately stained small neurons were detected.
In Pkr2-i- embryos, TH-immunopositive neur- ons were found to be scattered in all layers apart from the subependymal zone, in a randomly distributed pattern that was similar to that obser- ved at E16.5 (Fig. 4D-F).
At PO in WT mice, most of the TH-im- munopositive neurons were localized in the glomerular layer and the majority of TH neurons were arranged in a line in the glomerular layer (Fig. 5A-C). The neurons were larger than at E16.5 or E18.5 (Fig. 6A-C). Weakly labeled smaller neurons were also recognizable in the granular layer. In Pkr2-1- neonatal mice, TH- immunopositive neurons were scattered and did
28
, .
,• .1.5s, '',-,, Al- - ---4.--- ,• ,, <A.,,7v11v.= -",•:,7,.;:iii, - . :3, 1 Th --::',: , D ...,,...?,,,,„..,., . :-:, , '.::::::::,':/-:::::::,,' -',-, •••,,-, ‘-,..-"-;!',.--,4 ‘-,..,:-,..-_:.!::' '''•!. ,,, c- Wfw,:isii64.;'-''':" c'' ' ''','Ii.,'-,, :* ;,.•••,,,;,:s.:17:.:1'. :: :- ‘:: •
(
4,-,..,..
•'-; - i...,-,t:,;:,
%.4,•,i -.,‘ , ','"q's .-,, ,;:e.;.,...-;:;., '''.. '-Z,,',- >...,,7,4%. '' • l':' i:'-‘ I % M.:`''''• ‘,:;-'..4-..-.:,.'.
i'.'..,• ,' ''..--,•'':7,-•:"';',`f4.: ----..4-;',.•-:. : - - '.'‘,.-, -
sz - , - • , „3,• , • , ,.:!"--) - :, .' , -,,,,,,,f-. • t: ::'.'f''..-,..-• :-;,- , „es,: ,..,-A, ' ..: • , ,-.,-./i7,3,..,A,,,,ip..,,- ... ,,,t .,;,.: •,..•, .,•:-..:r.w'-:" .-,„..4,,,•-,!..:i ....P ,-.• , -: s• ,--- - ' ^ *', -'''' t.: . - -, i::'':'. • ,., - ,•-• ::,- 4.-,.. -•,....• .f. -1„
1,1', ,>),;,",.:r-•..,,," ',..-'-',.ft,,-1.':-;':".' --2---o's--',-;,--;':f:.:. ,,e'4.., ,4".i.. , . , : :,•-,!,,;,,,,,:..,:,-.;,,i, ''.•".,`'-',-,,,;,.: ..,;',-",r-.•:!=i4:1,-1.,,V:-„f•:- ''''''':;•:-.„. -":,-.,F. sf,,.:.,„,..;,.--.c.:* ,- ,- " A
- .
k , , ,. A • --• .,----;-).: ' r--- ", ,s,, '
L.- ., 2 •'-- :1•3.. Al•-• - .•• ,t;,,,,t,,,,A, -?,.-••.-.•••-./1• ''4,100. , '4•1-'•-:;N'• i+ •.. .- , , ' ' ',..,)', ..:•"' '.f '-4.• .:. 44,, . ir.s..; • ' -,4- igir• , .
