近畿大学学術情報リポジトリ
68
0
0
全文
(2) Transcriptional regulation of DNA repair gene expressions by bacterial signal transduction system Hiroshi Ogasawara. March, 2005 Graduate School, Kinld University Division of Agricultural Science Major: Applied Bioscience (Adviser: Prof. Ryutaro Utsumi). Submitted to the Graduate School, Kinki University, to fulfill the requirement for the Doctorate Degree..
(3) ABBREVIATIONS. ATP. adenosine 5' -triphosphate. Apf. ampicillin resistant. Bp. base pair. BPB. bromophenol blue. BSA. bovine serum albumin. Cmf. chloramphenicol resistant. cpm. counts per minute. D. aspartic acid. Da. dalton. DNA. deoxyribonucleic acid. DNase. deoxyribonuclease. DTT. dithiothreitol. EDTA. ethy lenediaminetetraacetate. HEPES. N -2-Hydroxyethy lpiperazine-N' -2-ethanesulfonic acid. His (H). histidine. IPTG. isopropyl-I-thio-D-galactopyranoside. Kmf. kanamycin resistant. LB. Luria-Bertani. OD. optical density. ORF. open reading frame. PAGE. polyacrylamide gel electrophoresis. PCR. polymerase chain reaction. RNA. ribonucleic acid.
(4) RNAP. RNA polymerase. RR. response regulator. SDS. sodium dodecyl sulfate tetracycline resistant. TCS. two-component system. Tris. 2-amino-2-hydroxymethyl-I,3-propanediol. UTP. uridine 5' -triphosphate. UV. ultraviolet.
(5) CONTENTS CHAPTER I. Introduction. 1. CHAPTER II. Negative regulation of DNA repair gene. 6. (ung) expression by CpxR/CpxA. two-component system in Escherichia coli K-12 and induction of mutations by increased expression of CpxR. CHAPTER III. Negative regulation of DNA repair gene. 28. (uvrA) expression by ArcA/ArcB. two-component system in Escherichia coli. CHAPTER IV. Summary. 44. ACKNOWLEDGEMENTS. 46. REFFERENCES. 48. PUBLICATIONS. 63.
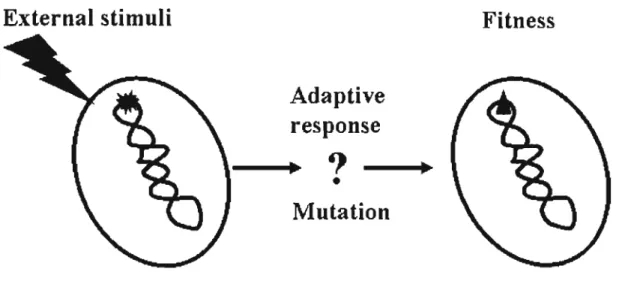
(6) CHAPTER I. Introduction. Evolution is defined as the result of environmental stress that selects the fittest. The yet unresolved and fascinating question concerns the origin of the variants from which the fittest survive. Dogma has it that these variants arise from pre-existing random mutations that are not directed by the different conditions of stress that select them. In other words, there would be no specific biochemical mechanisms initiated by each stressor that can increase mutation rates in those genes related to that stress (Wright et al., 2004). However, specific, stress-induced mutagenesis (Fig. 1) would offer an enormous advantage for evolution and would be selected. Therefore, if stress-induced mutagenesis exists, it should be present today in organisms coping with adverse conditions in their environment. Increased mutation rates would be directed to specific genes that must mutate to alleviate the stress, while avoiding random genome-wide damage. Recent understanding of stress-induced mutagenesis or adaptive mutagenesis in microbes indicates that mutation rates can be elevated in response to stress, producing beneficial and other mutations (Hersh et al., 2004). In Escherichia coli, two separate mechanisms of stress-induced genetic change occur that revert a lac frameshift allele allowing growth on lactose medium. First, compensatory frameshift (point) mutations occur by a mechanism that includes DNA double-strand breaks and their error-prone repair. Point mutation requires induction of the RpoS-dependent general stress response, and the SOS DNA damage response leading to upregulaiton of the error-prone DNA polymerase DinB (Pol IV), and occurs during a transient limitation of post-replicative. 1.
(7) mismath repaIr activity. A second mechanism, adaptive amplification, entails amplification of the leaky lac allele to 20-50 tandem repeats. These provide sufficient j3-galactosidase activity for growth, thereby apparently deflecting cells from the point mutation pathway. Similar processes are being found in other bacterial systems and yeast. Stress-induced mutagenesis may underlie much of microbial evolution, pathogenesis and antibiotic resistance, and also cancer formation, progression and drug resistance. (Bjedov et al., 2003, Wright et al., 2004). Two-component signal transduction systems have a key role in mediating the response of bacteria to environmental stimuli. Normally, receptor-mediated detection of a stimulus at the cell surface leads to autophosphorylation of a sensor kinase component, which then phosphorylates the effector protein component (i.e., the response regulator), enabling the effector to bind to operator/promoter sequences of target genes and either increase or repress transcription (Hoch & Silhavy, 1995). Using DNA microarrays (http://ecoli.aist-nara.ac.jp/xp_analysis/2_components/), 23 DNA repair and replication genes regulated by Two-component systems containing CpxR/cpxA and ArcAlArcB were found (CHAPTER III, Table1) (Oshima et ai, 2002). In this thesis, the author focused on CpxR/CpxA (Chapter II) and ArcAlArcB(Chapter III) two-component systems to clarify the transcriptional regulation of DNA repair and replication genes expression for stress-induced mutagenesis In Chapter II, Uracil-DNA glycosylase(ung) releases incorporation of dUTP or hydrolytic deamination of DNA cytosine residues. In Escherichia coli and in many other organisms, the enzyme catalyzes the first step in a repair pathway for uracil-containing DNA. Deamination of DNA cytosine to produce uracil is a frequent occurrence in the cell, the producing uracil base-pairs with adenine and causes. 2.
(8) G:C~A:T. transitions upon subsequent replication.. G:C~A:T. transitions were. stimulated about 30 fold in E. coli ung mutants. Therefore, molecular mechanism of ung transcriptional regulation is deeply related to the mutagenesis in E. coli. The expression of ung was strongly reduced in the stationary phase while the expression of cpxRA was induced. However, the molecular mechanisms were unknown. In this study, we demonstrated that the transcriptional mechanism of ung gene expression by CpxRlCpxA two-component signal transduction system in E. coli. In Chapter III, the ArcA/ArcB (anoxic redox control) two-component system is an important element in the complex transcriptional regulatory network that allows facultative anaerobic bacteria, such as Escherichia coli , to sense various respiratory growth conditions and adapt their gene expression accordingly (luchi & Lin, 1995; Malpica et ai., 2004). This system comprises the cytoplasmic response regulator ArcA and the transmembrane sensor kinase ArcB, ArcA is a typical response regulator possessing an N-terminal receiver domain with a conserved Asp residue at position 54 and a C-terminal helix-turn-helix DNA binding domain. In contrast, ArcB is an unorthodox sensor kinases as manifested by its unusually elaborate architecture. As a sensor, ArcB is deviant because in contrast to typical sensor kinases that have a substantial periplasmic sequence of only 16 amino acid residues delimited by two caninical transmembrane segments. Interestingly the ArcB transmembrane domain (amino acids 22-77) dose not directly participate in signal sensing but rather serves as an anchor that keeps the protein close to the source of the signal. As a kinase, ArcB is a typical because it contains three catalytic domains: an N-terminal transmitter domain with a conserved His-292 residue, central receiver domain with a conserved His-292 residue, a central receiver domain with a conserved Asp-576 residue, and a C-terminal. 3.
