Title Studies on the Diagnosis of Schistosoma japonicum InfectionUtilizing Real-time PCR and ELISA( 本文(Fulltext) )
Author(s) DANG TRINH MINH ANH
Report No.(Doctoral Degree) 博士(獣医学) 甲第552号 Issue Date 2020-03-13 Type 博士論文 Version ETD URL http://hdl.handle.net/20.500.12099/79353 ※この資料の著作権は、各資料の著者・学協会・出版社等に帰属します。
Studies on the Diagnosis of Schistosoma japonicum Infection
Utilizing Real-time PCR and ELISA
(リアルタイム
PCR 及び ELISA を応用した日本住血吸虫症の診断法に
関する研究)
2019
The United Graduate School of Veterinary Science, Gifu University
(Obihiro University of Agriculture and Veterinary Medicine)
i
CONTENTS
Contents i
List of Figures iii
List of Tables iv
Abbreviations v
Unit Abbreviations vii
General Introduction 1
1. Schistosomiasis 1
2. Schistosoma japonicum 2
3. Disease control 2
4. Schistosomiasis diagnosis 3
5. Objective of the present study 5
Chapter 1
Use of real-time PCR for the assessment of egg burden in the organs of mice
experimentally infected with Schistosoma japonicum 8
1.1 Introduction 8
1.2 Materials and methods 10
1.3 Results 13
1.4 Discussion 14
1.5 Summary 16
Chapter 2
Analyses of the expression, immunohistochemical properties and serodiagnostic potential of Schistosoma japonicum peroxiredoxin-4 22
ii
2.2 Materials and methods 25
2.3 Results 31
2.4 Discussion 33
2.5 Summary 36
Chapter 3
Application of multi-epitope recombinant proteins for serodiagnosis of Asian zoonotic schistosomiasis 45
3.1 Introduction 45
3.2 Materials and methods 47
3.3 Results 51 3.4 Discussion 53 3.5 Summary 54 General Discussion 63 General Conclusion 67 Acknowledgments 69 References 71 Appendix 1 83
iii
LIST OF FIGURES
Fig. 1 Life cycles of S. mansoni, S. haematobium, and S. japonicum………..6
Fig. 2 Scanning electron microscopic image of a pair of schistosome worms………...7
Fig. 3 Relationship between tissue egg count and Ct value in the liver……….19
Fig. 4 Relationship between tissue egg count and Ct value in the spleen at 18 weeks post infection………20
Fig. 5 Histopathology of liver tissue sections of S. japonicum-infected mice……….21
Fig. 6 Gel electrophoresis and SDS-PAGE of SjPrx-4………38
Fig. 7 Amino acid sequence alignment of SjPrx-4 and SmPrx-4………...39
Fig. 8 Different expression profiles of the SjPrx-4 gene at various developmental stages of S. japonicum……….40
Fig. 9 Antioxidant activity of SjPrx-4………41
Fig. 10 Immunolocalization of SjPrx-4 in eggs and in S. japonicum adult worms………...42
Fig. 11 ELISA results with SjPrx-4, SjTPx-1 recombinant antigens and combination of rSjPrx-4/rSjTPx-1……… 43
Fig. 12 Kinetics of 30 S. japonicum stool-positive sera responses to SjTPx-1 and SjPrx-4………. 44
Fig. 13 Constructs of multi-epitope recombinant proteins………58
Fig. 14 Amplification of target genes by PCR………..59
Fig. 15 Expression and purification analysis of single recombinant proteins………60
Fig. 16 Expression and purification analysis of multi-epitope recombinant proteins……..61
Fig. 17 ELISA results of single recombinant proteins and multi-epitope recombinant proteins against human serum samples……….62
iv
LIST OF TABLES
Table 1 Ct values of real-time PCR, copy number of the target gene and tissue egg counts in the liver, spleen and brain of infected mice at 8 and 18 weeks post infection……..17 Table 2 The assessment of egg burden between real-time PCR and microscopic examination of tissue sections in different organs at 8 and 18 weeks post infection……….18 Table 3 ELISA results with rSjTpx-1, rSjPrx-4 and combination of rSjPrx-4/rSjTPx-1 as the antigens……….37 Table 4 Primers used in this study for molecular cloning………...55 Table 5 List of selected peptides………56 Table 6 ELISA results of single proteins and multi-epitope recombinant proteins with human samples………..57
v
ABBREVIATIONS
Ct Threshold cycle
DNA Deoxyribonucleic acid
ELISA Enzyme-linked immunosorbent assays
GST Glutathione S-transferase
H&E Hematoxylin and Eosin
HRP Horseradish peroxidase
ICR Institute of Cancer Research
IPTG Isopropyl-thio-β-D-galactoside
MDA Mass drug administration
MFO Mixed-function oxidation
NADH Nicotinamide adenine dinucleotide
NPV Negative predictive value
O. h. nosophora Oncomelania hupensis nosophora
OD Optical density
PBS Phosphate-buffered saline
PCR Polymerase chain reaction
PPV Positive predictive value
PZQ Praziquantel
Prx Peroxiredoxin
qRT-PCR Quantitative real-time polymerase chain reaction
SD Standard deviation
SDS-PAGE Sodium dodecyl sulfate polyacrylamide gel electrophoresis
vi
SmPrx-4 Schistosoma mansoni peroxiredoxin-4
Sh Schistosoma haematobium
Sj Schistosoma japonicum
SjTPx-1 Schistosoma japonicum thioredoxin peroxidase-1
Sj7TR Schistosoma japonicum tandem repeat
SjPrx-4 Schistosoma japonicum peroxiredoxin-4
SjPCS Schistosoma japonicum phytochelatin synthase
SjLP40 Schistosoma japonicum-like protein 40
SjSAP4 Schistosoma japonicum saposin-like protein 4
SjSAP5 Schistosoma japonicum saposin-like protein 5
Sj23LHD Schistosoma japonicum 23 large hydrophilic domain
TPI Triose-phosphate isomerase
TPx Thioredoxin peroxidase
T-PBS Tween 20-phosphate-buffered saline
vii UNIT ABBREVIATIONS bp Base pair oC Degree Celsius h Hour kDa Kilodalton L Liter µg Microgram µL Microliter µM Micromolar mg Milligram mL Milliliter min Minute ng Nanogram sec Second U Unit
1
GENERAL INTRODUCTION
1. Schistosomiasis
Schistosomiasis is an acute and chronic parasitic disease caused by trematode flatworms (blood flukes) of the genus Schistosoma found in 78 endemic countries including parts of the Middle East and South America, and in sub-Saharan Africa, China and Southeast Asia (1). Globally, more than 240 million people are infected with Schistosoma spp., and an estimated 779 million people are at risk of infection (2-4).
There are six species of schistosomes known to infect human including S. mansoni, S.
japonicum, S. mekongi, S. guineensis and S. intercalatum, which cause intestinal
schistosomiasis, and S. haematobium, which causes urogenital schistosomiasis (1,5-7).
The life cycle of schistosome parasites is complex including a two-host lifecycle involving an asexual stage within a freshwater snail intermediate host and a sexual stage living in the definitive hosts (Fig. 1). The lifespan of an adult schistosome averages about 3–5 years but can be as long as 30 years (6).
Unlike other trematodes, schistosomes are dioecious. The male’s body forms a groove or gynecophoric channel, in which it holds the longer and slenderer female (Fig. 2). The adult male and female worms reside within the veins of their mammalian host, where they copulate and produce fertilized eggs. The eggs are either shed into the environment through feces or urine or are retained in host tissues where they induce inflammation and then die. The eggs released in freshwater will hatch, discharging miracidia (free-swimming ciliated larvae), which then penetrate the intermediate host snail. Within the snail, the miracidia undergo asexual replication including two generations of mother and daughter sporocysts. About 30 days after infection, the infected snail will start shedding cercariae into the water where they can remain infective for up to 3 days (8). After the cercariae penetrate the skin of the mammalian host, the resulting
2
schistosomula need about 4–6 weeks to become adults and produce eggs. These intervals (in both mammalian hosts and the snail) are termed pre-patent periods, when the infection is ongoing but the release of cercariae (from snails) or eggs (from humans) cannot be detected (Fig. 1).
2. Schistosoma japonicum
Schistosoma japonicum causes human schistosomiasis in China, the Philippines and some
parts of Indonesia. It differs from other human schistosome species in that it is zoonotic. The infection is transmitted naturally between human and more than 40 species of wild and domestic animals including cattle, water buffaloes, dogs and pigs (9). This has made the control of the disease particularly complicated. Among the main species causing human schistosomiasis, S.
japonicum is the most pathogenic due to it producing the largest number of eggs. A single
female S. japonicum can lay approximately 2200 eggs per day (10). About 30% of the eggs produced will be passed out in the feces, whereas the majority of the eggs are trapped in the liver and intestinal tissues (11) or may be carried throughout the circulatory system to other vital organs such as the brain and lungs (12).