,442.44..4.‘ 41„frit;,.. ;Ii....f • 4-''.0 -4'4 • ! Z' '''" *2'• tcl : • 't.:;k1; ' - ' 16. "rt'.1.-."r ; &4v:.: ., .1, .,.4-...,, ,:,,,... 4,•.;
t,P. • 41- -X "5,:''.. ••^ 4 %'•V *4e' ''li 0' 4%?: ,,,.,,,-,v.:,....•.,,lte: ..s.* •• 4 ,••••:".. •'. • VI :• t .• 1••••^••• ", '• ,. •*i • ' . '''''•-•t• '''' '..4 .,4 ,•••• •4tte et • •I'•*41-....s' • • • ' • .4141..,;7' - t• • a'''' ••••••:' '' .44' •;11: n'' ..., ' vi• tt :0', • 1 ' otte
, d . . • f., „. •,,‘„c..4,,I.-...A....-1 0 *,,,, l' ...., ; ,,,,,.! : • •IA•:. .., .11',V f . " : ,',... .,44....44.4. ....4,f. 1 i "...-1,
;4. ,'y. ,',YrdLd *.;.1;.°.:,:w.':,itials;.;:,,, 1. •-.: ,A t -; -...-4;..-4',.;.,5,,-e.t...f.-2,1
.,,, ji„, „VI_ ';',!',....:...-'-;y:::;:-,;r.,i'il„,,,....,,, .. ,.:( 13:y.Z-,^fy,,-..,....,11:v.ittif.;:ri,l,-,,,:,, ' , ...0e, ;$41 I ctiP_Cgi^' •^•46.1:;-:A-$: t:::04. .7,i, 7.1--,.:*;,,?.•-../1; "lv4:4
,-,..iiitT;$!:t`tV:',31v1,11
T .,,':, 4. - •.•s.:;(4,:11;;',A,:-.,,14
1 ..:-J ; ,,-;,,• -.1-;i: tA,0_,-; • . ' , • (-.-:,..,•vi,t,‘., .%•,*,,-.q.:;•,;.•::,/.
-.:.•-4-....T(.3,*
....',:'?.‘-' -al•
,.,^, , • ,„,,, ,,,,,,,,.,„,,.„. ,. ..,, • 0, ,.-ei
.t..\ 1 54:: fr- ' ' ...“ h....',', -. 71r:',.%'F.;ti''•:1 ' 4 V."‘ it.!i,...„1,1 ." ' s .e •fr,. V 1 e -4? ;i:;..!Vfc7Iii 7 14-‘'h .,14V4 alk 4 ',. i " -, i' i ,:0;;;•4::(74..Z: i.a.-1- k • ' -'4:' -' ' t: .-14,a,f...sy,,,,..1 ^ •:: .. • ' •• t '. •4 .,41%. ... V..• 1 • ,....'•Air:?:.4.4411 :1 1 .1 4i; ..1.:%4:,1*.,'' '•
•' I -,:',,,) '',4%,A .10 . , tri!. ,i.: ,,e ;A,- ,• ix "r • 41016 r; ,r
. ,, , -,,,,-1 , 41 EP Gil 4 .,e • r MiC
,, . • . _ ,, ., ,l'' 1 ta . *;0,
, m 0- , '' t :q411?esti,:.
4 i.4.5.1 . 6 A it'-'-' 5 : ' '4. r V11:-It' IC" Lyle f -11
y...-f. Af.,:, 1$ 4p. , 1 1:) 4" '°" A • 511...!..V, 1 1 t. .. .-. ,,. ,,6,.._, ' •,, -, , e #1,... .11 . e ,.• _ 1,-,4
, 0 t . ... !.*...) '' ) ' ) • • • - n• '', '..., • ' . i of '''" .,, • ..1. • vv. 0 •
4R, 1P• , ."' ; ' , ! 4. 0-, . , „ 4,1 • ..% tt. l' 4 :°C.-4 )`. -4 ' $4*,; ';':.- ,. 4,14i. a - 0 • ' *.e.' -. '. N ":!eitit 01
,,,,,- A ,.i • ..,,...‘. ) „ , ,,k- * Naratt ikt . a At , '
Ili- 'Tr' A•i11,.. )., P ' 1 '.."..' i,, %. ''-'', At 'X ' 11.,) , -.6. " • - . ' • 4 - t if briele71
Fig. 4 TH-immunopositive neurons in the OB of WT (A-C) and Pkr2-i- (D-F) mice at E18.5. Scale bars, 250 pm
(A, D) ; 100 pM (B, E) ; 50 prn (C, F) Arrowheads
indicate representative immunopositive neurons.
Abbreviations are as listed in Figure 2.
d' ,,,...„... . D .7 ,..-,.:,... .