(9) phosphotransfer domain with a conserved His717 residue . Moreover, in the linker that is the region connecting the catalytic domains with the transmembrane domain, there are a putative leucine zipper and a Per-Amt-sim (PAS) domain (Taylor et at., 1999). Under reducing conditions, ArcB undergoes ATP-dependent autophosphorylation, a process shown to be enhanced by certain anaerobic metabolites such as D-lactate, acetate, and pyruvate (Georgellis et at., 1999, Rodriguez et at., 2004), and transphosphorylates ArcA via a His292. Asp-576---His717----Asp54 phosphoreray. (Georgellis et at., 1997, Kwon et ai., 2000), Phosphorylated ArcA(P-ArcA) represses the expression of many operons involved in respiratory metabolism and activates a few operons encoding proteins involved in fermentative metabolism (Iuchi & Lin, 1988, Brondsted et at., 1994, Lynch & Lin, 1996). Chapter III describes how DNA repair and replication genes, uvrA, ssb genes are transcriptionally regulated dependent on reducing conditions by ArcAIArcB two-component signal transduction system. In Chapter II and Chapter III, DNA repair and replication genes (ung, uvrA and ssb) expression were negatively controlled by the two-component systems CpxR/CpxA. and ArcAIArcB, respectively, revealing molecular mechanisms by which a specific stress dose in fact affect mutation in related genes (stress-induced mutagenesis).. 4.
(10) External stimuli. Fitness Adaptive response. - ...~? •. II. Mutation. Fig. 1 A model of stress-induced mutagenesis for adaptation to a new environment. 5.
(11) CHAPTER II. Negative Regulation of DNA Repair Gene (ung) Expression by CpxRlCpxA Two-component System in Escherichia coli K-12 and Induction of Mutations by Increased Expression of CpxR. Introduction The cpxRA two-component system of Escherichia coii, consisting of CpxA sensor kinase/phosphatase and CpxR cognate response regulator, regulates biofilm formation, motility and chemotaxis, host cell invasion, and virulence (De Wulf et ai., 1999; Dorel. et ai., 1999; Hung et ai., 2001; Otto & Silhavy, 2002). Using the generally accepted DNA sequence for binding CpxR (two tandem GTAAA motifs separated by a 5-bp linker, CpxR box), DeWulf et ai. (De Wulf et ai., 2002) performed 15-bp weighted matrix analysis for CpxR recognition sites on the entire E. coli genome and identified new target genes (ung, ompC, psd, mviA, aroK, rpoE, secA, and aer) that might be regulated positively or negatively by CpxRlCpxA. The expression of cpxRA increases sharply at the onset of the stationary growth phase (De Wulf et ai., 1999), suggesting the involvement of the Cpx system in stationary phase survival. Uracil-DNA glycosylase (Ung) , the most abundant type among all of the glycosylases in bacterial cells (Lutsenko et ai., 1999; Mokkapati et ai., 2001), excises uracil residues from DNA, which arise as a result of either misincorporation of dUTP by DNA polymerase or deamination of DNA cytosine (Duncan & Weiss, 1982; Duncan, 1985; Wyszynski et ai., 1994; Gallinali & Jiricny, 1996; Lutsenko & Bhagwat, 1999). The ung mutants show the specific mutator effect of a G:C-to-A:T transition as. 6.
(12) Duncan and Weiss (1982) reported previously. The ung gene expressIOn remams constant up to the early stationary phase of E. coli but declines in the late stationary phase (Varshney et al., 1988). However, the mechanisms underlying these phenomena are not understood well. In order to clarify the possible involvement of the Cpx system in repression of ung expression, the effects of CpxR overexpression on ung expression are investigated in this study.. Materials and Methods Bacterial strains and growth conditions. The E.coli strains used are listed in Table 1. They were grown at 37°C in a Luria-Bertani (LB) medium (pH 7.5) containing 1% Bacto Tryptone (Difco), 0.5% Bacto Yeast Extract (Difco), and 1% NaCl. The following. antibiotics. were. added. to. the. medium:. ampicillin,. 100. chloramphenicol, 25 Ilg/ml; tetracycline, 12.5 Ilg/ml; and kanamycin, 25 Ilg/ml.. 7. Ilg/ml;.
(13) TABLE 1. Bacterial strains and plasmids Strains or. Genotype I Relavant characteristics. Plasmids. a. Sourcel ). Reference. E.coli Straius BW25113 BW27559. lac]q, rrnB T14, dlacZ WJI6, hsdR5I4, daraBAD AH33 , MhaBAD LD78 BW25113, d (cpxR) 623. BL21. V ompT,hsdS B (rB-mB), gal (AcI857, indI, Sam 7 ,nin5, lac UV5-T7 genel ), dcm, (DE3). BD2314. ung-I52 ::TnIO, lacYI ,gal-6, trpC45 , his-68 ,purC50, lyrA 2 , rpsL125 , malAI ,xylA 7, mtl-2, thil [fiuA2, tsx70, supE44 (AS) I. BWuug b). CCI02. CCI02ung. Datsenko et al. Oshima et al . Novageu Duncan et al . BW25113 x. BW25113, ungI52 ::TnIO P90C ara d (lac proB )XIII I F' lacIZ, proB + CCI02, ung-152 ::TnIO. Pl(BD2314) -->Tc r Cupples et al. CCI02 x Pl(BD2314) -->Tc r. plasmids pCA24N pCA24dGFP pET-21a (+) pBAD18. H. Mori e ). Cmr, expression vector dGFP, pCA24N. H. Morid) Novagen. Apr, expression vector. p41-5dGFP. Apr, a vector containing the arabinose P BAD promoter Apr, Kmr, lacZ operon fusion vector sfil Fragment (698 bp) containing cpxR was cloned into the. pKH50-3. correspondiug sites of pCA24dGFP Bam HI-Not I fragment (698 bp) containing cpxR was cloned. pRS551. Guzman et al. R.W. Simons H.Aiba K. Yamamoto. into the corresponding sites ofpET-21a (+) pRSung. pRS551, Eco RI-Bam HI (538 bp) fragment containing ung. This study. promoter pBADnlpE. Eco RI-Xba I fragment (751 bp) containing nlpE was cloned iuto. This study. the corresponding site of pBAD18 a) b). Ap, ampicillin; Km, kanamycin; Cm, chloramphenicol; Tc, tetracycline G:C-A:T tester strain. e)http://ecoli.aist-nara.ac.jp/gb4/resources/archive/ACI.gif. d)Notl digestion aud selfligatiou ofpCA24N. Construction of plasmids. The plasmids used are listed in Table 1. A DNA. fragment (538 bp) containing the ung promoter region was prepared by polymerase chain reaction (PCR) using Escherichia coli BW25113 genome DNA as template and a pair of primers, ung-EcoRI-F (5'-TGGAACTTCACGGAATTCAATGTCA-3') and ung-BamHI-R (5'-AAAATAGGGATCCTGCTTCTCTTCA-3'). After digestion with. 8.
(14) EcoRI and BamHI, the PCR-amplified fragment was inserted at the corresponding site. ofpRS551 to generate the plasmid pRSung. A DNA fragment (751 bp) containing the nlpE coding region was prepared by PCR using Escherichia coli BW25113 genome. DNA as a template along with a pair of primers, NLPEAF (5'-ATGCGCGGCAGAATT CGCAGCGGTCGGGAA-3') and NLPEAR (5'-TGCGTTTGTTTCTAGATCAAGAC GGGTTAC-3'). After digestion with EcoRI and BamHI, the PCR-amplified fragment was inserted at the corresponding site of pBAD 18 to generate the plasmid pBADnlpE.. Preparation of the labeled probe for 81 nuclease, gel shift and DNase I footprinting assays. Probe A was generated by PCR amplification of the ung promoter. region with the primers, ung-SlF (5'-TGATGCCTCCCCGGCAAAAT-3') and 32P-Iabeled. ung-SlR (5'-TAGGGTTGCTGCTTCTCTTC-3'), and E.coli BW25113. genome DNA (100 ng) as the template for the Ex Taq DNA polymerase. The PCR product with 32 p at its terminus was recovered from a polyacrylamide gel and then used for S 1 nuclease, gel shift, and DNase I footprinting (non-coding strand) assays. The ung-SIR and labeled ung-SlF primers were used to prepare probe B for an SI nuclease assay (transcription of yfiD). Probe C was generated by PCR amplification of the cpxR promoter region with the primers cpxRA-SIF (5'-GTTATCGCCTGAACCGACTT-3') and. 32P-labeled. cpxRA-SIR (5'-GAAGCCTTCCATCTCGAGCA-3'), and E.coli. BW25113 genome DNA (100 ng) was used as the template for the Ex Taq DNA polymerase. The PCR product with. 32 p. at its terminus was recovered from a. polyacrylamide gel and then used for SI nuclease. The cpxRA-SIR and labeled cpxRA-SIF primers were used to prepare probe D for an SI nuclease assay (transcription of cpxP). The labeled primers were prepared with 10 /-lCi of [y_32 p ] ATP. 9.