3. Disease control
Because no vaccine is available for schistosomiasis, disease control relies solely on chemotherapy using praziquantel (PZQ). Mass drug administration (MDA) with a single dose of praziquantel is the cornerstone of national control programs and the central strategy in disease-endemic countries to control morbidity (13,14). PZQ kills the adult worms thus preventing progression of the disease to the chronic and severe phases. However, the drug does not have the capability to kill the juvenile stage of the parasite and cannot prevent possible re-infection in endemic areas (15). Although there are no reports yet about the development of S.
3
japonicum resistance to PZQ following extensive use of the drug (16), risks such as PZQ
resistance associated with a MDA program cannot be dismissed. Therefore, continuous research focused on developing new anti-schistosomal drugs and tools for pathological assessment in animal models of drug development are crucial to the control of schistosomiasis.
Moreover, MDA has successfully caused a global decrease in the prevalence of schistosomiasis. The presence of low endemic areas, however, has highlighted the inferiority of the current surveillance tools used in endemic areas. The low sensitivity of stool examination might lead to the underdiagnosis and mis-treatment of many cases and perpetuation of the parasite’s transmission. Therefore, the development of diagnostic tests with high sensitivity and specificity would be helpful in effective disease surveillance and in the assessment of the efficacy of the MDA program.
4. Schistosomiasis diagnosis
Detection of parasite eggs in stool samples such as with the Kato-Katz technique (17) has been widely used in endemic areas in the diagnosis of intestinal schistosomiasis. The quantitative Kato-Katz fecal smear (protocol in Appendix 1) is simple, cheap and useful in quantifying eggs in 40-50 mg of fecal samples (17,18) and is recommended by the World Health Organization (WHO) as the gold standard method (19). However, Kato-Katz has low sensitivity if only one stool sample from each subject is examined. Sensitivity of the test can be increased by examining multiple stool samples from the same individual. In contrast, sedimentation and concentration techniques such as the Formalin Ether Concentration Technique (20) have also been used and may increase the diagnostic yield of patients with mild infection. However, these parasitological techniques are time consuming, laborious, require a skilled microscopist and have low sensitivity in areas with a low intensity of infection.
4
More sensitive methods based on polymerase chain reaction (PCR) technology, including conventional PCR (21) and real-time PCR (22,23), have been developed for the detection of schistosome DNA in stool samples. Real-time PCR has more advantages compared to conventional PCR. It is quantitative, can detect target DNA at lower concentrations (24), and is less labor intensive as there is no need for electrophoresis to visualize products. Although PCR methods can have high sensitivity and specificity, their application for large-scale surveillance is limited. Compared to the parasitological techniques, PCR requires expensive equipment and reagents.
Immunodiagnostic antibody detection has also become important over the past decade for the diagnosis of schistosomiasis. The most commonly used techniques include enzyme-linked immunosorbent assay (ELISA) and the circumoval precipitin test, indirect hemagglutination test, and indirect immunofluorescence test (15). These methods are more sensitive and less time consuming than parasitological diagnosis (25,26) and are used when the parasitological tests are negative, particularly for patients in low prevalence areas (27). However, the immunoassays that are currently available mostly use crude antigens, either soluble egg antigen or a soluble adult worm antigen preparation derived from the parasites. These crude antigen-based serological tests have poor reproducibility, low sensitivity and specificity, and exhibit cross-reaction with antibodies against other helminths (28-30). Thus, the use of a single recombinant protein or a combination of a few recombinant proteins (chimeric proteins) as antigens in the development of a more accurate and reliable test might improve the current diagnostic formats. So far, several recombinant antigens have been identified to have good diagnostic potentials for
S. japonicum infection such as SjTPx-1 (thioredoxin peroxidase-1), Sj7TR (tandem repeat),
SjSAP4 (saposin-like protein 4) and SjSAP5 (saposin-like protein 5) (31-34).
The recent WHO road map statement emphasized the provision of regular treatment for at least 75% of children at risk by 2020 to reduce the global impact of schistosomiasis, with the
5
aim of disease elimination in all 78 endemic countries by 2025 (1). Disease elimination requires strengthened disease surveillance to ensure the success of the control programs in these areas. Accurate diagnosis and prompt treatment during the initial stages of schistosomiasis should be conducted to prevent the occurrence of severe complications. Therefore, the development of better diagnostic methods to meet the current diagnostic needs of schistosomiasis would be of great benefit.
5. Objective of the present study
The general objective of this study, therefore, is to develop tools that will aid in the pathological assessment of S. japonicum-infected organs of animal models for drug development and improvement of the sensitivity and specificity of the diagnostic test for the control of Asian zoonotic schistosomiasis. The specific objectives are (1) to use real-time PCR for the assessment of egg burden in the organs of mice experimentally infected with S.
japonicum; (2) to determine the possible role of S. japonicum peroxiredoxin-4 (SjPrx-4) in the
parasite and evaluate its immunodiagnostic potential for human schistosomiasis; and (3) to develop multi-epitope fusion antigens for the detection of human schistosomiasis.
6
Figure 1: Life cycles of S. mansoni, S. haematobium, and S. japonicum. A) Paired adult
worms (♀: female and ♂: male, male enfolding slender female). (B) Eggs (left to right, S
haematobum, S mansoni, and S japonicum). (C) Ciliated miracidium. (D) Intermediate
7
Figure 2: Scanning electron microscopic image of a pair of schistosome worms.
Source: National History Museum, UK (modified from the original figure).
8
CHAPTER 1
Use of real-time PCR for the assessment of egg burden in the organs of mice experimentally infected with Schistosoma japonicum
1.1 Introduction
Schistosomiasis is a global public health problem in 78 countries (35). Schistosomiasis japonica caused by Schistosoma japonicum is a parasitic disease that remains of public health importance in China, the Philippines and parts of Indonesia (36,37). Disease manifestations vary from being asymptomatic to severe organ involvement. It is well known that schistosome eggs, and not the adult worms, induce the morbidity seen among infected individuals (38). The number of eggs in the tissue plays an important role in determining organ damage. The schistosome eggs can be deposited in the liver and intestine of mammalian hosts where they promote the formation of inflammatory granulomas, and the main pathology of schistosomiasis. Aside from being deposited in the organs associated with the hepatic portal vein, where they can cause serious ectopic lesions (39,40), the schistosome eggs can sometimes be seen to affect the nervous system (41-43).
Understanding the disease pathology of schistosomiasis has been crucial in the management of infected individuals and in its control. Experimental infection of the parasite in mice has been widely used in investigating disease pathology (44), and the use of animal models provides a better understanding of the effects of the schistosome parasites inside the hosts (45,46). One of the pathological assessments being performed in experimental animals is the determination of the presence of eggs and their pathological effects in the affected organs through histopathological examination. However, this traditional technique has some drawbacks such as laborious protocols and lengthy time required for the microscopic examination of the samples.
9
Molecular methods have been reported to show high sensitivity and specificity for the detection of DNA of various parasites including Schistosoma from clinical samples such as stool (21,47). The high sensitivity of such methods allows the diagnosis of schistosomiasis when no eggs can be detected by microscopy during the acute phase of the infection and in cases of mild infection (48). A real-time PCR method evaluated and compared with traditional microscopic examination for the detection and quantification of S. mansoni and S. haematobium eggs in an endemic setting has shown better results (49). Moreover, real-time PCR has successfully been applied to assess the egg burden in the stool of S. japonicum-infected individuals (22,23). However, none of the studies reported has shown the use of real-time PCR in the evaluation of the egg burden in the tissues of infected hosts.
The objective in chapter 1 was to develop a more reliable and easier test that can be applied to the assessment of the parasite egg burden in the organs of an experimentally infected animal model. The performance of real-time PCR was evaluated using several vital organ samples collected from infected mice and compared with microscopic examination of tissue sections at different time points of infection.
10
1.2 Materials and methods 1.2.1 Ethical statement
Animal care and all animal experimentation were carried out according to the guidelines for the Care and Use of Laboratory Animals established by Obihiro University of Agriculture and Veterinary Medicine. This study was approved by the Committee for Animal Experimentation of Obihiro University of Agriculture and Veterinary Medicine (approval no. 28-32).
