IheJmpaneseJo"rnatofRiychonomicScience
2012,VbL 30,No.2,16S-175
OriginalArticle
Spatial
modulation
ofmultisensory
integration
ofvisual
and
tactile
motion
Evidence
from
the
redundancy
gain
paradigm
Hiroshi
UsHioDA
and
YUichi
WADA
Tbhoku
Uitiversity*
In
two
experiments,the
redundancygain
paradigm
(Miller,
1982)
was employedto
examine whetherredun-dant
visual andtactile
motion signals areintegrated
acressthese
two
modalities, andhow
the
spatial
relationship of visual andtactile
signals affects cross-modalintegration.
A
visual motion stimulus andlor atactile
motion stimuluswere
presented,
andparticipants
had
to
identify
the
motion
direction
of stimulifrom
each medality asquickly
aspossible.
It
is
well
known
that
faster
reactiontime$
areobservedfor
bimodal
stimuiithan
for
unimodal
stimuli;this
facilitation
is
termed
redundancygain
(RG),
[[he
present
study manipulatedthe
spatial relationshipbetween
the
vi-sual
and
tactile
motion stirnulito
assess reactiontime
distributions
andthe
rnagnitude efthe
RG.
Results
indicate
that
visual
andtactile
motion signals are most effectivelyintegrated
when visual andtactile
stimuli arepresented
in
the
same
spatial
location,
Keywords:cross-modal
integration,
visuo-tactile, metionperception,
redundancygain
In
our environment, adynamic
eventprovides
varioustypes
of sensorv
information,
such asvisuaL
auditerF andtactile
in-formation.
The
perceptual
systemin
the
human
brain
inte-grates
these
signalsto
create a single representatien ofthe
event.There
has
been
agreat
deal
ofinterest
in
pursuing
the
wayin
which sensory signalsfrom
different
modalitiesinte-grate.
Frorn
an evolutionarypoint
ofviewl
motionperception
is
one ef
the
mostimportant
perceptual
functions
for
survivaLOur
survivalin
acomplex environmenthas
likely
depended
upon(ameng
otherthings)
an abilityto
extractfrom
dynamic
signalsthe
motions of objects as well asliving
things
whichmay
be
potential
predators
orprey
animals.For
rnanv animals,including
humans,
visionis
1ikely
to
be
the
primary
source of sensoryinformation
ef motion signals.Although
visualmo-tion
signalstypically
tend
to
dominate
over motionsignals
in
the
other modalities(Soto-Faraco,
Kingstone,
&
Spence,
2003),
we can also extract motioninformation
frem
hearing
asound
orfeeling
atactile
stimulus.For
example, considerthe
way weperceive
an approaching vehicle;typicalIB
we respondto
the
changing
retinal size(looming)
of such an object,but
we are also sensitiveto
the
correlated rate ofincreasing
sound*
Graduate
School
ofInformation
Sciences,
Tbhoku
Uni-versityl
6-3-09
Aramaki
Aza
Aeba,
Aoba-ku,
Sendai,
yagi
980-8S79,
Japan
copY
intensity
createdby
an oncoming car,By
the
sametoken,
wecan
feel
tactile
sensations of aninsect
crawlingdown
one's armin
accordance withits
visuallocalization.
Although
a rnajerity of research on multisenseryinterac-tion
processes
has
addressedthe
integration
of static events,only recently
have
some
authors
have
soughtto
exarnine multi-sensoryintegration
of motion signals(Alais
&
Burr,
2003;
Meyer
&
Wdergeg
2001i
Meyer,
Wlierger,
Rehrbein,
&
Zetzsche,
2005;
Mitierger,
Hofoauer
&
Meyer,
2e03).
These
studies
focused
uponbenefits
that
might arisefrem
multiplepresentations
of motion signals:that
is,
they
find
that
detec-tion
performance
involving
a single eventis
facilitated
whenthe
event
conveys rnultiple rnotions signalfrom
respectivelydifferent
modalities ratherthan
information
from
only asingle modalityWhile
significant evidencefor
the
integration
ofvisua! andauditory motion signals
has
been
reported, considerablyless
is
known
aboutthe
integration
of
visual
and
tactile
metionsig-nals,
Howeve4
some evidence appearsto
reflectavisual-tactileinteraction
in
motionperception
involving
motiondirection,
For
instance,
Craig
(2006)
found
that
accuracyin
judging
the
direction
oftactile
apparent motiondeclined
whenpresented
simultaneouslywith
visualapparent
metionthat
drifted
in
adirection
opposedto
that
ofthe
tactile
motion
(versus
whenpresented
in
the
samedirection).
Using
a similarparadigm,
Bensmala,
Killebreie
andCraig
(2006)
found
that
a166
1[he
Japanese
}ournal
ofPsychonomicScience
Vbl.30,
No.
2
evant
visual
grating
drifung
in
the
samedirection
as atactile
motion
increasedi
perceived
speed ofthe
latter,
Although
these
studiesdemonstrated
that
the
relativedirection
ofvisual
mo-tion
signals can modulatepercepts
oftactile
rnotiensignals,
they
merelyillustrate
a cross-modalinteraction;
however,
they
do
notprovide
direct
evidencefor
multi-sensery
integration
per
se.1[hus,
the
existence of cross-modalintegration
for
mo-tion
signalsbetween
vision
andtouch
remainsto
be
conclu-sivelyestablished.In
the
present
study we attemptedto
provide
evidencefor
cross-modal
integration
of motion signalsbetween
visionand
touch.
Tb
addressthis
issue,
weemployed
a
frequently
usedparadigm,
known
asthe
redundancygain
paradigm
(Miller,
1982).
1[his
paradigm
permits
assessmentof
illtegration
of rnultisensory signalsfrom
different
modalities.It
requirespar-ticipants
to
respondto
two
target
signals
from
respectivelydif
ferent
modalities which arepresented
either alone Qrsimulta-neousiy
These
two
modality motions(i.e.,
bimodal
targets)
areconsidered redundant motion signals when
the
same responseis
reguiredfor
the
targets
from
each
ofthe
different
modali-ties.