(15) (5000Ci/mmol) by T4 polynucleotide kinase (Toyobo).. RNA isolation and Sl nuclease assay. To prepare total RNA for the S 1 nuclease assay, overnight cultures were diluted 100-fold in 100 ml of LB medium grown to an optical density at 600 nm (OD 6oo) of 0.6 or 1.2 at 37°C. Subsequent purification steps were carried out as described previously (Minagawa et at., 2003). Probe A (10000 cpm, 2 fmol) was incubated with 100. ~g. of total RNA in hybridization buffer (80%. formamide, 0.4 M NaCI, 20 mM HEPES [pH 6.4]) at 75°C for 10 min, followed by further incubation at 37°C overnight and then digested with 50 units of S 1 nuclease. The undigested DNA was precipitated by ethanol, dissolved in formamide dye solution (95% formamide, 0.05% bromophenol blue, 0.05% xylenecyanol), and analyzed by electrophoresis on a 6% polyacrylamide gel containing 8 M urea.. Purification of CpxR. His-tagged CpxR for Gel shift, DNase I footprinting and in vitro transcription assays was purified from pKH50-3/BL21 (DE3) as described. previously (Yamamoto et at., 2002).. Gel shift assay. Probe A (10000 cpm, 2 fmol) was incubated at 37°C for 10 min with CpxR in 12.5 ml of 50 mM Tris-HCI (PH 7.8),50 mM NaCI, 3mM Mg acetate, 0.1 mM EDTA, and O.lmM dithiothreitol. After addition of the DNA dye solution (40%. glycerol, 0.025% bromophenol blue, 0.025% xylenecyanol), the mixture was directly subjected to a 6% polyacrylamide gel electrophoresis.. DNase I footprinting assay. Either 40000 cpm of probe A or B (8 fmol) was. 10.
(16) incubated at 37°C for 10 min with CpxR in 25 III of 50 mM Tris-HCI (PH 7.8), 50 mM NaCI, 3 mM magnesium acetate, 5 mM CaCh, 0.1 mM EDTA, 0.1 mM dithiothreitol, and 25 mg/ml BSA. After incubation for 10 min, DNA digestion was initiated by the addition of 5 ng of DNase I (Takara). After digestion for 30 s at 25°C, the reaction was terminated by the addition of 45 III of DNase I stop solution (20 mM EDTA, 200 mM NaCl, 1% sodium dodecyl sulfate, 250 Ilg/ml yeast tRNA). Digested products were precipitated by ethanol, dissolved in formamide dye solution, and analyzed by electrophoresis on a 6% polyacrylamide gel containing 8 M urea.. In vitro transcription assay. A 538-bp EcoRI-BamHI fragment from pRSung, including the ung promoter region, or a 205-bp EcoRI fragment from pKB252 (Backman et al., 1976), including the lacUV5 promoter region, was used as the template DNA. Single-round transcription by the reconstituted holoenzymes was carried out as described previously (Kajitani & Ishihama, 1983). In brief, 0.1 pmol of template was incubated with 0, 1.25, 2.5, 5, and 10 pmol CpxR for 10 min at 37°C in the presence or absence of 10 mM acetylphosphate in a total volume of 33 Ill. Into this reaction mixture, 0.5 pmol RNA polymerase was added and incubated for 20 min at 37°C to form an open complex. Then a substrate/heparin mixture containing [a_ 32 P]UTP was added and further incubated for 10 min at 37°C. The transcripts were subjected to electrophoresis on a 6% polyacrylamide gel containing 8 M urea.. Assay of Ung activity.. Cell-free extract for exammmg Ung activity was. prepared as described previously (Lutsenko & Bhagwat, 1999). Here, 5 ml of an overnight culture of E.coli cells was inoculated into 200 ml of LB medium (pH 7.5). 11.
(17) containing 1 mM of IPTG and grown at 37°C until reaching OD 6oo of 1.2. After harvesting by centrifugation, the obtained pellet was washed with 50 ml of H20. The cells were again harvested by centrifugation, and the pellet was resuspended in 3 ml of a lysis buffer (50 mM Tris-HCl, pH 7.6, 100 mM NaCl, 10% glycerol, 1 mM dithiothreitol). After the cells were sonicated, the cell debris was removed by centrifugation at 15000 rpm for 20 min at 4°C, and the supernatant was used for assaying. Ung. activity.. The. DNA. oligonucleotide. containing. uracil. (oligo. Lac-461Ruracil, 5'-AGCGCCATGGCCTGACUCATTCCCCAGCGA-3') was labeled with. 32 p. at the 5' end by T4 polynucleotide kinase (Toyobo) and hybridized to the. unlabeled oligomer, LAC-461 F( 5' -TCGCTGGGGAATGGGTCAGGCCATGGCGCT3 '), at a molar ratio of 1: 10 to form a duplex with a single U:G mismatch. The duplex was separated with a 20% polyacrylamide gel electrophoresis and recovered. It was reacted with cell-free extract for 30 min at 37°C in a treatment buffer (20 mM Tris-HCl, 10 mM EDTA) and then treated with 0.1 M NaOH to cleave at the apyrimidine (AP) site for another 30 min at 37°C. After that, the products were separated by electrophoresis on a 20% polyacrylamide gel containing 8% urea and autoradiographed.. Lac+ mutation assay. To measure the mutation frequency from G:C to A:T,. a. spontaneous Lac+ mutation assay using E.coU CCI02 (Cupples & Miller, 1989) was performed as described previously (Mackay et al., 1994). A single colony was inoculated into 5 ml of LB medium containing 1 mM IPTG and grown with aeration for 16 h. The overnight culture was diluted 106-fold with minimal A salt (Miller, 1992). Then, 0.1 ml of the diluted culture was added to 10 ml of minimal A medium containing up to 0.2% glucose, 1 mM MgS0 4 , 0.0005% thiamine hydrochloride, tetracycline. 12.
(18) and/or chloramphenicol. After incubation for 16 h, the culture was harvested by centrifugation. The cell pellet was resuspended in 1 ml of minimal A salt, plated on a minimal A medium (0.2% lactose or 0.2% glucose), and incubated at 37°C. The mutation frequency was then calculated as the ratio of lac+ cells to 1 x 109 cells. Results and Discussion Effect of overexpression of CpxR on Ung activity. To determine whether the Cpx. system is involved in the regulation of ung expression, we examined the Ung activity in crude cell extracts from E. coli BW25113 coli BW25113. (p41-5~GFP). (p41-5~GFP). overexpressing CpxR. When E.. was incubated in the presence of IPTG, it was confirmed. that the cpxR transcript from the lac promoter on the plasmid was overexpressed (Fig. lA, lane 4). To measure Ung activity, we used a model substrate consisting of a synthetic polynucleotide of 30 nucleotides in length containing a single uracil residue at position 17 from the 5' end. When cell extracts from E. coli BW25113. (pCA24~GFP). without CpxR expression were used, the labeled polynucleotide (30 mer) containing uracil was cleaved after treatment with 0.1 M NaOH to form a 16-mer oligonucleotide and two other bands as cleaved DNA products (Fig. 2A, lanes 2, 3). These bands were not detected in the absence or presence of the cell extracts from BWung (Fig. 2A, lanes1, 6). When CpxR was induced by addition oflPTG in BW25113. (p41-5~GFP). (Fig. 2A,. lanes 4, 5), Ung activity decreased to about fivefold lower than that of BW25113 (pCA24~GFP). (Fig. 2A, lanes 2, 3), implying that the Cpx system repressed ung. expression.. 13.