" '47 , • ' • ' .,-.1ss e ,.,,,,..7..:7-,-4:,,--,..7,...
1r
,., .:,,.„...,...,...•':,'
4., ,..:.-....?„„se., ,.,...., ii,„.,,,i.
.6 '.' '..r:4... • li-.1,:z..t*"...,,,.: ..- ...f,,.....-...:,‘;,:r :,i'..:,..':::-,....i„ . ;11-..i,.,„ -,,•
4A--...:.,.., ::: ..,:::-..f . :. • pi ,,,,,,,,,,:f.,,,,,,viz.4
4.1.,,„ . , • . * . • , , , • , ,,,, .,
,....t4,:-..-..,- „,..,...,,•‘;:•7,-: , . ,,,,.:, • ,,,,,..:, , .:-,.,.
•
4, `..,! :, : ,,,,, i , : ,,,, ., ,,,, •
1
f. c7.-,,..... , , !,',,;•
.,."-..e,
- - ,..t. . it)ie:.:••,*,, , ^ .., .''
NV.;VERL.I.:: ,, ,.. ., -;,,, IGrL.,. „:,,;;), .1 ,,,,At
. , „-,,, • 1 46 ,.
..,,,,1
,- ,. - ,,r.,„t.,..' : 1:•11;; ' '',..'' •
Ii.,•',
1 , ,, , , , ,... • ,,,. 110 ,41,1 .. .1' -AN
_,.. ' .' 4 ":::LIHIY,::
N;I: ,. .1.,A . ,..,4_ ....,
10.2'....:Olizesi,:".).,;:,:....7".717,-1 r ' Air:,
.... ,...
..,,,,,,,, ,, ,„ •.,..., .0. ,.4 rs,, •,.• ', 4. ,, , • , 4, ,..,
**i ., 0 ,,,,':.,•• . ;., . ,, -? -.4 , . 1 t•I'il.'i. 14-...- ). ''''' 4 /
...ca.. , MiL ' - ; '4 V 4), '..,;4 .,'•''...•s i ' : ' .6.
1116r1.2.
' ^ ,• tg/104. ., , i•V ,' 11, • o 1.'41.: --.5 . .. , • ', .1 ° • 4
, ^S r IV, ,j, , . , ,
^ T.‘,'**‘
4.00,.ridz-,,, . sPri,, A
lk , .1* err ., •-. , % ..a.'1 .!;is, J'..'r -, ^ ' . .: 1 - :
•,1, • 41; -,,, ' : • .,?`, . ' ; • ; ' * ' Oa ' : qt.') „it : I . A • , e; , ...," ii• - .11 ' ' ''', AK ' P 'I ' ,.. ) A ' rige t' 'sgf,:,10. ,"... n '6 A •,,,, • !,: a.. , .., .... .
..c.,.' ':k, , I f • y.... „ 1...;14.4.4-'," "Ai,* : % . • -.. • , • ,..*• • A " r '411iii-.;: .r, , , +, s' '., • . ,,;:. p
C.`eSi .. ° ... Et' 4., ...4 ' *, , d •;1 4" ;44 , 4 s,'' ' 4, ' 16, Fig. 5 TH-immunostained cells in the OB of WT (A-C) and
Pkr2-i- (D-F) mice at P0. Scale bars, 250 pm (A, D) ; 100 ,um (B, E) ; 50 ,um (C, F). Arrowheads
indicate representative immunopositive neurons.
Abbreviations are as listed in Figure 2.
not show a linearized arrangement ; however, most of these cells were generally situated in the periphery of the MOB, close to its edge in an area that should be the glomerular layer (Fig. 5D-F).
No apparent increase in cell size was observed (Fig. 6D-F). The glomerular layer was not easily detectable in PKR2 mutants because the cell- sparse region, the external plexiform layer, which is recognizable beneath the linearized TH-im- munopositive neurons in the MOB of WT em- bryos, was lacking. In both WT and mutant embryos at P0, TH-immunopositive fibers were found to be abundant. WT-immunoreactive fibers were mainly extended to the outer side of the cell bodies (Fig. 5C). In contrast, TH-im- munoreactive neural fibers were found to be extended randomly in mutant mice (Fig. 5F).