It
has
been
shownthat
the
reactiontimes
(I(Ils)
to
bimod-al
targets
arefaster
than
those
to
unimodaltarget
signalsl
this
is
the
redundant-signaiseffl:ct.
It
reflects afacilitation
termed
the
redundancygain
(RG)
effect(i.e.,
redundant-signals effects).
It
has
been
assumedthat
this
redundancyfaci!ltation
is
due
eitherto
aprobabilistic
summation of unirnodalsignals
from
different
sensory modalities(a
race model)or
from
asensoTimotor
facilitation
resultingfrom
the
convergence efthe
incoming
signals, suggestedby
a
co-activation
medeLBecause
arace-modelpredicts
that
signalsfrom
two
modal-ities
independently
competeto
evoke aresponseinitiation
and
the
responseis
elicited
by
the
winner
of aracebetween
two
pfocessing
processes
in
birnodal
trials.
Since
the
likelihood
efeither of
two
sigrialsyielding
afast
reactiontime
is
higher
than
that
from
one signal alone,the
averageIUr
for
the
winner ofthe
race
in
bimodal
trials
willbe
shorterthan
the
averageIrr
ofeither
of
unimedaltrial
(Raab,
1962).
[[hus,
simpleprobability
summation couldproduce
the
RG
effect.Accord-ing
to
aco-activation rnedel,the
processing
pathways
ofboth
medalities converge somehaw ata
pa-rticular
stage whose pro-cessing eMciencyis
increased
by
multimodalinput.
This
could resultin
faster
responsesto
bimedal
stimuli,yielding
the
RG
effbctAs
Miller
(1982)
has
pointed
out, a race model makesthe
strong
predietien
that
the
RT
on'bimodaltrials
cannotbe
shorterthan
the
shortestRT
on unimodaltrials,
A
co-activa-tion
medelis
accepted whenthe
RG
is
larger
than
predicted
by
the
racemodel
assumption,MiBer
(1982)
developed
a
methodtermed
the
race-medelinequality
tesg
to
assess diffler-entpredictions
from
these
two
medels.
According
to
this
test,
aviolationof
the
predicted
distribution
ineguality
by
the
race mDdelis
consideredas
a support ofthe
co-activation model,implying
multi-sensoryintegration
of sensoryinputs,
A
num-ber
ofstudies
have
shownthat
that
a violation ofthe
race-model
inegualfty
test
resultsfrom
cross-modalinteractions,
mostly
with
visual
and auditoryinteractions
(Diederich
&
Colonius,
2004;
Mille-
1982,
1986;
Schwarz
&
Ischebeck
1994),
but
also
withvisual
andtactile
or somatosensoryones
(ForsteB
Cayina-Pratesi,
Aglioti,
&
Berlucchi,
2002;
Murray
Foxe,
Higgins,
Javitt,
&
Schroedeg
2001).
Experiment
1
was aimed atestablishingthe
RG
effeet
withvisual
andtactile
motion
signals.
First,
IUb
to
redundantmo-tion
targets
were compared withthose
to
unimodaltargets,
because
the
RG
effectprovides
asimpleindex
ofmulti-sensory
advantage.
Secend,
Miller's
race-modelinequality
tests
(1982)
were usedto
ascertain whetherRG
effectif
present,
violates
the
race modelineqLtality
If
it
does,
then
this
is
strongevi-dence
for
strong evidencefor
visual-tactile
integration
ofmo-tion
signals.Experiment
1
Method
Participants,
Eleven
undergraduateand
graduate
students(including
the
authors
HU
andYW)
at
Tbhoku
University
participated
in
the
experiment.
All
had
normal orcorrected-to-normalvision.
4pparattts
andStimuli.
A
schemadcview
ofthe
experimen-tal
apparatus
is
shownin
Figure
1,
Stimulus
presentation
anddata
collection were controlledby
anIBM-cempatible
eom-puter
running custom-written software(written
in
C)
that
in-corporated routines
for
synchronizingvibrotactile
and visual stimuli.The
tacule
stimuli
werepresented
onthe
left
forefinger
pad
ofa
participant
through
avibro-tactile stimu}ator(Optacon
II:
Model
R2B,
Tblesensory
Systems
Inc.);
the
stimulator waslo-cated
ona
table
in
front
ofthe
participants.
1[he
tactile
stimu-lus
was a simulateddynamic
line
pattern
consistingof
a hori-zontal array offive
activatedtactors.
Ihis
pattern
began
to
shiftfrom
the
center of aforefinger
pad
in
eitherthe
baclcward
or
forward
direction
atavelocity of3.0
cmfs(corresponded
to
visual angle3,O
degis).
The
intensity
ofthe
vibrotactile
stimu-H.
UsHioDA
andYL
NNiLDA:
Spatial
modulation of multisensoryintegration
ofvisual andtactile
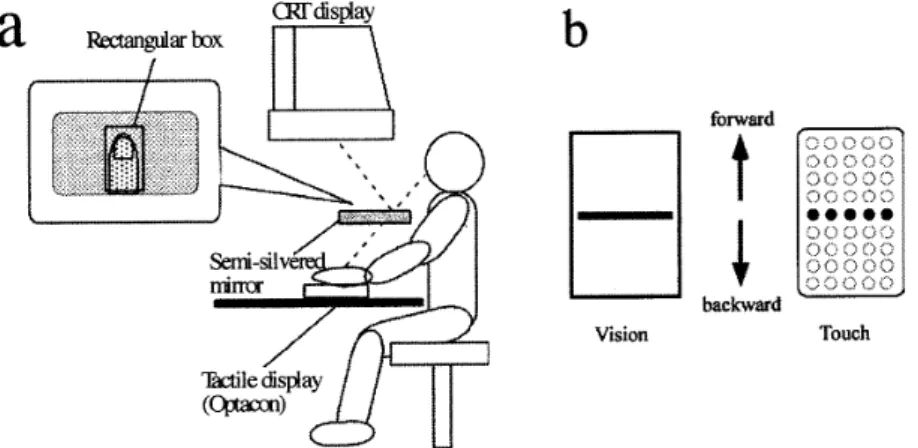
motion167
aila
anghtarboxcRrdisplay
b
forvut
t
`
Vlsienbackwaedlbuch
Figure
1.