1.2.2 Animals and parasites
Thirty-one 4 weeks old male ICR mice (CLEA Inc., Tokyo, Japan) were used for experimental schistosome infection. Oncomelania hupensis nosophora snails harboring S.
japonicum cercariae were obtained from the Department of Tropical Medicine and
Parasitology, Dokkyo Medical University, Japan. Mice were infected percutaneously with 30 to 60 cercariae by directly exposing their tails with the cercariae for 30 min (50). Infected mice sacrificed included 12 mice at 8 weeks post infection and 19 mice at 18 weeks post infection. Non-infected mice serving as negative controls were also sacrificed at each time point (2 and 3 mice, respectively). Tissue samples were taken from the spleen, brain and left lateral lobe of the liver, with each sample divided into 2 parts. One part was preserved in 10% neutral buffered formalin for tissue sections, and the other part was stored at -30oC for real-time PCR.
1.2.3 Microscopic examination of tissue sections
After fixing in 10% neutral buffered formalin, the organ tissues were embedded in paraffin and stained with hematoxylin and eosin (H&E) prior to microscopic examination. Three sections were examined for each sample. To estimate the schistosome egg burden in the sample, 30 fields from the three sections were examined. The egg number in each field was counted at
11
40× magnification (Olympus, Tokyo, Japan). Average egg count for the 30 fields was calculated and was used as the value representing the egg burden in each sample.
1.2.4 Real-time PCR
Primers and TaqMan probe for the real-time PCR were designed based on the mitochondrial nicotinamide adenine dinucleotide (NADH) dehydrogenase I sequence of S. japonicum
(GenBank accession no. AF215860) (23). These were
5′-ACTGGTTATGGTTTGTTGATGTTAGGT-3′ (forward primer),
5′-AGCCACACGAACAGCACTAATC-3′ (reverse primer) and
6-FAM-5′-AGGTTCTTGGAAAAAGTAT-3′-MGBNFQ (TaqMan probe), respectively. The expected length of the amplification product was 75 bp. Template DNA was extracted from the liver, spleen and brain samples using a Qiagen tissue kit (Qiagen, Hilden, Germany). Twenty milligrams of the tissue samples were used for DNA extraction, and the DNA was eluted in 50 µL of distilled water. The 20-µL reaction mixture contained 1 µL of template DNA, 10 µL of TaqMan Universal PCR Master mix (Applied Biosystems, CA, USA), 300 nM of each primer, 250 mM of probe (Applied Biosystems) and distilled water to give the final reaction volume of 20 µL. The conditions for real-time PCR were as follows: 50°C for 2 min, 95°C for 10 min, and 50 cycles at 95°C for 15 sec and 60°C for 1 min (22). Real-time PCR was performed in MicroAmp optical 96-well reaction plates using 7300 Real-Time PCR Systems v. 2.0 (Applied Biosystems). A standard curve was prepared using serially diluted target gene cloned plasmid DNA samples. The copy number of the target gene in each sample was determined based on the standard curve and threshold cycle (Ct) value (Table 1).
12
1.2.5 Statistical analysis
All statistical analyses were performed using Prism version 5.0 (GraphPad, La Jolla, CA, USA). Spearman’s rank correlation coefficient test was used to determine the correlation between Ct values of real-time PCR and egg burden determined by microscopic examination of the tissue sections. Samples either negative for both real-time PCR and microscopic examination of the tissue sections or positive solely by real-time PCR were excluded in Spearman’s rank correlation coefficient test. A P value < 0.05 was considered significant. Ct values < 50 were considered positive, whereas values > 50 were considered negative. The negative value of real-time PCR was redefined as a Ct value of 0 for statistical analysis.
13
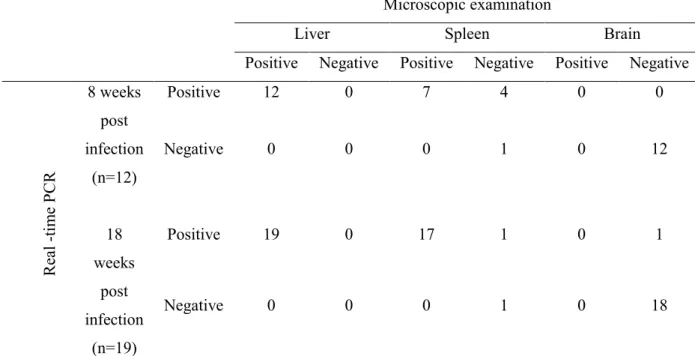
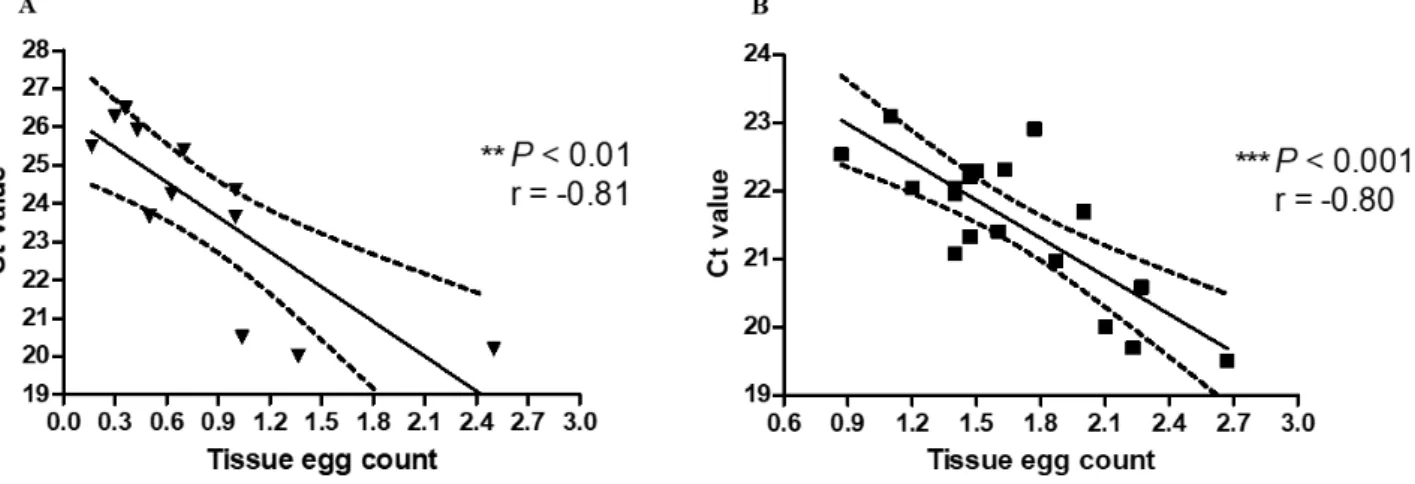
1.3 Results
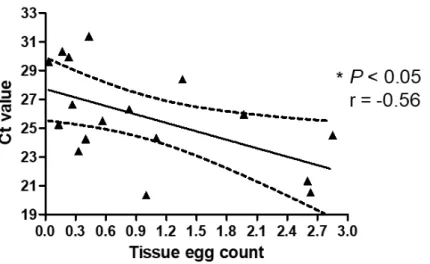
Schistosome eggs and DNA were found in all liver samples by tissue microscopy and by real-time PCR (Table 2), respectively. There was a negative correlation between the egg burden assessed by microscopy and Ct values determined by the real-time PCR. The correlation coefficients calculated between the egg burden and the Ct value for the samples at 8 and 18 weeks post infection were r = -0.81, P < 0.01 (Fig. 3A) and r = -0.80, P < 0.001 (Fig. 3B), respectively. Lower Ct values represent higher intensity of infection with increasing egg counts. Schistosome eggs were found by microscopy in 7 of 12 spleen samples collected at 8 weeks of infection (Table 2). In contrast, 11 of the 12 spleen samples collected at this time were positive by real-time PCR (Table 2). No correlation was found between the results from microscopic examination of the tissue sections and real-time PCR at 8 weeks of infection. At 18 weeks of infection, schistosome eggs were found in tissue sections of 17 of 19 spleen samples collected (Table 2). In contrast, 18 of the 19 spleen samples collected during this time were positive by real-time PCR (Table 2). There was a negative correlation between the egg burden assessed by microscopy and Ct values determined by the real-time PCR (r = -0.56, P < 0.05) (Fig. 4).
All 31 brain samples collected from the infected mice showed negative results in tissue section examination. However, one sample was positive in the real-time PCR with a Ct value of 30.60 (Table 1). The samples collected from uninfected mice were all negative in both the microscopic examination of the tissue sections and by real-time PCR.