Statistical analysis of cell size and number We performed ANOVA to determine whether there were significant differences in the number
and size of TH-immunopositive neurons per tissue section between WT and Pkr2 mutant
mice. Using Tukey's multi-comparison analysis, we did not find any differences between the number of TH-immunopositive neurons in WT and Pkr2-1- embryos at E16.5, but a significantly larger number of these cells was found in WT embryos both at E18.5 (P <0.05 ; Fig. 7A) and at PO (P < 0.01 ; Fig. 7A). Using ANOVA, we found that the size of the TH-immunopositive neurons was significantly greater in WT at PO but not at E16.5 or E18.5 (P <0.001 ; Fig. 7B).
Discussion
In our present study, we found that TH is expressed by E16.5 in the primitive glomerular and external plexiform layer and intermediate zone of the mouse embryo. This finding is consistent with previous studies in mice that
29
V A
El B
•
-II
=kb
• —
_ .
_
ts-41
Table 2 Average size of TH-immunopositive
neurons during development (Mean±
SEM) 60 neurons in three slices (normal 3 ; mutant 3) were measured for statisti- cal analysis.
Age
Mean length (,um)±SEM
WT Pkr2-i-
• •
111) OR
•
E16.5 E18.5 PO
6.56,um±0.11 6.79 ,u m± 0.20 9.06 pm+ 0.29
6.54pm±0.1 6.22 ,u m± 0.20 7.33 pm+ 0.30
op
-4 E113 .5
Fig. 6
Table 1
' 4
Highly magnified TH-positive neurons in the OB of E16.5 (A, D), E18.5 (B, E) and PO (C, F) of WT (A- C) and Pkr2-i- (D-F) mice. Arrowheads indicate representative immunopositive neurons. Scale bars,
10,um.
Averaged numbers of TH-im-
munopositive neurons per slice during development (mean ± SEM) Sections were counted in the section of OB (nor- mal 3 ; mutant 3).
Total number of neurons per slice
A
B
E
(73
90 80 70 60 50 40 30 20 10 0
^ +1+
E
.c
as
71,
U
10 9 8 7 6 5 4 3 2 1 0
E16.5
El -/-
E18.5
* p<0 05 PO
** p<0 01
Age
WT Pkr2-i- Fig. 7
E16.5 E18.5 PO
27.3+ 3.8 28.7+ 1.9 75.3+ 11.8
25.3+ 2.9 30.7+ 1.8 37+ 1.1
have shown that TH mRNA and protein expres- sion are detectable by E16.5 in the primitive glomerular and external plexiform layer and outer portion of the intermediate zone,25'26 pro- viding a clear contrast to the observation in rats that TH-immunoreactive neurons are recognized in the MOB only after birth.27,28
TH-immunoreactive neurons showed virtually no increase in number from E16.5 to E18.5 in our present analysis but demonstrated a marked
E16.5 E18.5 PO
*** p<0001 Cell number per slice (A) and length of the short axis (B) of TH-immunoreactive neurons in the OB of WT and Pkr2-1- mice. Values are the mean ±SEM (n=
3). *P <0.05, * *P <0.01 ; Tukey's multi-comparison analysis (B) values are the mean ± SEM (n=60). ***
P<0.001.
increase by P0. Further, a difference in the number of TH-immunoreactive neurons between Pkr2-1- and wild-type embryos became apparent at P0. These observations suggest that two types of TH-immunoreactive neurons with distinct developmental processes are present in the OB ; one appearing by E16.5 and the other around birth. Previous studies have also demonstrated that two types of dopaminergic neurons, tufted
30
and periglomerular cells (PGC), are localized in the glomerular layer.4 In mice, tufted cells develop between E 13 and E1822'23 locally in the subventricular zone (SVZ) of the primitive OB, whilst PGCs in OB develop later in the SVZ,22'23 and appear in the glomerular layer around P1.22,23 Hence, it is probable that TH-expressing neurons in the glomerular layer of embryonic OB at E16.5 are external tufted cells,29 and that a rapid increase in PGCs follows their develop- ment.