(a)
A
diagrammatic
illustration
ofthe
experimental apparatus and stimuli,Ihe
visual
stimulus wasgenerated
on aCIrr
monitorplaced
abevethe
participant's
head,
and wasprojected
onto a semi-silvered rnirror.Participants'
hands
werepositioned
beneath
the
mirror,
The
tactile
stimulus wasgenerated
by
a
vibro-tactile
stimulator(Optacon).
In
Experiment
1,
the
tactile
stimuli werepresented
on
the
participants'
left
forefinger
pad,
In
Experiment
2,
they
werepresented
onboth
left
and right
forefinger
pad
by
two
vibro-tactile stimulatorsplaced
bilaterally
onthe
table.
(b)
A
schematic view of visual andtactile
stimulus,Visual
stimulus was a singleline
pattern
whichdrifted
within a rectarigularbox,
Thctile
stimulus was asimulated motion
line
pattern
consisted of alinear
arrayof
five
activated
tactors.
Visuai
andtactile
motion stimuli movedin
eitherthe
backward
orforward
direction,
lus
was adjustedindividuaily
at
the
beginning
ofthe
experi-ment sothat
it
couldbe
perceived
clearIMVisual
stimuli werepresented
on
a19-inch
CRT
moniter(CPD-G420,
SONYI
1024X768
resolution;100Hz
refresh
rate),which wasplaced
abovethe
participant's
head,
driven
by
a
VSG2t5
visual stimulusgenerator
CCambridge
Research
Sys-tems),
Participants
yiewedthe
refiection ofthe
visual
stimuli on a mirrorin
front
ofthem.
The
distance
between
the
partici-pant's
head
andthe
mirror
was approximately20
cm, andthe
distance
between
the
mirror andtheir
hands
was about30
cm.The
mirror was semi-silvered(the
transmission
ratio was30%),
which allowedthe
participant's
hands
to remain visibletogether
with
the
yisual stimuli,The
participant's
head
was stabilizedby
a chinrestto
ensure
that
the
visual
andtactile
stirnuli werepresented
alongthe
samehorizental
plane.
The
visualstimulus
waspresented
either
withintwo
rectangularboxes
(2.2e
×4.00
of visual angle)that
were centered8.00
to
the
left
andright
ofa
white
fixation
cross(56
cdlmZ)on
a
grey
background
(18
cdlm2),It
wasa
clynamic
pattern
consisting ofa
white
line
pattern
(S6
cdtmi)that
movedwith
censtantve-locity
from
the
center of afixed
rectangularbox,
in
either abackward
orforward
direction
withinthis
box.
The
spatialalignment of
the
dynamic
visual
line
pattern
correspondedto
the
locations
ofthe
tactile
line
pattern
throughout
both
mo-tion
patterns,
Beth
visual
aridtactile
motionsignals
moved si-rnultaneouslywith
sarne velocity(3.0e/s),
over correspondingshifts
in
space,Procedure,
Prior
to
the
experiment,participants
performed
practice
blocks
for
approximatelyle
minutes untilthey
be-came
fainiliar
withthe
stimuli andthe
task,
Each
trial
began
with
the
presentation
the
presentation
of afixation
cross andtwo
rectangularboxes,
and1000
ms
later
a
bimodal
(visual
andtactile)
or
unimodal
(visual
ortactile)
targets
waspresent-ed,
The
drifting
direction
of visual andlortactile
targets
wasdetermined
randomlyfor
eachtrial,
Participarits
werein-structed
to
judge
asquickly
aspessible
if
the
target
wasmov-ing
either
in
the
forward
orbackward
directien
usingone
of
two
correspondingknys
with righthand.
Tb
nullify anyaudi-tory
cuesgenerated
by
the
vibro-tactile stimulators, whitenoise was
presented
throughout
the
entire experiment overheadphones
(Sony
MDR-CD57e)
at7e
dB
SPL
to
rnask anysounds made
by
the
operation ofthevibro-tactile
stirnulator.
Design
and conditions.Three
different
modality conditionswere
bimodal,
visual, andtactile.
In
the
bimodal
condition,vi-sual
andtactile
stimuli werepresented
simultaneously atthe
samelocation,
and alwayshad
samedirections.
In
the
visual
ortactile
conditions, onlyvisual
or onlytactile
stimuli were re-spectivelypresented,
A
two
factor
design
crossedthe
three
modality conditions(bimodal,
visual,tactile)
withtwo
motiendirection
conditions(forward,
backward).
Each
participant
completed
360
trials
in
total,
divided
into
three
blocks
of12e
trials,
with60
trials
in
each modality conditionfor
eachdirec-168
The
Japanese
Journal
ofPsychonomicScience
VdL
30,
Ne,
2
tion.
Trial
presentation
order
was randomized withineach
block,]
Resutts
andDiscussion
Ttials
in
which aparticipant's
IUI]
exceeded
±2
standarddeviations
frorn
the
meanIUr
andtrials
for
which
apartici-pants
madeincorrect
motiondirection
responses wereexclud-ed
from
the
data.
Results
ofmeaiiIUr
data
are
shownin
Tal)Ie
1.
In
first
analy-si$,
the
mean
R[fs
for
allconditions were comparedto
confirmthe
RG
effectsfrom
redundant visual aridtactile
motionsig-nals,
1[he
Irr
data
was analyzedby
means ofatwo-way
repeat-ed rneasures analysis ofvariance(ANOVA),
with
two
within-subjects
factors
ofmodality
(bimodal
vs,visual
vs.tactile)
and rnotiondirection
(baclcbGard
vs.forward).