(19) 123 4. Fig. 1. Overexpression of cpxR. The arrow indicates cpxR transcript from lac promoter on the plasmid p41-5L'.GFP. BW25113 (pCA24L'.GFP) (lanes I, 2) and BW25113 (p41-5L'.GFP) (lanes. 3,4) were grown to mjd-Iog pbase (OD600, 0.6) in the absence (lanes I, 3) or presence (lanes 2, 4) of I mM IPTG. SI nllclease assay for cpxR transcript was perfonned as described in Materials and Metbods.. 14.
(20) (A) 3. 2. 1. 4. 6. .. 30 mer. .. 16 mer (B) 100 90 ~ 80 011 70 ~ ('$ 60. ~. U. ~. Q. SO 40 30 20 10 0 1. 2. 3. 6. Fig. 2. Assay of Uracil-DNA glycosylase (Ung) activity. (A) Ung activ ity was assayed as described in Materials and Methods. For this assay, cell-free extracts (lanes 2 and 4, 25 ng; lanes 3 and 5, 50 ng) from BW25 I 13. (pCA24~GFP). (lanes 2 and 3), BW25 I I 3. (p41-5~GFP). (lanes. 4 and. 5) and BWung (lane 6, 50 ng) were used. Thirty-mer oligonucleotide was c leaved by Ung and NaOH to form 16-mer product. In lane I, no ce ll -free extract was used. (B) C leaved DN A products (F ig. 2A) were quantified with BAS 1000 Mac (Fuj i fi lm). Percent cleavage was detennined by dividing the intensity of the c leaved product ( 16 mer) by the total intens ity, which was defined as the sum of the intensiti es of the intact substrate and the cleaved products.. 15.
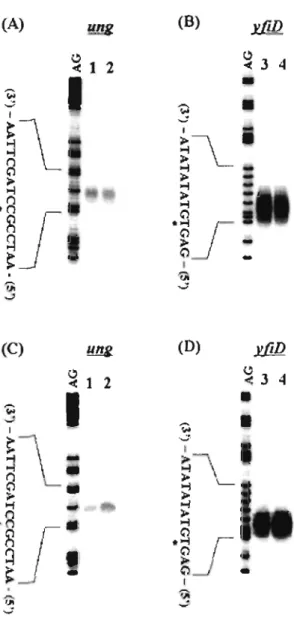
(21) As described previously (Duncan & Weiss, 1982), a G:C-to-A:T transition was stimulated in E. coli ung mutants (CC102ung) (Table 2). We investigated the effects of CpxR overexpression on the mutator activity of the G:C-to-A:T transition by using a spontaneous Lac+ mutation assay (Mackay et al., 1994). The results in Table 2 indicate that the G:C-to-A:T transition frequency for strain CCI02. (p41-5~GFP),. CpxR, was stimulated to about sixfold more than that for CC 102. overexpressing. (pCA24~GFP).. TABLE 2. G:C--+A:T mutation frequency Strain CCI02 CCI02ung pCA24~GFP / CCI02 p41-5~GFP / CCI02. No of Lac+ colonies per 109 cells· (±SD) 1.7 (± 0.021) 5.9 (± 0.253) 1.8 (± 0.085) 11.2 (± 1.060). "Each value represents the average ofthe data from three independent experiments. SD: standard deviation. Negative regulation of ung gene expression by overexpression of CpxR. To investigate the effect of CpxR overexpression on the transcription of the ung gene, we performed an S 1 nuclease assay using RN As prepared from CpxR-expressing BW25113 (p41-5~GFP). as well as BW25113. (pCA24~GFP).. When CpxR was overexpressed by. addition of IPTG (Fig. lA, lane 4; Fig. 3A, lane 4; Fig. 3B, lane 4), the transcription of ung decreased by 5-fold (Fig. 3A, lane 4) compared to that ofBW25113. (pCA24~GFP). (Fig. 3A, lane 2 ) while cpxP increased by lO-fold (Fig. 3B, lane 6 and 8). In addition, we examined whether overproduction of the outer membrane lipoprotein NlpE can repress the ung expression (Fig. 3C, lane 4), since overproduction ofNlpE activates the Cpx signal transduction pathway (Snyder et al., 1995; Danese & Silhavy, 1997; DiGiuseppe & Silhavy, 2003). When nlpE expression was induced by arabinose, the ung transcription was repressed by 5-fold (Fig. 3C, lane 4) compared to that of. 16.
(22) BW25113 (pBAD 18) (Fig. 3C, lane 2), while the transcriptional level of cpxP increased by lO-fold (Fig. 3D, lane 6 and 8). Neither IPTG nor arabinose repressed ung and cpxP transcriptions in BW25113 (data not shown), therefore these results suggest that the ung expression was negatively regulated by the Cpx pathway.. 17.
(23) (A). ~,. '<" 1. ~. (B). Il1lK. '<" 5 6. 2 J 4. 7 8. ~. ...::. ,. ~L. ~L. n~ ·n. .>. L r --. Lf. I. ...:: '". ~. -:;. (0). ( ) ~. ~. « 1 2 J 4. < 5 6 7 8. ~. .... ....... I. ~L'. ~. I. (). > ..;. •. •. ~L ~. 1. ..;. ~f. ~J. I. ~. V;. -:;. ..J. Fig. 3. Effects of overexpressed CpxR or NlpE on ling and cpxP transcription.. BW25113. (pCA246.GFP) (A, B, lanes I , 2, 5, and 6) and BW25113 (p4 1-56.GFP) (A, B, lanes 3, 4, 7 and 8) were grown to mid-log pbase (OD 600 , 0.6) in the absence (A , B, lanes I , 3, 5, and 7) or presence (A, B, lanes 2, 4, 6 and 8) of I mM IPTG. S I nuclease assays for ung (A) and cpxP (B) transcripts were performed as described in Materials and Methods. BW25113 (pBAD 18) (C, D, lanes I , 2, 5, and 6) and BW25113 (pBADnlpE) (C, D, lanes 3, 4, 7 and 8) were grown to mid-log phase (OD 600 , 0.6) in the absence (C, D, lanes I , 3, 5, and 7) or presence (C, D, lanes 2, 4, 6 and 8) of 0.2% arabinose. After that, S I nuclease assays for ung (C) and cpxP (D) transcripts were performed. Lane AG represents the Maxam-Gilbert sequence ladder. Transcription start site is marked with an asterisk.. 18.
(24) To confinn this result, we next examined the influence of cpxRA deletion on the expression of the ung gene. In wild-type BW25113 cells, the level of ung mRNA decreased in the stationary phase (Fig. 4C, lane 1), but the level of yfiD transcription did not change independently of the growth phase (Fig. 4B and D). The growth phase-coupled reduction in ung transcription was not observed in strain BW27559 (Fig. 4A and C, lane 2). These results indicate that CpxR negatively regulates ung transcription in the stationary phase and that this repression does not take place in the absence of the Cpx system. To further confinn the role of CpxR in the transcription of the ung gene, we perfonned in vitro transcription using reconstituted RNA polymerase holoenzyme E a. 70. and truncated DNA fragments (538 bp) containing the promoter region of ung in the presence or absence of CpxR. Addition of CpxR caused repression of ung transcription, while it did not affect the expression from the lacUV5 promoter (Fig. 5A). Such a repression was stimulated in the presence of 10 mM acetylphosphate (Fig. 5B, lanes 5-8).. 19.