Pkr2-'- and WT mice showed no difference in the number and cell size of TH-immunoreactive neurons at E16.5, suggesting that the mutation hardly affects the phenotype of TH-immunor- eactive neurons by this age. The Pkr2-1- mouse phenotype includes a complete loss of axon invasion arising from olfactory neurons into the OB 1546 In WT mice, axons from olfactory epith- elial neurons invade the OB from E1430'3' and begin to form synapses in the glomerular layer.17 Furthermore, in the glomerular layer, the num- ber of synapses formed between olfactory neuron axons and mitral/tufted neurons markedly in- creased from EIS ;17 therefore, it is highly prob- able that TH mRNA and protein expression at E16.5 and E18.5 in the MOB are independent of the innervation of projections from olfactory neurons. Taken together, the present findings suggest that mitosis, migration, and TH tran- scription in TH-immunoreactive neurons are maintained even in Pkr2-1- in embryos.
In contrast, the effects of a Pkr2 mutation became apparent around birth. The number of TH-immunoreactive cells increased markedly around the time of birth in WT mice but not in Pkr2-1- mice. What causes the difference in cell number between WT and Pkr2-1- mice ? Most of the increase in neurons in WT mice is possibly due to the increase of PGCs that arise later than external tufted CellS.23'32'33 Previous studies have shown that cell proliferation is not impaired in the SVZ of cerebral hemispheres in pkr2-1- mice.15,34 In WT mice, neurons expressing Pkr2 are observed both in the SVZ and rostral migra- tory stream (RMS).35 These earlier findings together suggest that Pkr2 is necessary to enable primordial neurons to leave the RMS to a more superficial position of the OB. It is therefore possible that, in Pkr2-i- mice, neuronal migra- tion from the SVZ via the RMS to the OB is impaired due to the lack of Pkr2 ;15'34'35 how- ever, whether dopamine neurons in the OB
express Pkr2 remaines to be elucidated. One other possibility is that a decrease in TH expres- sion is due to the suppression of TH transcrip- tion, which, in PGCs, is also regulated by un- known factors possibly from the olfactory nerve layer. Previous reports demonstrated that dener- vation of olfactory nerve projections3° and naris closure7'36 produce a dynamic decrease in TH expression ;7'36 however, in both of these cases, the dopamine neurons survive and localize in circumference areas of the OB, which suggests that migration from the SVZ via the RMS to the periphery of the OB is preserved even after loss of afferent projections from olfactory neurons.8'37 Further studies are necessary to elucidate the fate of dopamine neurons in the OB of Pkr2-i- mice.
We have previously reported that Pkr2-i- mice exhibit hypogonadotropic hypogonadism."
Therefore, it is possible that the hypogonadism have some effects on the morphological abnor- mality of the OB. However, a previous study suggests that it is unlikely that the hypogonadotropic hypogonadism contributes to the abnormal formation of the OB. In the
Ebf2-i- mice that also develops hypogonadotropic hypogonadism, the authors found no abnormality in the OB.38 The finding suggests that the hypogonadotropic hypogonadism in Pkr2-1- mice was not involved in the formation of the abnormal OB.
Interestingly, TH-immunoreactive neural fibers extend from TH-immunoreactive neurons in the deep layer of the glomerular layer toward the outer region of the MOB. This suggests that some attractants are present in the outer superfi- cial layer of the glomerular layer or olfactory nerve layer. In Pkr2 mutant mice, TH-immunos- tained fibers were observed to extend randomly
around the neurons, suggesting that the appro- priate axon guidance or preservation of axonal architecture of the dopamine neurons requires input from olfactory neurons and, possibly also synapse formation between olfactory neuron axons and mitral/tufted cells.
Acknowledgments
We thank Dr. Shun-Ichiro Matsumoto for kindly pro- viding the Pkr2 knock out mice.