14
1.4 Discussion
Although histological examination has been widely accepted for the assessment of egg burden in the organ samples collected from experimentally infected animals, the accuracy of this method largely depends on the number of eggs deposited in the tissues examined. Moreover, microscopic examination is labor intensive and requires skilled personnel. The results of the present study suggested that the real-time PCR examination was more reliable and easier than microscopic examination of the tissue sections for assessment of the egg burden in organs collected from experimentally infected animals (Table 2). The real-time PCR technique has been used previously to detect DNA in S. japonicum eggs in the stool samples (22,23); however, this is the first time, to my knowledge, that the method has been applied to tissue samples for the assessment of egg burden.
The liver is the principal organ affected in S. japonicum infection due to a heavy egg burden and subsequent granuloma formation (51). A heavy egg burden was observed in liver of all animals in their tissue sections at both 8 and 18 weeks of infection (Fig. 5). Real-time PCR also showed positive results for all liver samples. There was a strong correlation between the egg burden assessed by microscopy of the tissue sections and the Ct value of the real-time PCR at both 8 weeks and at the late stage of infection at 18 weeks (Fig. 3).
In contrast, a statistical correlation between microscopy and real-time PCR was found only at 18 weeks but not at 8 weeks of infection in the spleen samples (Fig. 4). Spleen incurs a moderate egg burden in the infection where approximately 0.12% of eggs laid from adult worms may be deposited (52). Real-time PCR could detect egg burden in 5 spleen samples in which eggs were not found in the tissue sections (Table 2).
In this study, real-time PCR also detected S. japonicum DNA in one brain sample of the mouse No. 6 with the highest egg burden in the liver at 18 weeks post infection (Table 1). As mentioned in a previous study, intense oviposition and continuous embolization of eggs through
15
the collaterals of the portal circulation might explain egg deposition in the brain (53,54). Furthermore, brain is known to be an organ with low egg burden in the infection where only approximately 0.03% of eggs laid from adult worms might deposit (52). Cerebral schistosomiasis has been the most understudied consequence of the disease as its neurological symptoms (55) and radiological findings are nonspecific (56). The guidelines of the European Neurological Society and the European Federation of Neurological Societies recommended that molecular techniques allow the diagnosis of infections of the nervous system from cerebrospinal fluid or biopsy material with very low burden of parasites compared with microscopic examination. (57). Therefore, the use of real-time PCR could be a means to investigate the onset and pathology of cerebral schistosomiasis in the clinical samples as well as in experimental infection.
The results of this study suggested that real-time PCR can be a useful method offering easy and accurate assessment of the intensity of infection in the organs with high egg burden such as the liver of the experimentally infected animals. This method can also be applied to organs with moderate or low egg burdens such as spleen and brain where the infection may be missed if microscopic examination alone is used. Further study is needed to prepare a standard protocol for PCR to widely apply this method in the evaluation of the intensity of infection.
16
1.5 Summary
Schistosoma japonicum, which causes zoonotic intestinal schistosomiasis, is found in
China, the Philippines and parts of Indonesia. Severe disease manifestations are basically due to the deposition of eggs in certain vital organs such as the liver, spleen and brain. Traditionally, histopathological microscopic examination of the egg burden was used to evaluate the intensity of infection in the affected organs. However, this technique is laborious, time consuming and requires trained personnel. In this chapter, the problem was addressed by establishing real-time PCR targeting the mitochondrial NADH dehydrogenase I gene to compare with microscopic examination of tissue sections in evaluating the egg burdens in different affected organs. Livers, spleens and brains of the S. japonicum-infected mice after 8 and 18 weeks of infection were harvested and examined. Statistically significant correlations were found between the egg burden evaluated by tissue section examination and the Ct values of the real-time PCR of livers with heavy egg burden at 8 (r = -0.81) and 18 (r = -0.80) weeks of infection. Furthermore, a correlation (r = -0.56) was also observed between the egg burden assessed by the microscopic examination and the Ct value of the real-time PCR of spleens with moderate egg burden after 18 weeks, but not after 8 weeks, of infection. Brains in mice with low egg burden showed no schistosome eggs in the microscopic examination, whereas one sample tested positive by real-time PCR. These results suggested that real-real-time PCR may be useful in evaluating schistosome egg burden in the organs of an experimentally infected mouse model and may provide further insights into the pathology of schistosomiasis.
17
*: Results shown as “not detected” by real time PCR was redefined as 0.0 for statistical analysis
Table 1. Ct values of real-time PCR, copy number of the target gene and tissue egg counts in the liver, spleen and brain of infected mice at 8 and 18 weeks post infection.
Mouse No.
Real-time PCR Tissue section examination
Liver Spleen Brain Liver Spleen Brain
Ct value (Mean) Copy number (Mean) Ct value (Mean) Copy number (Mean) Ct value (Mean) Copy number
(Mean) Tissue egg count (Mean)
8 wee ks p os t in fe ctio n 1 20.01 30812.37 25.50 816.37 0.00 0.00 1.37 0.60 0.00 2 24.35 1840.98 28.30 135.00 0.00 0.00 1.00 0.53 0.00 3 25.40 931.60 35.85 1.01 0.00 0.00 0.70 0.07 0.00 4 23.65 2953.19 24.63 1431.12 0.00 0.00 1.00 0.40 0.00 5 20.50 22744.84 25.44 852.99 0.00 0.00 1.04 0.00 0.00 6 26.28 521.61 0.00* 0.00 0.00 0.00 0.30 0.00 0.00 7 20.19 27150.02 23.72 2602.10 0.00 0.00 2.50 1.03 0.00 8 25.5 872.17 26.22 514.75 0.00 0.00 0.17 0.00 0.00 9 26.49 452.07 28.89 91.32 0.00 0.00 0.36 0.67 0.00 10 25.93 654.17 25.98 595.96 0.00 0.00 0.43 0.00 0.00 11 24.25 1969.93 28.02 159.87 0.00 0.00 0.63 0.27 0.00 12 23.67 2879.91 26.51 427.04 0.00 0.00 0.50 0.00 0.00 18 wee ks p os t in fec tio n 1 22.05 7763.71 20.53 26359.04 0.00 0.00 1.40 2.63 0.00 2 21.70 9865.94 31.38 23.92 0.00 0.00 2.00 0.43 0.00 3 19.70 35805.35 25.25 1196.73 0.00 0.00 2.23 0.13 0.00 4 21.41 11796.33 23.41 4011.89 0.00 0.00 1.60 0.33 0.00 5 22.54 5616.48 24.31 2213.77 0.00 0.00 0.87 1.10 0.00 6 19.51 40400.45 20.35 29627.53 30.60 35.84 2.67 1.00 0.00 7 20.59 19955.61 30.33 44.17 0.00 0.00 2.27 0.17 0.00 8 22.29 6660.61 28.40 152.17 0.00 0.00 1.50 1.36 0.00 9 20.97 14899.54 26.64 482.15 0.00 0.00 1.87 0.27 0.00 10 21.08 14606.91 24.51 1976.15 0.00 0.00 1.40 2.85 0.00 11 22.32 6511.60 26.30 607.16 0.00 0.00 1.63 0.83 0.00 12 20.01 29168.66 29.93 56.25 0.00 0.00 2.10 0.23 0.00 13 22.20 7074.01 24.24 2324.40 0.00 0.00 1.47 0.40 0.00 14 21.96 8196.81 25.52 1023.53 0.00 0.00 1.40 0.57 0.00 15 21.33 12473.06 21.34 15568.49 0.00 0.00 1.47 2.60 0.00 16 23.10 3922.27 25.96 10165.52 0.00 0.00 1.10 1.97 0.00 17 22.91 4419.76 29.62 68.87 0.00 0.00 1.77 0.03 0.00 18 22.04 7775.69 25.05 1358.41 0.00 0.00 1.20 0.00 0.00 19 22.30 6578.24 0.00 0.00 0.00 0.00 1.47 0.00 0.00
18
Table 2. The assessment of egg burden between real-time PCR and microscopic
examination of tissue sections in different organs at 8 and 18 weeks post infectiona
Microscopic examination
Liver Spleen Brain
Positive Negative Positive Negative Positive Negative
R ea l -ti m e PC R 8 weeks post infection (n=12) Positive 12 0 7 4 0 0 Negative 0 0 0 1 0 12 18 weeks post infection (n=19) Positive 19 0 17 1 0 1 Negative 0 0 0 1 0 18
19
Figure 3. Relationship between tissue egg count and Ct value in the liver. (A) at 8 (▼)
weeks post infection. (B) at 18 (
■
) weeks post infection. The Ct values and the tissue egg counts were determined and statistically evaluated as mentioned in section 2.1. The solid line is the regression line, and the dotted lines indicate 95% prediction bands.20
Figure 4. Relationship between tissue egg count and Ct value in the spleen at 18 (▲)
weeks post infection. The Ct values and the tissue egg counts were determined and
statistically evaluated as mentioned in section 2.1. The solid line is the regression line, and the dotted lines indicate 95% prediction bands.