(25) (8). (A). ~ J 4. ~ 1 2. (0). (C). )!fiD. ~ 3 4. Fig. 4. ling and yfiD gene expression dependent on growth phase. E. coli B W25113 (A, C lane I and B, 0 lane 3) and BW27559 (/!'cpxR) (A , C lane 2, and B, Diane 4) were grown to mid-log phase (OD600=0.6) (A, B) or earl y stationary phase (0 0 600 = 1.2) (C, D). After that, S l nuclease assays for ung (A , C) and yjiD (B, D) transcripts were performed. Lane AG represents the Maxam-Gilbert seq uence ladder. Transcription start site is marked with an asterisk.. 20.
(26) (C). (A). ..... 1.2. .ung. 1. l.fIeUVS.. 0.8. 8II. ~. 0.6 0 .... 1 2 3. ~. S 6 7 8. 0.2. (B). 0. US. 2.5. 3.7S. CpxR(pmor). . • uIIg. lac. 5.. 2 3. ~. S 6 7 8. Fig. 5. In vitro transcription assay. Single-round transcription in vitro of 0. 1 pmol ung DNA template (-452 to +86 of promoter region, A lanes 5-8, B lanes 5-8) or laeUV5 DNA template (A lanes 1-4, B lanes 1-4) was performed in the presence (B) or absence (A) of 10 mM acetyl phosphate. The amounts of CpxR were as follows: lanes I and 5, 0 pmol; lanes 2 and 6, 1.25 pmol ; lanes 3 and 7, 2.5 pmol ; lanes 4 and 8, 5 pmol. Electrophoresis was performed with a 6% polyacrylamide sequencing gel. Bold arrows indicate the ung and laeUV5 transcripts. (C) ung (circles) and laeUV5 (square) transcripts in the absence (open symbols) and presence (c losed symbols) were quantified with BAS 1000 Mac (Fuji film) . The relative value in shown as a ratio between each transcript and that in the absence of CpxR.. 21.
(27) Identification of the CpxR-hinding site. To define the mechanism underlying the repression of ung transcription by CpxR, we tested whether CpxR directly interacts with the ung promoter region. For this purpose, we carried out gel shift assays using the 32P-labeled DNA fragment containing the ung promoter region. In the absence of acetylphosphate, a marked shift was observed only at higher concentrations of CpxR (Fig. 6A, lanes 4-6). In contrast, a significant gel shift was observed even at the lowest concentration of CpxR when 25 mM acetylphosphate was added (Fig. 6A, lanes 8-12). CpxR is a member of the OmpR family of winged helix-turn-helix transcription factors. We investigated the binding ability of another OmpR family protein, PhoP, and that of Bovine serum albumin to the ung promoter. No interaction of protein-ung promoter DNA was detected (Fig. 6B). These results indicate that the affinity of CpxR to the CpxR box of the ung promoter region is increased by phosphorylation.. 22.
(28) (A). (B) . A(~fylphosph.ale. +Acd)'lphCl<spllaCt. _ . . . . .S.PhoP .. 1. 2. 3. 4. 5. 6. 7. 8. 9 10 11 12. 1. 2. 3. 4 !!. __...... C.BSA. 6. 7. 8. 9. .10. Fig. 6. Gel shift assay. (A) Probe A was incubated at 37°C for 10 min with CpxR (lanes 1-6) or CpxR phosphorylated by acetyl phosphate at 37°C (lanes 7-12). The amounts of CpxR were as follows: lanes I and 7, 0 pmol ; lanes 2 and 8, 1.25 pmol ; lanes 3 and 9, 2.5 pmol; lanes 4 and 10, 5 pmol ; lanes 5 and 11 , 7.5 pmo l; lanes 6 and 12, 10 pmol. (B) Probe A was incubated at 37°C for 10 min with PhoP (lanes 1-5) or BSA (lanes 6-10) at 37°C. The amounts of PhoP or BSA were as follows: lanes I and 6, 0 pmol ; lanes 2 and 7, 1.25 pmol ; lanes 3 and 8, 2.5 pmol; lanes 4 and 9, 5 pmol ; lanes 5 and 10, 10 pmol). 23.
(29) Next, we tried to identify the specific CpxR-binding site on the ung promoter region by using a DNase I footprinting assay. CpxR was found to bind to the CpxR box containing the direct repeat between -61 and -86 upstream from the transcriptional start position (Fig. 7). The CpxR box in the ung promoter is located 61 bp upstream of the transcription start site (Fig. 8) and corresponds with the site predicted by DeWulf et al (2002). It is well known that the C-terminal domain of the a subunit of RNA polymerase (a-CTD) interacts with an AT-rich sequence (UP-element) located upstream of the -35 hexamer (Ross et al., 1993). The CpxR box in the ung promoter also contains an AT-rich sequence corresponding to the UP-element. CpxR binding to the ung promoter may repress its transcription by interfering with the ability of the a-CTD to bind to the UP-element. In Salmonella enterica, the transcriptional factor PmrA binding to the 59 bp upstream site in the pmrD promoter represses pmrD transcription by interfering with the ability of the a-CTD to interact with a putative UP element that overlaps the AT-rich PmrA binding site (Kato et ai., 2003). CpxR seems to repress the. ung expression in a similar manner as shown for the pmrD promoter (Kato et ai., 2003). Taken together, these results indicate that the ung gene transcription is directly controlled by phosphorylated CpxR.. 24.
(30) Fig. 7. DNase I footprinting assay. Probe A was incubated with various amounts of the purified CpxR (lane 1, 0 pmol; lane 2, 10 pmol ; lane 3, 20 pmol ; lane 4, 30 pmol ; lane 5, 40 pmol ; lane 6, 50 pmol ; lane 7, 60 pmol ; lane 8, 70 pmol ; lane 9, 80 pmol) and subjected to DNase I footprinting assays. Lanes AG represent the Maxam-Gilbert sequence ladder. The black boxes and bold arrows indicate CpxR binding region and the direct repeat, respectively.. 25.
(31) -90. -80. -70. -60. -50. -40. +1 0. +10. +10. I I I I 5'-TATGCGGGTGTGAAATTTTACC CTCGTTTAAGTCTAAAAATGAGdATGAT'li 3'-ATACGCCCTCACTTTAAAATGGT-':':======::I_GAGCAAATTCAGATTTTTACTCGTACTAA. -)0. -10. +1. I. -If. TTGTTCTGTAGAAAGAAGC~GTTAAQcTA~CGGATTGAAGATTCGCAGGAGAGCGAGATGGCTAACGA-5' AACAAGACATCTTTCTTCGTCAATTCGATCCGCCTAACTTCTAAGCGTCCTCTCGCTCTACCGATTGCT-3'. Fig. 8. ung promoter and CpxR box. Transcription start site is marked with an asterisk. The nucleotide number represents the distance from the transcription initiation site of the ung promoter. Putative recognition sequences for a. 70. (-10 and -35) are boxed. The arrows indicate the direct. repeat in the CpxR box shown previously (De Wulf et al., 2002). In the present study, we demonstrated negative regulation of ung expression by overexpression of CpxR (see Fig. 2, 3). Transcription of the ung gene at the stationary phase is about 3-fold higher in the cpxRA deletion strain than in the parental strain (see Fig. 4C). DeWulf et al. (2002) predicted positive control of the ung, ompC, psd, mviM, aroK, and secA genes by phosphorylated CpxR and negative control of rpoErseABC and aero In this paper, we suggest that ung is expressed in the exponential growth phase but. decreases in the stationary phase concomitantly with the increase in phosphorylated CpxR. This finding indicates that ung is a target of negative control by CpxR. Furthermore, we also found an increase in the mutation level of E. coli overexpressing CpxR. The increased mutation might be attributable, at least in part, to the repression of ung expression by the increased CpxR. This is because it is not possible to repair DNA uracil residues, which arise as a result of either misincorporation of dUMP by DNA polymerase or deamination of DNA cytosine residues. This finding. 26.