We also thank Prof. Toshio Kosaka for important discussions. This research was supported by a Grant-in- Aid for Scientific Research from the Japanese Ministry of Education, Science, Sports and Culture of Japan.
31
References
1. Switzer RC, Johnson JI (1977) Absence of mitral cells in monolayer in monotremes. Variations in vertebrate olfactory bulbs. Acta Anat (Basel) 99 : 36- 42
2. Cave JW, Baker H (2009) Dopamine systems in the forebrain. Adv Exp Med Biol 651: 15-35
3. Davila NG, Blakemore LJ, Trombley PQ (2003) Dopamine modulates synaptic transmission between rat olfactory bulb neurons in culture. J Neurophysiol 90 : 395-404
4. Halasz N, Johansson 0, Hokfelt T, Ljungdahl A, Goldstein M (1981) Immunohistochemical identifica- tion of two types of dopamine neuron in the rat olfac- tory bulb as seen by serial sectioning. J Neurocytol 10 : 251-259
5. Jaffe EH, Cuello AC (1980) The distribution of catecholamines, glutamate decarboxylase and choline acetyltransferase in layers of the rat olfactory bulb.
Brain Res 186 : 232-237
6. Specht LA, Pickel VM, Joh TH, Reis DJ (1981) Light-microscopic immunocytochemical localization of tyrosine hydroxylase in prenatal rat brain. I. Early ontogeny. J Comp Neurol 199: 233-253
7. Ehrlich ME, Grillo M, Joh TH, Margolis FL, Baker H (1990) Transneuronal regulation of neuronal specific gene expression in the mouse olfactory bulb. Brain Res Mol Brain Res 7 : 115-122
8. Baker H, Kawano T, Albert Vet al. (1984) Olfactory bulb dopamine neurons survive deafferentation-in- duced loss of tyrosine hydroxylase. Neuroscience 11 : 605-615
9. Baker H, Kawano T, Margolis FL, Joh TH (1983) Transneuronal regulation of tyrosine hydroxylase expression in olfactory bulb of mouse and rat. J Neurosci 3 : 69-78
10. Kawano T, Margolis FL (1982) Transsynaptic regu- lation of olfactory bulb catecholamines in mice and rats. J Neurochem 39 : 342-348
11. Li M, Bullock CM, Knauer DJ, Ehlert FJ, Zhou QY (2001) Identification of two prokineticin cDNAs : recombinant proteins potently contract gastrointestinal smooth muscle. Mol Pharmacol 59 : 692-698 12. Lin DC, Bullock CM, Ehlert FJ et al. (2002) Identifi-
cation and molecular characterization of two closely related G protein-coupled receptors activated by pro- kineticins/endocrine gland vascular endothelial growth factor. J Biol Chem 277: 19276-19280
13. Masuda Y, Takatsu Y, Terao Y et al. (2002) Isola- tion and identification of EG-VEGF/prokineticins as cognate ligands for two orphan G-protein-coupled receptors. Biochem Biophys Res Commun 293: 396- 402
14. Soga T, Matsumoto S, Oda T et al. (2002) Molecular cloning and characterization of prokineticin receptors.
Biochim Biophys Acta 1579: 173-179
15. Ng KL, Li JD, Cheng MY et al. (2005) Dependence of olfactory bulb neurogenesis on prokineticin 2 signa-
ling. Science 308: 1923-1927
16. Matsumoto S, Yamazaki C, Masumoto KH et al.
(2006) Abnormal development of the olfactory bulb and reproductive system in mice lacking prokineticin receptor PKR2. Proc Natl Acad Sci U S A 103: 4140 -4145
17. Hinds JW, Hinds PL (1976) Synapse formation in the mouse olfactory bulb. I. Quantitative studies. J Comp Neurol 169: 15-40