21
A B
Figure 5. Histopathology of liver tissue sections of S. japonicum-infected mice. (A) at 8
weeks of infection. (B) at 18 weeks of infection. Paraffin-embedded liver tissues from infected mice were stained using H&E. Tissue images were captured with a microscope (Olympus) under 100× magnifications. Sites of egg deposition are shown with arrows. Scale bar = 200 µm.
22
CHAPTER 2
Analyses of the expression, immunohistochemical properties and serodiagnostic potential of Schistosoma japonicum peroxiredoxin-4
2.1 Introduction
Schistosomiasis japonica, a zoonotic disease caused by S. japonicum, still remains an economic and public health concern in China, the Philippines and some parts of Indonesia (7). A major concern making disease prevalence less controllable is the use of diagnostic methods with low efficiency in detecting infection and assessing the success of control measures (58). The standard test for intestinal schistosomiasis diagnosis in endemic areas is still the parasitological detection of eggs in stool samples, as exemplified by the Kato-Katz technique (59). Although Kato-Katz is simple and has a specificity of 100%, it is time consuming, laborious and offers unsatisfactory sensitivity mostly in endemic areas with a low prevalence of schistosomiasis (60). Therefore, many individuals with a light to moderate intensity of infection were underdiagnosed using the Kato-Katz method (61,62). Immunodiagnostic tools based on schistosome antigens have high sensitivity and are less time consuming compared to the coprological Kato-Katz test, and thus these assays have become an attractive alternative for schistosomiasis surveillance especially in those locations where the disease has been nearly eliminated. Many antigens with high sensitivity and specificity have been reported as promising biomarkers for diagnosing S. japonicum infection (32,34). However, as a single recombinant antigen used for the serodiagnosis of human schistosomiasis, the defined antigenic proteins were found to have decreased sensitivity with lower S. japonicum infection intensity level in different human cohorts (33). The low sensitivity of single recombinant antigens could possibly be improved by using multivalent cocktail antigens in leishmaniasis (63), in clonorchiasis (64), in malaria (65) and in schistosomiasis (33). To achieve this, identification of the synergistic
23
diagnostic potentials of different combinations of specific antigens that can be recognized by the host immune system is essential and will help improve testing to detect schistosome infection.
The schistosome adult worms can live in the host hepatic mesenteries for up to 20 to 30 years without treatment being administered (66,67). In this environment, the parasite must deal with reactive oxygen species mainly produced by the host immune system and those produced by the parasite itself during erythrocyte ingestion. One of the defense mechanisms which the parasite uses to protect itself is to produce an antioxidant firewall (68).
Peroxiredoxin (Prx) is a ubiquitous family of antioxidants involved in the detoxification of hydrogen peroxide (69). Based on the number and position of conserved cysteine residues, Prx has been classified into 1-Cys and 2-Cys Prx (70). The 2-Cys Prxs are also known as thioredoxin peroxidases (TPxs) because they reduce H2O2 with the use of electrons from the thioredoxin
system (71). In S. japonicum, three forms of Prxs, namely TPx-1, TPx-2 and TPx-3, have already been characterized (72). A fourth Prx, reported in S. mansoni as SmPrx-4 (73), has not yet been identified in S. japonicum.
Moreover, many studies have identified TPxs as candidate antigens for the diagnosis of various parasitic diseases such as Fasciola gigantica infection in water buffaloes (74) and
Plasmodium falciparum malaria in humans (75). Furthermore, SjTPx-1 was identified as a
potential antigen for serodiagnosis of S. japonicum infection in humans and water buffaloes in previous studies by my colleagues and me (31,76).
In this study, S. japonicum Prx-4 (SjPrx-4) gene was cloned, expressed and then its expression levels among different developmental stages, and the antioxidant activity of the recombinant SjPrx-4 (rSjPrx-4) were further analyzed. Localization of the protein was also examined and finally its diagnostic potential for human schistosomiasis was evaluated in a
24
single-antigen enzyme-linked immunosorbent assay (ELISA) and a cocktail-antigen ELISA with SjTPx-1.
25
2.2 Materials and methods
2.2.1 Ethics statement and consent to participate
Serum samples were collected from patients in the Philippines with ethical clearance sought from the University of the Philippines in Manila (clearance no: UPM REB 2011-098), which includes informed consent obtained from the individuals involved in the study.
Animal care and all animal experimentation were carried out in accordance with the recommendations in the guidelines for the Care and Use of Laboratory Animals established by Obihiro University of Agriculture and Veterinary Medicine (approval no. 29-52) and Dokkyo Medical University (approval no. 0006).
2.2.2 Animals and parasites
The Yamanashi strain of S. japonicum was maintained using O. h. nosophora snails to provide the cercariae required in this study. Miracidia were collected after the eggs were hatched in fresh water. O. h. nosophora snails were infected with 5 miracidia and were crushed six months later to collect the cercariae. Five female ICR mice aged 5 weeks old (CLEA Inc., Tokyo, Japan) were used for S. japonicum infection. The mice were infected percutaneously with 40 cercariae each and sacrificed at 8 weeks post-infection.
2.2.3 Sequence analysis
The full-length sequence of SjPrx-4 (accession no. Sjp_0053380) was retrieved from GeneDB with the S. mansoni SmPrx-4 sequence as the bait using BLASTX in NCBI (http://www.ncbi.nlm.nih.gov/). The molecular weight and isoelectric point of SjPrx-4 were calculated using the Compute pI/Mw tool (https://web.expasy.org/compute_pi/). The signal peptide was predicted using the SignalP 4.0 server (http://www.cbs.dtu.dk/services/SignalP/).
26
2.2.4 Cloning, expression and purification of rSjPrx-4
Primers were designed according to the SjPrx-4 sequence obtained from GeneDB (accession no. Sjp_0053380) with BamHI and XhoI restriction enzyme sites (underlined): forward primer (5′-GCG GAT CCA TGC TGT TAC CAA CG-3′) and reverse primer (5′-GCC TCG AGT CAT GTA GAT GAA GAG-3′), respectively. The PCR product was cloned using the TOPO PCR 2.1 vector (Invitrogen, CA, USA) according to the manufacturer’s instructions, and positive clones were selected for sequencing. The whole SjPrx-4 open reading frame was subcloned into pGEX-6p-1 vectors (Novagen, WI, USA) after digestion with the BamHI and
XhoI (Roche, Mannheim, Germany) restriction enzyme. rSjPrx-4 was subsequently expressed
in Escherichia coli strain Rosetta (DE3) (Novagen, MA, USA) in broth medium supplemented with 1 mM isopropyl-thio-β-D-galactoside (IPTG) (Wako, Osaka, Japan) and incubated at 25°C overnight. The bacterial culture was centrifuged at 10,000 × g at 4°C for 20 min, lysed by alternating freezing and thawing, and then resuspended in phosphate-buffered saline (PBS). The lysate was sonicated (Sonics & Materials Inc., CT, USA), and cellular debris was pelleted by centrifugation at 8000 × g at 4°C for 20 min. The supernatant was collected and purified using Glutathione Sepharose 4B resin (GE Healthcare BioSciences AB, Sweden) and cleaved from glutathione S-transferase (GST)-tag by PreScission Protease (GE Healthcare BioSciences AB, Sweden) according to the manufacturer’s instructions. Purified rSjPrx-4 was then dialyzed three times with PBS. The concentration of the rSjPrx-4 was determined by bicinchoninic acid (BCA) assay (Thermo Fisher Scientific Inc., MA, USA), and the protein was stored at -80oC until use.
2.2.5 Anti-rSjPrx-4 serum development
To obtain anti-rSjPrx-4 antibodies, rSjPrx-4 emulsified with the adjuvant TiterMax Gold®
(Sigma-Aldrich, MO, USA) was inoculated subcutaneously into 8-week-old female ICR mice (100 µg rSjPrx-4/200 μL inoculum/mouse). Two booster subcutaneous injections were given at
27
an interval of 2 weeks. The serum was collected and labeled as anti-rSjPrx-4 mouse serum after the third immunization. The anti-rSjPrx-4 mouse serum was used for immunohistochemical analysis as the primary antibody.