An
ANOVA
shewed
sigriificant main effbcts of medality condition
(F(2,2e>=
46.12,
p<.OOOI)
and motiondirection
(F(1,10)=15.98,
p<.O05),
and
asignificant
interaction
between
modalitycon-dition
andmotion
direction
(F(2,
20)=
7.37,p<.O05).
Simple
main effects analysis confirmedthat
the
effeetsof
modality condltion were significant
for
both
ofthe motion di-rection(baclcward:
F(2,
40)=[44.51,
p<,OOOI;
forward:
F(2,
40)=30,Sl,
p<.OOOI).
Multiple
comparisons(Holm-Bonfer-roni methodi
with
alphaset
at
O,05)
showedthat
the
F(Ils
to
the
bimodal
targets
were significantlyfaster
than
those
to
either visual ortactile
target
for
each motiondirection,
In
erder
to
obtaln very conservative
RG
effects,further
analysis comparedthe
RT
ofbimodal condition withthe
faster
RT
between
visualand
tactile
conditionfor
each
participant
(unimodal
condi-tion)
each motiendirection.
As
a resultthe
Rth
ofbimodal
Ial)le
1
Mean
IUTk
(rns)
(SD
in
parentheses)
for
visual,
tactiLe,
andbimodal
stimuliin
each motiondirection
in
Experiment
1.
The
meanRTS
for
unimodal conditionis
faster
Rl;
be-tween
visual andtactile
conditionfor
eachparticipant,
Motiondirection
Modalitycondition
BimodalVisualThctile
Unimodal
i
One
reviewer was concernedthat
the
randomtarget
se-quences
provide
possibie
modality switches only atthe
unimedaltrials.
It
has
been
shawnthat
the
RTb
are slew-er whenthe
target
is
preceded
by
the
target
for
adifferent
modality(Spence,
Nicholls,
&
Driveg
2001),
1[hus,
the
bi-modalfacilitation
is
possible
to
be
due
notto
the
bimodal
processing
gain,
but
to
the
modality switch costs atthe
unimodaltrials
{Gondan,
Lange,
Rosier,
&
Rode-
2004).
Howevell
the
RG
effects were stil1significant when weadded
additienal
unirnodal-enly
brocks
in
which only vi-sual ortactile
stimuli werepresented
through
eachblock,
and adoptthis
data
asthe
unimodalRts
for
analy$is(Ushioda
&
Wada,
2007).
We
confirmedthat
the
results obtained withthis
'tblockedtrials"
procedure
wereessen-tially
the
same
asthat
observedin
the
current studydemonstrating
that
the
RG
effects cannotbe
attributed
to
the
modality switch costsat
the
unimeda1trials,
Backward
325(2Z6)
342(26,7)
382(40.0)
342(26.7)
Forward
310(21.6)
340(28.4)
358(37.9>
335{27.9)
conditiens were
still
faster
than
those
of
unimedal
conditions regardless of motiondirection
[ts(9)>6.74,
ps<,Oel).
The
significance ofthe
difference
was not afuctedby
alog
trans-formation
ofthe
RIS.
Ihese
results showedthat
the
RG
effects werefound
with visual andtactile
motionstimuli,
consistentwith
previous
studies using statictargets
(Diederich
&
Coloni-us,
2004;
Forster
etaL,2002;
Milleg
1982).
Next
wedetermined
if
the
observedRG
effectsviolated
racemodel
inequality:
We
tested
race
model assumptions ofinde-pendence
ofthe
cumulativedistribution
functions
(CDFs)
ofRTb
for
different
rnodality conditions,Accordingly
CDF's
werecalculated
for
visual,tactile
and
bimodal
conditionsfor
eachmetion
direction
for
eachparticipant.
Tb
comparebimodal
and unimodal conditions accordingto
the
race modelinequal-its
this
analysiscomputed
the
sum ofvisual andtactile
CDFs
(sum
unimodalCDFs),
and
comparedthis
summedunimodal
distribution
withthe
bimodal
CDF
in
each motiondirectien
(Figure
2).
The
CDFs
ofbimodal
and unimodal cenditions wereanalyzed
by
means ef arione-tailedt-test
(p<O,05
for
matchedpairs)
at eachof
the
10
percentile
points
{5th,
15th
25th
and so on).in
the
backward
condition,the
CDF
for
bi-modal
condition
was significantlyfaster
than
those
for
uni-modal cenditionfrem
15th
percenule
to
45th
peTcentile.
Simi-larlx
the
forward
conditienalso
revealed significantlyfaster
RIls
for
bimodal
conditionthan
for
unirnodal conditionfrom
5th
percenule
to
45th
percentile.
These
resultsdemonstrate
that
RG
effectsemergedin
visualand
tactile
rnotion
signals,
and
that
the
RG
effects areconsis-tent
with
aco-activationmodel
ratherthan
with a race model,1[he
present
findings
suggestthat
visual andtactite
motien sig-nals wereintegrated
across modalities;thus,
they
are
consis-tent
withprevious
studies
that
have
shown multi-sensoryH,
UsmoDA
andYZ
MLrtDA:
Spatial
modulation of multisensoryintegration
ofvisual
and
tactile
motion169
g
i
backward
1 {ssaose4l.or`
o,tforward
iaso ev} con aN sc eeo pm 4oC
rrTCm.,c) m(crmaop
Figure
2.
Cumulative
distribution
function
(CDF)
ofIUr
for
each
motiondirection
in
Experiment
1.
0pen
circles reCDF
for
bimodal
conditien, andfi11ed
squares referto
the
sum oftwo
CDFs
for
visual
and
tactile
conditions.ferto
the
(Soto-Faraco
et
aL.
2003).