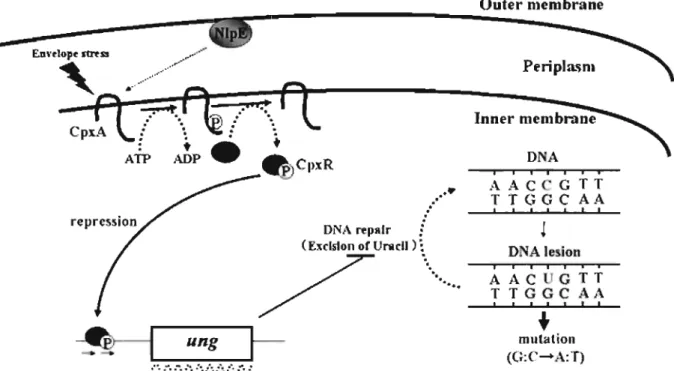
(32) further implies that the CpxR/CpxA two-component system plays a role in controlling the mutation rate.. Outer membrane. Peripta.sm. CpxA. :. '.. :. Inner. ~. A~ jP. •. D A. pxR. / ....•. TT. DNA repair : (Excision or Un ell ) ~. / Fig. 9.. ruemb"'D~. D. '" '. ...... AAC GrT TrGGC A ,. ,. ,. ,. ,. I. ,. +. mutation ( : ....A :T). A model of negative regulation of ung gene expression by CpxR/CpxA. two-component system.. 27.
(33) CHAPTER III. Negative regulation of DNA repair gene (uvrA) expression by ArcA/ArcB two-component system in Escherichia coli. Introduction The Arc AIArcB two-component system (TCS) of Escherichia coli regulates the expression of about 30 operons that respond to redox conditions (Tardat & Touati, 1993; Iuchi et ai., 1994; Compan & Touati, 1994; Drapal & Sawers, 1995; Nystrom. et ai., 1996; Lynch & Lin, 1996; Chao et ai., 1997; Pellicer et ai., 1999). In addition to these redox-responsive genes, at least 3 targets that are not involved in the respiratory metabolism have been found to be under the control of ArcAiArcB. These targets are the tra operon for plasmid conjugation, the psi site for Xer-based recombination, and the. oriC site for chromosome replication (Colloms et ai., 1998; Strohmaier et ai., 1998; Lee et ai., 2001). Recently, Liu and De Wulf (2004) identified, using a DNA micro array and the AlignACE program, as many as 55 new Arc-regulated operons under anaerobic growth conditions, indicating that the Arc-controlled pathway recruits a total of 100-150 operons for adaptation to the availability of external oxygen (Liu & De Wulf, 2004). On the other hand, using a DNA micro array, Oshima et ai. (2002) performed a systematic examination of mRNA profiles for a total of 36 mutants, each lacking one two-component system, under aerobic conditions and found that the ArcAiArcB system showed the most drastic changes in transcription pattern (http://ecoli.aist-nara.ac.jp/xp_ analysis/2_components), which suggested that the gene (uvrA) involved in DNA repair is regulated by ArcA/ArcB TCS. uvrA encodes the nucleotide excision repair protein. 28.
(34) UvrA and is induced as part of the SOS response (Backman et al., 1976). Gifford et al. (2000) demonstrated that when cells were shifted from anaerobic to aerobic growth, the transcript levels of the uvrA increased. These results suggested that the ArcAlArcB two-component system should plays a role in controlling the uvrA gene in response to the respiratory growth conditions independent of the SOS system. Thus, we tried to clarify the detailed molecular mechanism of transcriptional regulation of the uvrA gene by ArcAiArcB TCS.. Materials and Methods Bacterial strains and growth conditions. E. coli BW25113 (factI, rrnBT14, L1 lacZwJ16, hsdR514, L1araBADAH33, L1 rhaBADLD78) (Datsenko &. Wanner, 2002),. BW26422 (BW25113, L1(arcB) 41), and BW27422 (BW25113, L1(arcA) 43) (Oshima et al., 2002) were cultured in a Luria-Betani (LB) medium (pH 7.5) containing 1% Bacto. Tryptone (Difco), 0.5% Bacto Yeast Extract (Difco), and 1% NaCI for RNA preparation. BL21(DE3) (F, ompT, hsdSB, (rB-,mB-), dcm, gal, A(DE3)) was cultured in the LB medium to express ArcA. These were grown at 37°e. When necessary, the following antibiotics were added to the medium: ampicillin, 100 I-lg/ml; kanamycin, 25 I-lg/ml.. Construction of plasmids. A DNA fragment (530 bp) containing the uvrA promoter region was prepared by polymerase chain reaction (PCR) using Escherichia coli BW25113 genome DNA as a template and a pair of primers, uvrA-EcoRI-F (5'-CCTTTACGCAGGAATTCGCTCGCCA-3') and uvrA-BamHI-R (5'-TTGTCGC GGGGGATCCCGAGGTTGA-3 '). After digestion with EcoRI and BamHI, the PCR-amplified fragment was inserted at the corresponding site ofpRS551 (Simons et ai,. 29.
(35) 1987) to generate the plasmid pRSuvrA. A DNA fragment (827 bp) containing the arcA coding region was prepared by PCR using Escherichia coli BW25113 genome DNA as a template along with a pair of primers, arcA-BAD-EcoRI-F (5'-GACTTTTGGAATT CCTGTTTCGATT-3') and arcA-BAD-XbaI-R (5'-GAGTTACGTATCTAGAAATAA GGTT-3'). After digestion with EcoRI and XbaI, the PCR-amplified fragment was inserted at the corresponding site of pBAD18 (Guzman et al., 1995) to generate the plasmid pBADarcA. The plasmid pKH50-3 (Yamamoto et al., 2004) containing Bam HI - NotI fragment (698 bp) of ArcA, was cloned into the corresponding sites of pET-21 a. (+) (Novagen).. Preparation of the labeled probe for gel shift, SI nuclease and DNase I footprinting assays. Probe A was generated by PCR amplification of the uvrA promoter region with the primers, uvrA-SIF (5'-GTCCTGACCCAGATTACCAA-3') and 32P-Iabeled uvrA-SIR (5'-CGAGGTTGATGTTTTTGAGA-3'), and E. coli BW25113 genome DNA (100 ng) as the template for the Ex Taq DNA polymerase. The PCR product with 32p at its terminus was recovered from the polyacrylamide gel after electrophoresis and then used for gelshift assay, SI nuclease assay (transcription of uvrA), and DNase I footprinting assay. The uvrA-SIR and labeled uvrA-SIF primers. were used to prepare probe B for an SI nuclease assay (transcription of ssb) and DNase I footprinting assay. The mfd-S1F (5'-GCATTCAGCGATCTGTTGT-3') and 32p labeled mfd-S1R (5'-GGCGTGACGTTCGGCAATTT-3') primers were used to prepare probe C for SI nuclease assay (transcription of mfd). The cydA-SIF (5'-GGTCAAC CGTGCTGTTTTTGCTTCGT-3') and 32 p labeled cydA-SIR (5'-GCTCATCGCATG AAGACTCCGAGAGT-3') primers were used to prepare probe D for gelshift assay and. 30.