2.2.6 Evaluation of rSjPrx-4 for antioxidant activity
The antioxidant activity of rSjPrx-4 was evaluated by mixed-function oxidation (MFO) assay (77). The reaction mixture containing 40 µM FeCl3, 10 mM dithiothreitol (DTT), 20 mM
EDTA and 25 mM HEPES (pH 7.0) was pre-incubated with or without rSjPrx-4 (10 ng, 100 ng, 1 µg and 5 µg/mL) at 37oC for 1 h. Then, 500 ng of supercoiled pBluescript plasmid DNA
(Stratagene, CA, USA) was added, and the reaction mixture was further incubated at 37oC for
3 h. Nicking of the supercoiled plasmid by MFO was assessed using 0.8% agarose gel (Sigma, MO, USA) electrophoresis. The agarose gel was stained with ethidium bromide and visualized under ultraviolet light (ATTA, Tokyo, Japan). The heat-denatured rSjPrx-4 protein was included as the negative control.
2.2.7 Quantitative real-time RT-PCR (qRT-PCR) analysis
The transcriptional level of the gene coding for SjPrx-4 in each developmental stage was examined by qRT-PCR. The intramolluscan stages of S. japonicum were obtained from schistosome-infected O. h. nosophora whereas the intramammalian stages were obtained from infected mice. The total RNA samples were prepared from adult worms, eggs and sporocysts using TRIZOL reagent (Invitrogen, Thermo Fisher Scientific, CA, USA) following the manufacturer’s protocol. The cDNA library was synthesized using the PrimeScript RT reagent kit (Takara, Shiga, Japan) according to the manufacturer’s instructions. The primer pair including the forward 5′-GTTACCAACGAAACCGGCG-3′ and reverse 5′-CCCAATCCTCCTGCTTTACGA-3′ primers were used to amplify the SjPrx-4 gene with a size
28
of 273 bp. qRT-PCR was performed using an Applied Biosystems 7900HT Fast Real–time PCR System to determine the level of expression. The triose-phosphate isomerase (TPI) gene (forward primer ATGGCAGTAGAGCCGACAAC-3′ and reverse primer 5′-AACGCTTAGACCTTCTGCAA-3′) was used as an internal standard gene (72). PCR reaction mixture containing 0.5 µL of template cDNA, 10 µL of PowerUp SYBR Green Master Mix (Applied Biosystems, Thermo Fisher Scientific Baltics, UAB, Lithuania), 200 nM of each primer, and distilled water to give a final reaction volume of 20 µL was prepared. The reactions were run at 95oC for 3 mins, followed by 40 cycles of 95oC for 30 sec and 60oC for 1 min, and
a dissociation stage as per the instrument’s guidelines. Data were analyzed according to the 2–
ΔΔCt method (78,79) using the TPI gene as the internal control for each sample. The fold-changes
in transcriptional levels of the SjPrx-4 gene in the eggs and male and female adult worms were calculated relative to that of the sporocyst. Samples without the parasite DNA were included as a negative control in each PCR run.
2.2.8 Immunolocalization of SjPrx-4 in schistosomes
Immunohistochemistry was performed on 10% formalin-fixed, paraffin-embedded sections of adult worms and visceral organs obtained from the infected mice according to standard techniques (80), with some modifications. Antigen retrieval was done with microwave heat pretreatment at 97°C for 15 min in citrate buffer solution (pH 6.0). Endogenous peroxidase activity was inhibited by incubating the section on the slide glass with 0.3% hydrogen peroxide (Wako, Osaka, Japan) for 10 min at room temperature. The sections on the slide glasses were incubated with the anti-SjPrx-4 mouse serum (1:200 in PBS) overnight at 4°C in a humidified chamber. Non-specific antigen-antibody reaction was blocked with a Histofine Mouse Stain Kit (Nichirei Biosciences, Tokyo, Japan), whereas Histofine Simple Stain AP (Nichirei Biosciences, Tokyo, Japan) was used as the secondary antibody, both according to the
29
manufacturer’s instructions. Labeling was visualized with a Fast Red chromogen (Fast Red II Substrate Kit, Nichirei Biosciences, Tokyo, Japan), and the sections were counterstained with Meyer’s hematoxylin. The sections incubated with normal mouse serum instead of the anti-SjPrx-4 mouse serum served as negative controls.
2.2.9 ELISA with recombinant antigens
Three types of ELISA plates containing either rSjPrx-4, rSjTPx-1 or a combination of rSjPrx-4 and rSjTPx-1 mixed in equivalent molar amounts were prepared. Two hundred nanograms of proteins for each single antigen or combined antigen suspended in 100 µL of carbonate/bicarbonate buffer (pH 9.6) was used to coat each well of 96-well polystyrene plates
(Nunc Maxisorp, Thermo Fisher Scientific, Roskilde, Denmark) overnight at 4°C. Then, all wells
were washed three times with PBS containing 0.05% Tween 20 (T-PBS) (Wako, Osaka, Japan) to remove unbound antigens. The wells were blocked with 120 µL of 3% skim milk (Wako, Osaka, Japan) in T-PBS at room temperature for 5 min. The diagnostic performance of the antigens in ELISA was evaluated with a panel of archived serum samples including 45 non-endemic negative controls, 41 non-endemic negative controls collected from stool-egg-negative individuals in the Philippines and 30 stool-egg samples confirmed to be positive collected from the patients in the Philippines. To check the cross-reactivity of the rSjPrx-4/rSjTPx-1 combined antigen with other parasitic infections, archived serum samples from patients infected with
Paragonimus westermani (n = 10), Entamoeba histolytica (n = 9), Opisthorchis viverrini (n =
6) and Trichuris trichiura (n = 1) were used. These serum samples were diluted 1:400 with 3% skim milk in T-PBS, and 100 µL of each was delivered to the wells in triplicate. The wells were incubated at 37°C for 1 h and were subsequently washed three times using T-PBS. Then, 100 µL of horseradish peroxidase (HRP)-conjugated anti-human IgG (diluted at 1: 20,000) (Proteintech, Manchester, UK), was added as the secondary antibody to each well. The wells
30
were incubated at 37°C for 1 h and were subsequently washed three times with T-PBS. The antigen-antibody reaction was detected by adding 100 µL of HRP substrate TMB (KPL, Gaithersburg, MD, USA) to the wells, and the enzymatic reaction was allowed to proceed for 10 min. The reaction was stopped by adding 50 µL of 1 M phosphoric acid. Optical density (OD) values were read at 450 nm using an ELISA plate reader (Corona Electric, Tokyo, Japan). The cut-off values for OD were calculated from the values of 45 non-endemic negative control samples as the mean + 3 SD.
2.2.10 Statistical analysis
Data from qRT PCR (relative expression in Fig. 7) are expressed as the mean ± SD. All data were analyzed by the Student t-test, with a P value < 0.05 considered significant. Sensitivity, specificity, positive predictive value (PPV) and negative predictive value (NPV) were calculated to compare the diagnostic performance of the single and combined antigens in ELISA using MedCalc software, which is available online at https://www.medcalc.org/calc/diagnostic_test.php.
31
2.3 Results
2.3.1 Cloning and expression of SjPrx-4 recombinant protein
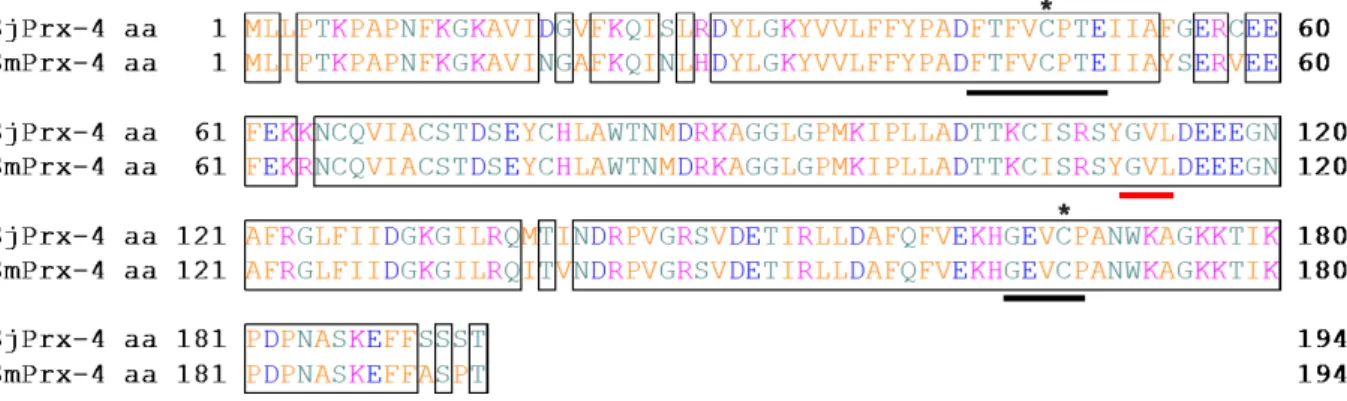
The SjPrx-4 gene was amplified by PCR from the cDNA libraries of eggs and 8-week-old adult worms (Fig. 6A). The SjPrx-4 gene was 585 bp in size, which is identical to the size recorded in GeneDB. The deduced amino acid sequence of SjPrx-4, which contains 194 amino acid residues, was 93% identical to the amino acid sequence of Prx-4 in S. mansoni (GenBank accession no. XP_018651610.1) (73). Two active site sequences (FYPADFTFVCPTE and GEVCPA) each contained the peroxidatic Cys at the N-terminus and the resolving Cys at the C-terminus, respectively, and were conserved in the SjPrx-4 amino acid sequence (Fig. 7). The SjPrx-4 cDNA was successfully expressed as a GST fusion recombinant protein in E. coli Rosetta (DE3) (Fig. 6B, lane 2). Analysis of the recombinant protein with sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) confirmed rSjPrx-4 with an expected size of 21.7 kDa after removal of the GST tag (Fig. 6B, lane 3).