The
primary
purpose
ofthis
analysis wasto
confirmthat
RG
effects obtain with visual andtactile
metion signals, How-eve4the
analysis of meanIUls
also showed asignificant main effectof
motion
direction
and a significantinteraction
be-tween
modality and motiondirection.
Post
hoc
analysisre-vealed
that
the
RTs
to
forward
direction
were significantlyfaster
than
those
to
backward
direction
in
bimodal
conditfon(F(1,
30)=le.Ol,
p<.O05)
andtactile
condition(F(l,30)=
25.20,
p<,OOOI),
whereas
nodirectional
difference
appearedin
the
visual
condition(F(1,
30)
==O.17,
p=.68).
The
difference
observed
in
tactile
condition couldbe
accountedfor
by
the
fo11owing
explanation,The
tactile
motion stimulus usedin
thi$
study was a single
line
pattern
which emerged atthe
center ofa
forefinger
pad,
andthen
began
to
movein
either
a
backward
or
forward
direction.
This
meansthat
the
backward
and for-wardtactile
motionpattern
first
stimulated eitherthe
bottom
or
top
sideof
a
forefinger
pad,
respectivelyPrevious
neuro-physiological
studieshave
shownthat
tactile
sensors
sensitive
to
transient
stimulation are mainlydistributed
on
the
top
ef
a
forefinger
pad
rather
than
onthe
bottom
(e.g.,
Thlbot,
Darian-Smith,
Kornhuber,
&
Mountcastle,
1968;
Vallbo
&
Johansson,
l978).
Thus,
this
type
of sensorson
the
top
of aforefinger
pad
presumably
produced
the
faster
detection
speedsfor
the
for-ward rnotion stimuli,
This
raises aquestion
concerning adifference
found
in
the
bimodal
condition.In
this
conditionthe
sizeof
the
RG
effectdiffered
withdirection;
RG
for
the
forward
direction
(25
ms)was
greater
than
that
for
the
backward
direction
(17ms),
However,
the
I(fs
for
the
unimodal conditionsthe
RTb
(faster
Rts
from
each ofthe
twe
single modality conditions)did
notdiffer
due
to
direction,
A
possibie
explanationis
that
the
RT
difference
between
visual
andtactile
conditionsis
relateclto
80
60
Ee
4oUer
20
o
O
20
40
60
80
absolute
(RTtV]
-
RTITI)
{rnsec)
Figure
3,
The
magnitude of redundancygain
(RG)
for
each
participant
asafunction
ofRT
difference
between
visual and
tactile
conditionsin
Experiment
1.
ward and upward
triangles
referto
the
backward
andforward
motiondirection,
respectively
the
magnitude ofRG
effect.Here
we calculatedthe
RG
valuesfor
each motiondirection
for
eachparticipant,
then
preduced
acorrelation coeficient
between
the
Irr
difference
ofunimod-al conditions andthe
obtainedRG
(Figure
3),
As
a result,a
significantnegativecorrelatienwasfound(r='O.48,p<,05),
indicating
that
smaller
differences
in
the
IU]
difference
ofuni-modal conditions accompany correspondingly
greater
RGs.
If
we regardthe
smal1RT
differences
between
visual and tactileconditions as support
for
the
notionthat
the
time
course of senseryprocessing
is
similar
for
both
stirnuli,these
results seernedto
be
in
line
withthe
previous
studies with staticstim-uli
showingthat
the
largest
RG
was observed whentwo
ColQni-170
TheJapanese
Journal
of
Psychonemic
Science
Vbl.30,
No.
2
us,
2004;
MfileL
1986).
Experiment
2
The
purpose
ofExperirnent
2
wasto
exarnine whetherthe
RG
effectsshownin
Experiment
1
depend
onthe
spatial
sepa-ration of visual andtactile
motiontargets.
Multisensory
inte-gration
appearsto
occur most stTongly when stimulifrom
dif
ferent
sensory modalitiesare
presented
atthe
same spatialposition
at
approxirnatelythe
sarne
time,
implying
the
impor-tance
ef spatiotemporal coincidencefor
the
processing
ofsi-multaneous
stimuli
from
different
modalities(e,g,,
Stein
&
Meredith,
1993).
For
audio-visual motionintegration,
it
has
been
reportedthat
integration
of multisensory motionsignals
appearedto
be
most effective whenthe
stimulipresented
to
different
modalities aredelivered
from
the
same or adjacent spatiallocation
(Meyer
&
Wiierger,
2001;
Meyer
et
al..
2005;
Soto-Faraco,
lyons,
Gazzaniga,
Spence,
&
Kingstone,
2002).
For
example,Meyer
etal.
(2005)
found
that
multiple visualand auditory motion signals
decreased
discrimination
thresh-oldfor
motiondirection.
FurtheTmore,
they
found
titat
the
lowest
threshold
wasobserved
when visualand
auditorymo-tion
signals were spatially co-localized, as consistentwith
find-ing
involving
static
stimuli(Frassinetti,
Bolegnini,
&
Ladavas,
2e02),
In
addition,Soto-Faraco
etal,
(2oo2)
examinedthe
vi-sual-auditory
interaction
ofmotion signals and alsofound
aninfluence
ofthe
spatial separationof
simultaneous
compo-nents on
the
perceptual
interaction
of auditory andvisual
sig-nals,
By
contrast,
there
has
been
relativelylittle
research onthe
effects ofspatial
modulationfor
multisensory motionintegra-tion
between
vision
andtouch.
Data
relevantto
this
issue
haxre
been
reportedby
Craig
(2e06),
who used aparadigm
in
which stibjectsjudged
the
directien
oftactile
apparent motion while simultaneouslyviewing
task-irrelevant
visuai apparentmo-tion.
He
found
adecline
in
performance
whenthe
tactile
andvisual apparent motions moved
in
opposite
directions.