(36) Sl nuclease assay (transcription of cydA). The labeled primers were prepared with 10 ).lCi of [y_32 p ] ATP (5000 Ci/mmol) by T4 polynucleotide kinase (Toyobo) as described previously (Minagawa et at., 2003).. RNA isolation and SI nuclease assay. To prepare total RNA for the S 1 nuclease assay, overnight cultures were diluted 100-fold in the LB medium and grown under both aerobic and anaerobic conditions. In the aerobic growth condition, cells were grown in the LB medium at 37°C by reciprocal shaking (160 rpm, stroke: 3 cm) in a 300-ml round-bottom flask (lwaki Co., Japan) containing 50 ml of medium. In the anaerobic condition, cells were grown in the LB medium at 37°C under cap-sealed conditions in a 300-ml narrow-mouth flask (lwaki Co., Japan) containing 300 ml of medium. Cells were harvested during the logarithmic growth phase (OD6oo=0.4) to prepare total RNA. Subsequent purification steps were carried out as described previously (Minagawa et at., 2003). Probe A, B, C, or D (10000 cpm, 2 fmol) was incubated with 100 ).lg of total RNA in hybridization buffer (80% forrnamide, 0.4 M NaCI, 20 mM HEPES [pH 6.4]) at 75°C for 10 min, followed by further incubation at 37°C overnight and then digested with 50 units of S 1 nuclease. The undigested DNA was precipitated by ethanol, dissolved in forrnamide dye solution (95% forrnamide, 0.05% bromophenol blue, 0.05% xylenecyanol), and analyzed by electrophoresis on a 6% polyacrylamide gel containing 8 Murea.. Purification of ArcA. His-tagged ArcA for DNase I footprinting and in vitro transcription assays was purified from pKH50-3/BL21 (DE3) as described previously (Yamamoto et ai., 2002).. 31.
(37) Gel shift assay. Probe A, B, or D (10000 cpm, 2 fmo1) was incubated at 37°C for 10 min with ArcA in 12.5 m1 of 50 mM Tris-HC1 (pH 7.8),50 mM NaCl, 3 mM Mg acetate, 0.1 mM EDTA, and 0.1 mM dithiothreitol. After addition of the DNA dye solution (40% glycerol, 0.025% bromophenol blue, 0.025% xylenecyanol), the mixture was directly subjected to 6% polyacrylamide gel electrophoresis.. DNase I footprinting assay. Either 40000 cpm of probe A or B (8 fmo1) was incubated at 37°C for 10 min with ArcA in 25 ).11 of 50 mM Tris-HC1 (PH 7.8), 50 mM NaCI, 3 mM magnesium acetate, 5 mM CaCh, 0.1 mM EDTA, 0.1 mM dithiothreito1, and 25 mg/m1 BSA. After incubation for 10 min, DNA digestion was initiated by the addition of 5 ng of DNase I (Takara). After digestion for 30 s at 25°C, the reaction was terminated by the addition of 45 ).11 of DNase I stop solution (20 mM EDTA, 200 mM NaCI, 1% sodium dodecy1 sulfate, 250 ).1g/m1 yeast tRNA). Digested products were precipitated by ethanol, dissolved in formamide dye solution, and analyzed by electrophoresis on 6% polyacrylamide gel containing 8 M urea.. In vitro transcription assay. A 530-bp EcoRI-BamHI fragment from pRSuvrA, including the uvrA promoter (-128 to +402), or a 205-bp EcoRI fragment from pKB252 (Backman et al., 1976), including the lacUV5 promoter (-65 to +140), was used as the template DNA. Single-round transcription by the reconstituted ho10enzymes was carried out as described previously (Kajitani & Ishihama, 1983). Briefly, 0.1 pmol of template was incubated with 0, 2.5, 5, 10, and 20 pmo1 ArcA for 10 min at 37°C in a total volume of 33 ).11. Into this reaction mixture, 0.5 pmo1 RNA polymerase was added and incubated. 32.
(38) for 20 mm at 37°C to form an open complex. Then a substrate/heparin mixture containing [y}2 p ]UTP was added and further incubated for 10 min at 37°C. The transcripts were subjected to 6% polyacrylamide gel containing 8 M urea for electrophoresis.. Results and Discussion Negative regulation of uvrA gene expression by ArcA/ArcB two-component system. To isolate the DNA repair and replication genes regulated by the ArcAIArcB two-component system, we first selected the uvrA gene as an up-regulated gene in arcB-defective strains using DNA micro array analysis data (http://ecoli.aist-nara.ac.jp/. xp_analysis/2_components, Oshima et al., 2002). To confirm this micro array data, we performed Sl nuclease assays of RNAs prepared from BW27422 (BW25113 LlarcA), BW26422 (BW25113 LlarcB), and the wild-type strain (BW25113) under both aerobic and anaerobic growth conditions. As a result, transcription of uvrA increased in BW27422 and BW26422 more than that in BW25l13, which was remarkable under the anaerobic growth condition (Fig. 1 A, B). On the other hand, transcription of cydA decreased in BW27422 and BW26422 more than in BW25113 (Fig. 1 C, D), as described previously (Govantes et ai., 2000). These results indicated that transcriptions of uvrA were negatively regulated by the ArcAlArcB two-component system, which was remarkable under the anaerobic condition.. 33.
(39) TABLE 1 The DNA repair and replication genes regulation by the TCS in E. eoli I). TCS. genes. ( 1.92, 1.54 ). epxRA. nei. ( 0.54, 0.51 ). hsdR. ( 0.48, 0.54 ). kdpDE. mutS. ( 0.39, 0.53 ). ada. ( 0.06, 0.11 ). reeF. ( 2.53, 1.90 ). hsdR. ( 0.43, 0.40 ). rstAB. dinP. ( 2.09, 1.53 ). narXL. ntpA. ( 2.06, 1.59 ). torSR. reeB. ( 0.48, 0.44 ). narQ. ntpA. ( 1.63, 1.53 ). eusRS. merA. ( 1.92, 2.27 ). ntrBC. hsdR. ( 1.84, 1.62 ). jimZ. mutH. ( 0.54, 0.40 ). areA. priC. ( 1.62, 1.59 ). cheABY. topB. ( 1.53, 1.76 ). areB. hsdR. ( 0.45, 0.41 ). ntpA. ( 1.60, 1.63 ). uvrA. ( 2.18 11.91jJ. basSR. hsdR. ( 1.59, 1.52 ). mfd. ( 1.91, 1.50 ). yjhA. merA. ( 0.43, 0.25 ). phrB. ( 1.81, 1.63 ). TCS. genes. rssB. aidB. ompR/envZ. 1) Expression ratio of normalized signal intensities of Cy5-dUTP (TCS mutant) against Cy3-dUTP (Wild type, BW25113) from DNA microarray analysis (http://ecoli.aist-nara.ac.jp/xp_ analysis/2 _components). (8). ( ). (C). .... (0). ~. Q..,. ~. ]r~~. r'!}. E~. 1(~ ~ :#. I.A. Q. <. ]. 2. J. "". t.o. · ~I. ~. if. if. ~. < 4. S. 6. >. -i l#. 0. -i. -;-I. '"'". J i.). < 1 2 J. b~ ~. a. f'~J ~. n. ~. • <-'. < ... 5. '-'!. Fig. 1. Effect of respiratory condition on llvrA and cydA gene ex pressions. BW25113 (A, B, e, 0 , lanes 1 and 4), BW27422 (t.arcA) (A, B, e, 0 , lanes 2 and 5), and BW26422 (t.arcB) (A, B, e, 0 , lanes 3 and 6) were grown to early- log phase (0 0 600 =0.4) under an aerob ic (Janes I, 2, and 3) or anaerob ic (Janes 4, 5, and 6) cond iti on. After that, S 1 nuclease assays for uvrA (A, B) and cydA (e , 0 ) transcripts were perfonned. Lane AG represents the Maxam-Gilbert sequence ladder. The transcription start site is marked with an asterisk.. 34. 6.