2.3.2 Transcription levels of SjPrx-4 gene at different stages of the S. japonicum life cycle
The transcription levels of SjPrx-4 were examined in eggs, sporocysts and 8-week old male and female adult worms with qRT-PCR. The SjPrx-4 transcript was found in all stages examined (Fig. 8A). The highest expression level was found in the male adult, whereas the sporocyst showed the lowest transcription level (Fig. 8B).
2.3.3 Antioxidant activity of rSjPrx-4
Hydroxyl radicals produced in the MFO system allowed nicking to pBluescript supercoiled plasmid DNA, which was confirmed by changing the running behavior of the DNA in the agarose gel electrophoresis (Fig. 7, lane 4). However, the presence of >1 µg/mL rSjPrx-4 in the reaction prevented the nicking of DNA (Fig. 9, lanes 7 and 8). In addition, the presence of
heat-32
inactivated rSjPrx-4 did not prevent the nicking of DNA (Fig. 9, lanes 9 and 10). These results confirmed the antioxidant activity of rSjPrx-4.
2.3.4 Immunolocalization of SjPrx-4 in schistosomes
Immunohistochemistry assays using the anti-rSjPrx-4 mouse serum were conducted to observe the distribution of SjPrx-4 in the eggs deposited in the infected liver sections and schistosome adult worms. Sections treated with the anti-rSjPrx-4 serum showed that SjPrx-4 was mainly localized inside the egg and on the tegument and in the parenchyma of adult worms (Fig. 10A and C), as opposed to the sections incubated with normal mouse serum, which showed no positive reactions (Fig. 10B and D).
2.3.5 ELISA with recombinant rSjPrx-4 as antigen
Individually, SjPrx-4-ELISA showed 83.3% sensitivity and 87.6% specificity whereas SjTPx-1-ELISA showed 83.3% sensitivity and 97.6% specificity (Fig. 11 and Table 3). However, when rSjTPx-1 and rSjPrx-4 were combined, the sensitivity and specificity of the test improved to 90% and 92.7%, respectively (Fig. 11 and Table 3).
Human serum samples collected from the patients infected with T. trichiura and E.
histolytica showed no cross-reaction in the ELISA with the rSjPrx-4/rSjTPx-1 combined
antigen. However, 1 out of the 10 P. westermani- and 1 out of the 6 O. viverrini-infected human serum samples showed minimum cross-reactions against the combined antigen with OD values that were slightly higher than the cut-off value.
33
2.4 Discussion
Molecular cloning and characterization of 2-Cys Prxs such as 1, 2 and SjTPx-3 from S. japonicum (72,81,82) and SmTPx-1 (8SjTPx-3), SmTPx-2 and SmTPx-SjTPx-3 (84) from S.
mansoni have been reported in previous research. A fourth type, SmPrx-4, was first identified
by Protasio et al. in 2012 when they systematically improved the S. mansoni draft genome using a combination of traditional Sanger capillary sequencing, second-generation DNA sequencing from clonal parasites and reanalysis of existing genetic markers (73). Moreover, the Prx-4 homologue was also found in S. mekongi when egg proteins were analyzed using proteomic strategies (85). In the present study, a Prx-4 homologue from S. japonicum, SjPrx-4 was successfully cloned and characterized.
In this study, the SjPrx-4 full-length cDNA, which comprises a 585-bp-long open reading frame that encodes a 194 amino acid protein without a signal peptide, was obtained and it has a predicted molecular weight of 21.7 kDa (Fig. 6B). Bioinformatics analysis revealed that SjPrx-4 was most closely related to SmPrx-4 and exhibited 2 conserved cysteines, the peroxidatic Cys and the resolving Cys (Fig. 7). These cysteines of the 2-Cys Prx family are known to be essential in Cys-dependent Tpx activity (86). Moreover, the amino acid sequence of SjPrx-4 contains the GVL motif (Fig. 7), which was reported to be essential to the catalase-like activity of Prx-1 as seen in the green spotted puffer fish Tetraodon nigroviridis (87). This catalase-like activity might explain how some parasitic helminths such as Fasciola hepatica and S. mansoni are deficient in catalase (88) and have relatively low levels of glutathione peroxidase (89) yet possess highly expressed Prxs (90,91). SjPrx-4 may be an enzyme having dual functions by acting as Cys-dependent TPx and having a catalase-like activity at the same time. The catalase-like activity of SjPrx-4 should, however, be confirmed with further studies.
The antioxidant activity of Prx proteins from other helminths (84,92) showed similar activity in protecting supercoiled plasmid DNA in MFO assays. In this study, rSjPrx-4 also
34
showed antioxidant activity by protecting pBluescript supercoiled DNA from oxidative damage in the MFO system (Fig. 9). This suggests that SjPrx-4 could protect schistosome macromolecules including nucleic acids from oxidative damage by reactive oxygen species produced from the host immune system and from the internal metabolism of the worms themselves.
The results of the qRT-PCR analysis showed that the SjPrx-4 transcript was expressed in all stages of the S. japonicum parasite examined (Fig. 8A). The finding that expression of the SjPrx-4 transcript in the male worm was higher than that of the female worm (Fig. 8B) may imply the fact that the latter requires less SjPrx-4 expression as they are enclosed in the gynecophoral canal of the male worms. The more exposed male worms in turn express high levels of this enzyme to protect themselves from oxidative stress posed by the host immune system.
Immunolocalization of SjPrx-4 revealed that the protein is distributed in the space between the miracidium and the eggshell, on the tegument and in the parenchyma of adult worm (Fig. 10). The present study showed that SjPrx-4 is a tegument protein that has a better chance to be exposed to the host’s immune system during infection, and thus it may be a potential antigen for serodiagnosis in detecting the parasite infection.
The rSjPrx-4 used as a single antigen in ELISA showed comparable sensitivity of 83.3% to that of rSjTPx-1 (Fig. 11). Although both rSjPrx-4 and rSjTPx-1 were shown to be potential diagnostic antigens, their sensitivities and specificities need to be further improved to meet the current diagnostic needs of schistosomiasis. One way to improve the diagnostic capabilities of ELISA in parasitic diseases has been by mixing multiple antigens in a cocktail format. Some of the patients’ sera have high OD values with SjPrx-4 but not with SjTPx-1 and vice versa (Fig. 12). Hence, the two antigens might complement each other to have better synergistic diagnostic potential. The combination of SjTPx-1 and SjPrx-4 antigens improved the sensitivity and
35
specificity of the ELISA to 90% and 92.7%, respectively (Fig. 11). There have already been some reports on the application of fusion recombinant antigens (chimeric proteins) to the diagnosis of schistosomes (93-95) to improve the sensitivity of diagnostic methods for schistosome infection in experimental animals, goats and water buffaloes. Therefore, it appears possible to construct fusion recombinant antigens of SjTPx-1 and SjPrx-4 to enhance the accuracy and reliability of the diagnostic tools in detecting human schistosomiasis.