How-evegCraig
also Teportedfinding
noinfluence
ofthe
spatialdistance
between
the
visual andtactile
stimuli onthe
sire ofthis
effect.1[he
latter
finding
seemsto
be
inconsistent
with
the
notionthat
the
spatial separation amongsensory
modalitiesis
crucial
factor
for
integrating
motion signals,However,
as
nDt-ed
by
the
author
himselC
Craig's・
null
findings
could sufferfrem
severalpossible
problems
with
the
interpretation
ofthe
results,perhaps
the
rnostimportant
ofwhichis
the
possibility
that
the
visual
andtactile
stimuLi might net suMciently apartfrom
each otherfor
revealing
the
effectin
the
condition(the
"far"condition)
in
which
the
two
stimuli werepresented
in
different
spatialpositions.
This
leaves
some
uncertainty
asto
the
reality
of nul1 effect ofthe
spatial separationfor
visuo-tacule
motionperception.
[Eherefore
wethought
it
worthwhileto
determine
whetherthe
cross-modal motionintegration
we$ modulatedby
the
spatial relationshipbetween
the
tactile
and visual stimuli. Ac-cordinglyin
Experiment
2
weinvestigated
RG
effects as afunction
ofvariationsin
spatiallocations
ofvisualand
tactile
stimuli
in
order
to
assess
integration
efecbs usingthe
race-model
ineguality
test.
If
it
is
the
casethat
the
spatialcoinci-dence
plays
arole,then
we shouldfind
1arger
RG
efflectsalongwith violations of
the
ineguality
test
for
multisensoryintegra-tion
when
the
tactile
and visual stimuli appear atthe
same spatiallocations
than
whenthey
appearat
spatiallydistinct
lo-cations,
1[hus
in
Experiment
2,
the
visual
arid
tactile
motion stimuli werepresented
either
at
same
location
orat
different
locitions.
Mbthod
Participants.
:thirteen
undergraduate andgraduate
studentsat
lbhoku
University
participated
in
this
experiment.Except
for
the
authorsHU
andyw,
al1
participants
were naiveto
the
experimentalmariipulations.apparatus
andStimuli,
Z[his
experiment
usedtwo
vibro-tac-tile
stimulators which wereplaced
13,O
cm apart(horizontal-ly)
onthe
table
in
front
ofthe
participants,
One
ofthe
two
vi-bro-tactile
stirnulaters wasfor
participants'
left
forefinger
pad,
andthe
otherfor
the
right
forefinger
pad,
In
eachtrial,
atac-ule
stimulus waspresented
at eitherleft
or
rightforefinger
pad,
SimilarlF
visual stimuli werepresented
at-eitherleft
or rightside
of
a
central-fixation
cross withtwo
fixation
boxes
positioned
6.5
degrees
left
and right ofthe
fixation
cross(cor-respond
to
the
locations
oftactile
stimuli).Procedure.
Each
trial
began
withthe
presentation
of afixa-tien
cross andtwo
fixation
boxes
and10oo
ms1ater
targets
were
presented.
Participants
judged
motion
direction
usingone of
two
correspQndingfoot
pedal
with
right
foot
Design
and conditions.In
this
experiment,the
cgnditionfor
bimodal
condition
was
divided
into
two
different
bi-modal
conditions
by
the
spatial separation:matehed
bimedal
(m-bi-modal) and unmatchedbimodal
(u-bimodal),
In
the
m-bi-moda1 condition,
visual
andtactile
stimu}i werepresented
at
same}ecation
in
eitherleft
or right side, whereasin
the
u-bi-H,
UsHioDA
andYL
MeLDA:
Spatial
modulationofmultisensoryintegrationofvisual andtactile
motion171
medFl condition,
the
stimuli werepresented
atdifferent
loca-tiens
separatelvIn
the
visual
ortactile
condition,only
visual
er
only
tactile
metien stimuli werepresented
at eitherleft
or
rightside,Tb
equalizethe
occurrence rate ofbimodal stimuliwith
that
in
Experiment
1
(a
third
oftotal
trials
wasfor
bimedal
cendi-tion),
the
numbers of m-bimoda! and u-bimodaltrials
werehalf
ofthe
visual
andtactile
trials
(SO
trials
for
m-bimodal andu-bimodal
and
1OO
trials
for
visual andtactile
×2
motion
di-rection conditions ×2
locations),
[lhus,
eachparticipant
com-pleted
l200
trials
in
total,
divided
into
ten
blocks
of120
trials.
ResultsandDiscussion
Thble
2
showsthe
meanRIg
for
al1
conditions.[[he
data
were analyzed
by
means of a two-way repeated measuresANOVA
withfacters
of modality conditions(m-bimodal
vs.u-bimodal vs. unimodal) and motion
direction
(backward
vs,
forward).
1[here
wasa
significant
rnain effect of modalitycon-ditien
(F(2,
24)
==72.52,p<.OeOl)
and a significantinterac-tion
between
modality condition and motiondirection
(F(2,
24)=4,32,
p<.05),
whereasthere
was no significant main eefect
of motiondirection
{F(1,
12}==O.14,
p=O,71).
Simple
main effects analysis confirmedthat
the
effects of rnodality condition weresignificant
for
both
motiondirectiens
(back-ward:F(2,48)=24,10,
p<.OOel;
fbrward:
F(2,40)==40.87,
Thble2
Mean
IUls
(ms)
(SD
in
parentheses)
for
each experiment conditionin
Experiment
2.
The
meanRTs
for
un-imodal
conditionis
faster
Irr
between
visual
andtactile
conditionfor
eachparticipant
Motiondirection
Modalitycondition
Bimodal
(matched)
Bimodal
(unmatched)
Visual
factile
Unimodal
Backward
Forward
352
(29.0)
346
(24,6)
358
(3e,4)
355
(24.8)
370
(29.8)
375
(22.3)411
(36.8)
393
(32.2)370
(29.8)
373
(23.7)
teas
2.
oei"g,.o
matehed(backward)
7
/e'
'"
fZ lr1ee-sies'S
eAgo.t mpas e-. sus5eRTtmDec) mstched(forward)!
i
f
o.ssSo,SOAsn.z
unmatchedibackward)
bO・ei e.eE・g ,,,:t O.2 30PaseRVCmeeptimmatched(forward)
4scFigure
4,
CDFs
of spatially matched and unmatched conditionsfor
each metiondirection
in
Experiment
2.