18. Hinds JW, Hinds PL (1976) Synapse formation in the mouse olfactory bulb. II. Morphogenesis. J Comp Neurol 169: 41-61
19. Barrachina M, Dominguez I, Ambrosio S et al.
(2003) Neuroprotective effect of citicoline in 6-hydrox- ydopamine-lesioned rats and in 6-hydroxydopamine- treated SH-SY5Y human neuroblastoma cells. J Neur- ol Sci 215: 105-110
20. Boon ME, Kok LP (1994) Microwaves for immuno- histochemistry. Micron 25 : 151-170
21. Suurmeijer AJ, Boon ME (1993) Notes on the appli- cation of microwaves for antigen retrieval in paraffin and plastic tissue sections. Eur J Morphol 31 : 144-150 22. Hinds JW (1968) Autoradiographic study of his- togenesis in the mouse olfactory bulb. II. Cell prolifera- tion and migration. J Comp Neurol 134: 305-322 23. Hinds JW (1968) Autoradiographic study of his-
togenesis in the mouse olfactory bulb. I. Time of origin of neurons and neuroglia. J Comp Neurol 134: 287- 304
24. Bailey MS, Puche AC, Shipley MT (1999) Develop- ment of the olfactory bulb : evidence for glia-neuron interactions in glomerular formation. J Comp Neurol 415: 423-448
25. Bulfone A, Wang F, Hevner R et al. (1998) An olfactory sensory map develops in the absence of nor- mal projection neurons or GABAergic interneurons.
Neuron 21 : 1273-1282
26. Harrison SJ, Parrish M, Monaghan AP (2008) Sa113 is required for the terminal maturation of olfactory glomerular interneurons. J Comp Neurol 507: 1780- 1794
27. McLean JH, Shipley MT (1988) Postmitotic, postmi- grational expression of tyrosine hydroxylase in olfac- tory bulb dopaminergic neurons. J Neurosci 8 : 3658- 3669
28. Matsutani S, Senba E, Tohyama M (1988) Neurope- ptide- and neurotransmitter-related immunoreactivities in the developing rat olfactory bulb. J Comp Neurol 272: 331-342
29. Singh DN, Nathaniel EJ (1977) Postnatal develop- ment of mitral cell perikaryon in the olfactory bulb of the rat. A light and ultrastructural study. Anat Rec 189: 413-431
30. Lopez-Mascaraque L, de Castro F (2002) The olfac- tory bulb as an independent developmental domain.
Cell Death Differ 9 : 1279-1286
31. Blanchart A, De Carlos JA, Lopez-Mascaraque L (2006) Time frame of mitral cell development in the mice olfactory bulb. J Comp Neurol 496: 529-543
32
32. Altman J (1969) Autoradiographic and histological studies of postnatal neurogenesis. IV. Cell proliferation and migration in the anterior forebrain, with special reference to persisting neurogenesis in the olfactory bulb. J Comp Neurol 137: 433-457
33. Bayer SA (1983) 3H-thymidine-radiographic studies of neurogenesis in the rat olfactory bulb. Exp Brain Res 50 : 329-340
34. Prosser HM, Bradley A, Caldwell MA (2007) Olfac- tory bulb hypoplasia in Prokr2 null mice stems from defective neuronal progenitor migration and differenti- ation. Eur J Neurosci 26 : 3339-3344
35. Puverel S, Nakatani H, Parras C, Soussi-Yanicostas N (2009) Prokineticin receptor 2 expression identifies migrating neuroblasts and their subventricular zone
transient-amplifying progenitors in adult mice. J Comp Neurol 512: 232-242
36. Baker H, Towle AC, Margolis FL (1988) Differential afferent regulation of dopaminergic and GABAergic neurons in the mouse main olfactory bulb. Brain Res 450: 69-80
37. Stone DM, Wessel T, Joh TH, Baker H (1990) Decrease in tyrosine hydroxylase, but not aromatic L- amino acid decarboxylase, messenger RNA in rat olfactory bulb following neonatal, unilateral odor deprivation. Brain Res Mol Brain Res 8 : 291-300 38. Corradi A, Croci L, Broccoli V et al. (2003)
Hypogonadotropic hypogonadism and peripheral neur- opathy in Ebf2-null mice. Development 130: 401-410