36
2.5 Summary
Schistosoma japonicum, living in the mesenteric vein of mammalian hosts for up to 20-30
years, is subjected to oxidative stresses from the host defense mechanism during its life cycle. To counteract this host immune attack, the parasite uses its antioxidant system for survival inside the host. Prxs, thiol-specific antioxidant proteins, play an essential role in protecting the parasite against oxidative stress by reducing hydrogen peroxide to water. Only three types of 2-Cys Prxs have been previously characterized in S. japonicum whereas a fourth Prx has been identified for S. mansoni as Prx-4. A sequence coding homologous to this gene in the S.
japonicum database was identified, characterized and expressed as recombinant SjPrx-4
(rSjPrx-4) protein. Furthermore, rSjPrx-4 was evaluated in this study for its diagnostic potential in detecting S. japonicum infection in humans. The gene found in the parasite genome contained 2 active-site cysteines with conserved sequences in the predicted amino acid sequence and showed 75% identity with that of the previously characterized Prx (TPx-1) of S. japonicum. This gene was cloned into pGEX-6P-1 vector and then transfected into E. coli Rosetta for expression of the rSjPrx-4 protein. The result of a mixed-function oxidation assay in which rSjPrx-4 prevented the nicking of DNA from hydroxyl radicals confirmed its antioxidant activity. Subsequently, immunolocalization analysis showed localization of SjPrx-4 inside the egg, on the tegument and in the parenchyma of the adult worm. ELISA results showed that rSjPrx-4 has 83.3% sensitivity and 87.8% specificity. Its diagnostic potential was further evaluated in combination with recombinant SjTPx-1 protein, yielding an improved sensitivity and specificity of 90% and 92.7%, respectively. These results suggested that SjPrx-4 may play a role as an antioxidant of S. japonicum to deal with oxidative stresses, and improvement of its diagnostic potential by coupling it with SjTPx-1 is proof that a serological test with better diagnostic performance can be developed for human schistosomiasis.
37
Table 3. ELISA results with rSjTpx-1, rSjPrx-4 and combination of rSjPrx-4/rSjTPx-1 as the antigens
Antigen Sensitivity (%) Specificity (%) PPV (%) NPV (%)
SjTPx-1 83.3 97.6 96.2 88.9 SjPrx-4 SjPrx-4/SjTPx-1 83.3 90.0 87.8 92.7 83.3 90.0 83.3 92.7
38
Figure 6. Gel electrophoresis and SDS-PAGE of SjPrx-4. (A) Agarose gel electrophoresis
image of the PCR-amplified SjPrx-4 gene from the cDNA library of eggs and adult worms. M: marker. Lane 1: eggs: Lane 2: adult worms. (B) SDS-PAGE of recombinant SjPrx-4. rSjPrx-4 was expressed in E. coli. M: marker; Lane 1: E. coli lysate before adding IPTG; Lane 2: E. coli lysate at 25 h after adding IPTG shows expression of the recombinant protein with GST-tag (SjPrx-4-GST); Lane 3: purified rSjPrx-4 without the GST-tag (SjPrx-4).
39
Figure 7. Amino acid sequence alignment of SjPrx-4 and SmPrx-4. The identical amino
acids between two sequences are boxed. Asterisk indicates the conserved cysteines at Cys47
and Cys168. Conserved amino acid sequences around two Cys conserved domains are
underlined in black (N-terminal FTFVCPTE and C-terminal GEVCPA motif). The GVL motif is underlined in red.
40
Figure 8. Different expression profiles of the SjPrx-4 gene at various developmental stages of S. japonicum. (A) Transcription of the SjPrx-4 gene was detected by RT-PCR in the
different developmental stages. Lane 1: male; Lane 2: female; Lane 3: eggs; Lane 4: sporocysts. Upper panel: Transcription of SjPrx-4 gene; Lower panel: Transcription of SjTPI gene as internal control. (B) Expression levels of the SjPrx-4 gene were determined by qRT-PCR. Data were analyzed according to 2–ΔΔCt method using the TPI gene as the internal control for each
sample. The fold changes of transcriptional level in eggs, male, and female worms were calculated relative to that of the sporocyst. Asterisk denotes statistical significance (p < 0.05).
41
Figure 9. Antioxidant activity of SjPrx-4. Lane 1: pBluescript plasmid DNA; Lane 2:
pBluescript plasmid DNA+ FeCl3; Lane 3: pBluescript plasmid DNA+DTT; Lane 4:
pBluescript plasmid DNA+ FeCl3 + DTT; Lanes 5-8: pBluescript plasmid DNA+ FeCl3 +
DTT and 10 ng, 100 ng, 1 µg, and 5 µg/mL of rSjPrx-4, respectively; Lanes 9, 10: pBluescript plasmid DNA+ FeCl3 + DTT and 1 µg, and 5 µg/mL of denatured rSjPrx-4,
42
Figure 10. Immunolocalization of SjPrx-4 in eggs and adult worms of S. japonicum. The
red arrows indicate the existence of SjPrx-4 expression. (A) The liver section from S.
japonicum-infected mice reacted with anti-SjPrx-4 mouse serum shows immunopositivity in
the space between the miracidium and the egg shell; (B) There is no reaction in the liver section from S. japonicum-infected mice reacted with normal mouse serum; (C) Immunopositive reactions of SjPrx-4 were observed on the tegument (te) and in the parenchyma (p) of S.
japonicum adult worms (♀: female and ♂: male); (D) The section from S. japonicum adult
worms reacted with normal mouse serum did not reveal any immunopositive reactions. Scale bars: 50 µm.
43
Figure 11. ELISA results with SjPrx-4, SjTPx-1 recombinant antigens and combination of rSjPrx-4/rSjTPx-1. (A) Negative serum samples from American; (B) Negative endemic
sera from the Filipinos; (C) S. japonicum Kato Katz-positive serum samples; (D) Sera positive for Paragonimus westermani (n = 10), (E) Sera positive for Entamoeba histolytica (n = 9); (F) sera positive for Opisthorchis viverrini (n = 6); and (G) sera positive for Trichuris trichiura (n = 1). Cut-off values are represented by the red dotted lines. The number of samples having OD values higher than the cut-off values are shown.
44
Figure 12. Kinetics of 30 S. japonicum stool-positive sera responses to SjTPx-1 and SjPrx-4. The red and black dotted lines correspond to the cut-off values of SjTPx-1 and
SjPrx-4, respectively. Red circles indicate the serum samples that are negative for both SjTPx-1 and SjPrx-4. Gray circles indicate the serum samples that are positive for either 1 or SjPrx-4. Black circles indicate the serum samples that are positive for both SjTPx-1 and SjPrx-4.
45
CHAPTER 3
Application of multi-epitope recombinant proteins for serodiagnosis of Asian zoonotic schistosomiasis
3.1 Introduction
Schistosomiasis is highly prevalent and remains the second most important parasitic disease in 78 countries of the world after malaria (35). The method routinely applied for diagnosis of the disease is stool-based microscopic observation for eggs of the parasite via the Kato-Katz technique. However the Kato-Katz technique is not sensitive enough to detect infections especially in locations with decreased intensity of the disease, and it may result in underestimation of the disease’s prevalence (96-98). This may hamper efforts to eliminate ongoing disease transmission and may allow re-emergence of transmission in areas where the disease was once being eliminated (99). Thus, a more sensitive means of diagnosis, such as an antibody-based test, is required (100). Several studies have reported the diagnostic potential of schistosome enzymes, schistosome excretory-secretory proteins (33,34) and tegument proteins as antigens to be used with ELISA (101). In a recent study, 10 proteins were reported to be good antigens for serodiagnosis to detect human schistosomiasis in the Philippines. Of these 10 antigens, two excretory-secretory proteins (SjSAP4, SjSAP5) and one tegument protein (Sj23LHD [large hydrophilic domain]) were reported to be good antigens for detecting the disease in human (33). Moreover, the combination of SjSAP4 and Sj23LHD or that of SjSAP5 and Sj23LHD showed better diagnostic performance compared with that of each single antigen alone. In addition, TPx-1 (SjTPx-1), tandem repeat (Sj7TR) (31), peroxiredoxin-4 (SjPrx-4) (see Chapter 2), S. japonicum-like major egg protein 40 (SjLP40) and phytochelatin synthase (SjPCS) (unpublished data) has also identified and reported as good antigen candidates with ELISA for diagnosis of the disease in humans. These antigens are highly specific and pose
46
lower cross-reactivity but are less sensitive in general in comparison with soluble egg antigen, which is commonly used in ELISA for diagnosis in humans.
If the antibody-targeting amino acid motif (B-cell epitope) within each antigen can be deduced and combined into “multi-epitope recombinant proteins”, they would display improved diagnostic performance as ELISA antigens compared with that of each single antigen (93,94). In this study, therefore “multi-epitope recombinant proteins” were constructed based on the epitope analysis of eight antigens, namely SjTPx-1, Sj7TR, SjPCS, SjLP40, SjPrx-4, SjSAP4, SjSAP5 and Sj23LHD. The diagnostic performances of three “multi-epitope recombinant proteins” as antigens in ELISA were evaluated as compared with that of each single antigen. The aim of this study was to develop a more promising ELISA antigen for detecting S.