0pen
circ!esrefer
to
the
CDF
for
bimodal
matched
or unmatched condition, andfilled
squares referto
the
sum oftwo
CDFs
for
visual andtactileconditions.172
1[he
Japanese
Journal
ofPsychonomlcScience
Vbl,
30,
No.
2
p<.OOOI).
Multiple
comparisons showedthat
the
IUb
for
the
m-bimodal and u-bimodal conditions werefaster
than
those
for
unimodal conditionfor
both
motiondirections;
moreim-portantly
the
M]
of
the
u-bimodal condition was significantlyslower
than
those
ofthe
m-bimodal
condition.Next,
CDFs
for
the
m-bimodaland
u-bimodalconditions
were compared with
the
sum ofthe
visual andtactile
condi-tions
(Figure
4)
in
orderto
test
whetherthe
observedRG
ef-fects
violate
the
race modelinequality
As
in
Experiment
1,
the
CDFs
ofbimodal
and unimodal conditions were analyzedby
means of an one-tailed
t-test
ip<e.05
for
rnatchedpairs)
at each ofthe
10
percentile
points
(5th,
15th,
25th
and so on).Results
showedthat
violations
of
the
race
model
occurred
in
the
m-bimodal condition; significantdifferences
between
bi-medal and unimodal conditions occurredfoT
percentiles
be-tween
O
and25
for
the
backward
motion condition andbe-tween
O
andthe
45th
percentile
in
the
forward
motion condition.HoweveG
in
the
u-bimodal condition, afar
smallerviolation
wasfound,
and
this
ernerged enly at15th
percentile
point
in
the
forward
condition.
Experiment
2
replicatedthe
basic
results ofExperiment
1
for
the
modality conditionin
which
visual andtactile
motion stimuli occurred atthe
same spatiallocation.
On
the
other
hand,
when visual andtactile
metion stimuli werepositioned
separatelF
the
RG
effectsdecreased
appreciablyand
the
viola-tion
of race modelinequality
almost
vanished,thus
implying
that
the
spatial
separationis
acriticalfactor
for
integrating
yi-sual
andtactile
motion signals.1[his
finding
is
in
line
with
a similarstudy
which
combined
visual
motion signals withau-ditory
ones(Meyer
etal.,
2005);
in
the
iatter
research,the
spa-tially
unrnatched visual and auditorymetion
signals
produced
a small reductionin
the
detection
threshold
in
the
extent
of
probability
summation.Spence,
Pavani,
andDriver
(2004)
also
showed
that
the
effectsof
a
visual
distractor
on
detection
per-formance
in
atactile
localization
task
were considerably di-minished when avisual
distractor
waspresented
atalocation
of
the
tactile
stimulationbut
onthe
oppositehand.
On
the
etherhand,
as
mentionedear]ier,
Craig
(20e6)
has
reported contradictory results,in
which visual motioniniiormation
ro-bustly
influenced
tactile
rnotionperception
even whentheir
locatiens
were s?atiall7 unmatched.Several
noteworthy methodologtcaldifferences
distingujsh
the
present
studyfrom
Craig's
andthey
renderdiMcult
direct
comparisons across
the
two
studies.For
instance,
in
Craig's
experiment
the
main
dependent
variable
irrvorved
cross-modal
interference
effects
caused
by
task-irrelevant
visual motion.In
addition,
the
spatial separation(13cm)
in
the
unmatchedconditiens
(u-bimodal)
ofthepresent
studydiffers
from
those
in
the
Far
condition ofCraig's
studyIn
the
current study visu-al andtactile
motion stimuli werepresented
in
the
opposite
hemispaces
whereasin
the
unmatched conditiQnin
Craig's
studM
tacdle
stimuli
and
visual
stimuli
appearedin
the
samehemisphere.
That
is
tactile
stimuli werepresented
only atthe
left
forefinger
pad,
and spatiany unmatchedvisual
stimuli
were
located
along
the
vertical meridian,Thus,
althoughposi-tions
of
the
visual
andtactile
stimuliin
Craig's
design
werespaced
29.5
cm apart,they
were yerticallyaligried
meaning
that
the
two
stimuli appearedin
the
samehemispace.
In
this
regard, we can assume
that
the
stimulus settingin
Craig's
study mightgenerate
multisensory motion sensationthat
orig-inates
from
the
same event.[[he
verticalalignment
oftwo
stimuli
may
have
provided
suMcient cuesto
encourageper-ceptual
grouping,
and
consequentlsthe
two
motion signalsmay
have
been
treated
as asingle motion stream,As
highlight-ed above, co-occurrence oftemporal
andspatial
stimuli
is
censideredto
animportant
factor
for
multisensoryintegration
<Stein
&
Meredith,
1993),
In
this
regard we, can assumethat
parsing
inputs
into
one
perceptual
group
couldfacilitate
mul-tisensory
integration.
This
could
be
a
possible
explanationfor
Craigds
(2006)
failure
te
observe ariy effects of spatialsepara-tion
in
his
study.CIearlM
atthis
point,
this
explanationis
spec-ulattve.It
also
should
be
noted
that
in
Forster
etal,(2002)
the
Rfs
for
bimodai
stimulipresented
atdifferent
hemi-spheres
did
not sigriificantlydiffer
from
those
presented
atsarnehemi-sphere
(see
also
Strybel
&
Vatakis,
2004).
As
the
present
studyhas
not systematically manipulatedthe
spatialdistarice
oralignment
between
the
visual
and
the
tactile
stimulus
location,
we cannotdirectly
addressthe
question
ofhow
the
spatialdis-tance
or alignment ofbirnodal
stimuli affectthe
degree
ofRG
effectsquantitativelM
[thus,
future
researchshould
further
in-vestigatethe
role ofspatialfhctors
onmultisensory
motion
in-tegration
between
vision
andtouch.
General
Discussion
The
present
study usedthe
redundancygain
paradigm
to
ascertain whether redundant yisual and
tactile
motion signalsproduce
greater
performance
on motiondirection
discrimina-tion
than
singlevisual
ortacule
signal, and whether perfor-mancedepends
uponthe
spatial separation ofvisual
andH,
UsHioDA
andYZ
INOxDA:
Spatial
modulatien ofmultisensoryintegration
ofvisual andtactile
motion173
In
Experiment
1,
it
was shownthat
the
detection
speedfor
the
rnotiondirection
of visual andtactile
targets
was signifi-cantlyfaster
than
the
response
speed
for
visual
ortactile
uni-modaltargets,
Le.,
anRG
effect.An
impertant
finding
in
this
experimentwas
that
the
RG
effect was well explainedby
a co-activation model ratherthan
a racemodel.
This
implies
that
the
ebservedI(Ir
facilitation
in
certainbimodal
conditions maybe
explainedby
aneural summatienprocess
rather
than
by
aprobability
advantage,
Although
some empiricaldata
have
shownpossible
cross-modallinks
in
the
processing
ofvi-sual and
tactile
motioninformation
(Bensmaia
etal.,
2e06;
Craig,
2e06),
to
our
knowledge,
the
present
studyis
the
first
to
show asignificant
RG
effect andto
demonstrate
the
violation of race modelinequality
with
visual andtactile
motion
signals,
As
such
Experiment
1
is
consistentwith
the
hypothesis
that
vi-sual and
tactile
motion signals areintegrated
across sensorymodalities.
In
Experiment
2,
spatial separatien ofvisual
andtactile
stimuli significantly reduced
the
RG
effects relativeto
tho$e
observed
for
spatially coincidingstimulL
Furthermore,
where-as
with
the
spatially coinciding stimulithe
RG
effects wereconsistent with aco-activation model,
the
RG
effectsfound
for
spatialiy
separated conditions were morein
line
with
a race modeLThat
is,
when visual andtactile
signals arepositioned
separatelythey
appearto
be
processed
by
the
independent
de-tection
systems,
It
shouidbe
noted asignificant
violation ofthe
racemodel
inequality
was ob$ervedin
the
unrnatched conditionin
Experiment
2,
suggestinglimitations
on infer-ences oftotally
independent
systems.In
this
case,however,
the
violation
wasquite
modest and restrictedto
a
very
narrow range ofpercentile
points
efthe
RT
data,
Nevertheless,
it
sug-gests
that
we cannotfirmly
concludethat
visual andtactile
motion signals are
not
integrated
even when sources ofthese
signals are spatially separated.Based
onthe
results ofthis
studFit
is
not clear whythe
latter
phenomenon
occurred.
From
the
fact
that
we can see an obvieusdifference
in
the
re-sults ofthe
race modelinequality
test
between
the
matched and unmatched cenditions, webelieve
that
the
results of ouri
second experiment reflect
the
spatialmodulation
on eMciency of multisensoryintegration
between
vision
andtouch.
The
possibility
remains,however,
that
the
reduction ofde-tection
performance
due
to
spatial separationin
Experiment
2
resulted
frorn
possible
factors
associated withspatial
attention(Spence
&
Driver,
1997).
When
visual
andtactile
stimuli werepresented
atdifferent
locations,
divided
spatial attention couldproduce
slowerur
for
detecting
the
two
signalsin
different
locations
than
RT
for
detecting
signals occurring atthe
samelocation,
If
this
werethe
case,then
one might assumethat
the
spatial modulatien of
RG
effects observedin
Experiment
2
is
the
result of cross-modal consequences of spatial attention.Howeve4
this
potential
argumentis
weakenedin
the
light
of
several recent
findings
suggestingthat
multisensoryintegra-tion
can occurprior
to
orindependent
of attentional selection(Bertelson,
Vlreemen,
De
Gelde4
&
Driver,
2000;
Driver,
1996;
Soto-Faraco,
Ronald,
&
Spence,
2004;
Vteemen,
Bertelson,
&
de
Gelde4
2001).
Howeveg
asnotedby
McDonald,
Ileder-Slilejarvi,
and
Ward
(2001),
it
is
dithcult
to
unambiguously separatethe
processes
of multisensoryintegration
and spatial attentionin
someex-perimental
paradigms.
This
is
because
the
two
efiects cance-exist and
produce
additivefacilitation
of responsesto
targets.
In
addition,because
attention
is
consideredto
play
a criticalrole
in
the
integration
efstimulus
features
within asingiesen-sory
medality
(e.g,,
[freismari
&
Gelade,
1980)
it
is
not unrea-senableto
assume considerable overlap obtainsbetween
the
processes
ofmultisensoryintegration
andthat
for
cross-modalspatial attention.
In
this
regard,Oruc,
Sinnett,
Bischoff]
Soto-Faraco,
Lock,
andKingstone
(2008)
reported
that
attentiondid
modulatethe
degree
to
whichthe
motion signals arecom-bined
across modalities atleast
in
some ca$e$.It
remains un-clea4however,
whetherintegration
process
occurspreatten-tively
for
al1
kinds
ef multisensory events without employingattention,
or whether attended objects areintegrated
different-ly
from
those
that
are not attended,It
is
also unclear whether attention affectsthe
eMciencyof
integration
of stimulu$fea-tures
acressdifferent
sensory modalities.As
there
were no di-rect manipulations of attentionin
the
present
studMfurther
workis
neededto
revealthe
possible
role of spatialattention
onthe
effectivenessof
integrating
two
or more sensoryinputs.
Overall,
the
present
studydemonstrated
that
redundantvi-sual
andtactile
metien stimulifacilitated
detection
perfor-mance, and