(40) Negative regulation of overexpressed AreA on uvrA gene expression. To confinn the negative effect of ArcA on uvrA gene expreSSIOn, we next examined overproduction of ArcA under the aerobic growth condition. In BW25113 containing vector pBAD 18, the level of uvrA transcript did not change in the presence or absence of arabinose (Fig. 2A, lanes 1 and 2). When ArcA was overexpressed in BW25113 containing pBADarcA, the transcription of mid encoding the transcription-repair coupling factor was not affected (Fig. 2B, lanes 3 and 4); however, the transcription of uvrA was markedly repressed compared to that in the absence of arabinose (Fig. 2A,. lanes 3 and 4). These results indicated that transcriptions of uvrA were negatively regulated by overexpressed ArcA, even under the aerobic growth condition.. ( ). 1. 2. J. ~. .1. 2. J. ~. Fig. 2. Effect of over ex pressed ArcA on lIrvA transcription. BW2S 113 (pBAO 18) (A, B, lanes I and 2) and BW2S113 (p BAOarcA) (A, B, lanes 3 and 4) were grown to early-l og ph ase (00 600 , 0.4) in the absence (A, B, lanes 1 and 3) or presence (A, B, lanes 2 and 4) of 0.2% arabinose. S J nuclease assays for uvrA (A) and mid (B) transcripts were perfonned as described in Materia ls and Methods. Lane AG represents the Maxam-Gilbert seq uence ladder. The transcription start site is marked w ith an asterisk.. 35.
(41) Effects of DTT on uvrA gene expression. Under reducing conditions, ArcB undergoes ATP-dependent autophosphorylation and transphosphorylates ArcA (Malpica et at., 2004, Iuchi & Lin, 1995). Phosphorylated ArcA (P-ArcA) represses the. expression of many operons in the respiratory metabolism and a few operons encoding proteins involved in the fermentative metabolism such as the cydA gene. The addition of thiol-reducing agent Dithiothreitol (DTT) to the aerobic culture of a wild- type strain led to an immediate increase in the expression of the P-ArcA-activatable cydA-tacZ reporter (Malpica et at., 2004). To show that uvrA gene expression is regulated under reducing conditions (e.g. in the presence ofDTT), we investigated the effects ofDTT on uvrA gene expression (Fig. 3). As a result, when the wild- type E. coli was incubated in the presence of 10 mM DTT aerobically, uvrA gene expression was inhibited (Fig. 3, lane 2), but cydA gene expression was increased (Fig. 3, lane 2). Furthermore, gelshift experiments resulted in enhancement of ArcA-promoter complexes for uvrAIssb and cydA promoter fragments in the presence of acetylphosphate (Fig. 4). These results suggested that negative regulation of uvrA gene expression is caused by ArcA phosphorylated by ArcB activated under reducing conditions. In the previous study, when cells were shifted from anaerobic to aerobic growth, the transcript levels of the oxidative DNA glycosylases (mutY, jjJg, nei, and nth) and uvrA increased (Gifford et at, 2000), but the molecular. mechanism was unknown. Our findings strongly indicated that ArcAIArcB TCS directly inhibits the uvrA expression under a reducing condition in E. coli. These results imply that the AreAlArcB TCS could play a role in controlling the DNA repair under the respiratory conditions of E. coli.. 36.
(42) ( ). (B) I.;. ,.. I.;. o.r~l. r· ~. ~J. i. " > 0-1 > 0-1. 0. n. f;. n n. ~. 0. 0-1. •. -;-i. ~. ~. l. J. 1 2. Fig. 3. Effects of DTT on lIvrA gene expression. 8W25113 was grown aerobically in an L8 medium containing 0. 1 M MOPS (PH 7.4) and 20 mM O-xylose to early-log phase (00 600=0.4) in the absence (lane I) or presence (lane 2) of 10 mM OTT. S I nuclease assay for uvrA (A) or cydA (8) transcript was performed as described in materials and methods. Lane AG represents the Maxam-Gilbert sequence ladder.. uv -acetyl P. - =e=:::=::1. 2. J. ,.. +acetyl P. -acetyl P. 5. AreA. 6. 8. \I 10. ArcA. 1. ~. 1. ". 5. 6. 7. 8. \I. 10. Fig. 4. Gel shift assays. (A) Probe A for uvrA promoter was incubated at 37°C for 10 min with ArcA (lanes 1-5) or ArcA phosphorylated by acetyl phosphate at 37°C (lanes 6-10). The amounts of ArcA were as follows: lanes I and 6, 0 pmol ; lanes 2 and 7, 1.25 pmol ; lanes 3 and 8, 2.5 pmol; lanes 4 and 9, 5 pmol ; lanes 5 and 10, 10 pmol (8) Probe 0 for cydA promoter was incubated at 37°C for 10 min with ArcA (lanes 1-5) or ArcA phosphorylated by acetyl phosphate at 37°C (lanes 6-10). The amounts of ArcA were as follows: lanes I and 6, 0 pmol ; lanes 2 and 7, 1.25 pmol ; lanes 3 and 8, 2.5 pmol ; lanes 4 and 9, 5 pmol ; lanes 5 and 10, 10 pmol.. 37.
(43) Effects of ArcA on ssb transcription. The ssb gene encoding single-strand DNA-binding protein is located next to uvrA but transcribed in the opposite direction (see Fig. 7C). Both uvrA and ssb genes are organized in the SOS regulon, and are regulated by the LexA repressor (Backendorf et al., 1983; Brandsma et ai., 1983). We then examined the possible influence of ArcA on ssb transcription. In the early log-phase cells of BW27422 lacking areA, the level of ssb mRNA was apparently the same as that of BW25113 (data not shown). From S1-mapping, we detected three transcripts, indicating the presence of three promoters (P 1, P2 and P3 in this order from downstream) (Fig. 5). When ArcA was overexpressed in BW25113 (pBADarcA) by addition of 0.2% arabinose, the transcription from ssbP3 decreased to an undetectable level, and that from ssb p1 decreased by 3-fold (Fig. 5, lane 4) compared to that of BW25113 (pBAD18) (Fig. 5, lane 2). These results indicate that the Arc system also represses transcription from the ss b promoter.. 38.
(44) Fig. 5. Effect of overexpressed ArcA on ssb transcription. BW25113 (pBAD 18) (lanes I and 2) and BW251 13 (pBADarcA) (lanes 3 and 4) were grown in LB to early-log phase (OD 600 =0.4) in the absence (lanes I and 3) or presence (lanes 2 and 4) of 0.2% arabinose. S I nuclease assay was performed as described in Materials and Methods.. In vitro transcription of uvrA and ssb. For detailed analysis of the repreSSlOn mechanism of uvrA and ssb transcription by ArcA, in vitro transcription using RNA polymerase holoenzyme Ecr 70 and truncated DNA templates containing either the uvrA or ssb promoter region were carried out in the presence or absence of ArcA. The addition of ArcA significantly caused repression of transcription from the uvrA promoters in a dose-dependent manner but did not affect the expression from the. lacUV5 promoter (Fig. 6). In good agreement with the in vivo observations (see Fig. 5), transcription of ssb was also repressed by ArcA (Fig. 6). The addition of ArcA caused marked repression of transcription from ssb P3 and caused weaker repression of transcription from ssb p1 and ssb n. 39.
(45) - - .... -. +.s.TbP3. " ___. III .. _. -. ~.s.TbPl. ~lt"TA. -. a- aaa +lacUVS 1 :2 3 .. 5. Fig. 6. In vitro transcription assay. Single-round transcription in vitro was performed with a 0.1 -pmol uvrA -ssb DNA template (-128 to +402 ofuvrA promoter region) and a lacUVS DNA template. The amounts of ArcA were as follows: lane I, 0 pmol ; lane 2, 2.S pmol ; lane 3, S pmol ; lan e 4, 10 pmol ; lane S, 20 pmol. Electrophoresis was performed with a 6% polyacrylamide sequencing gel. Bold arrows indicate the uvrA, ssbPJ, ssbf'2, ssb P3 and lacVVS transcripts.. Identification of AreA-binding sites by DNase I footprinting. To define the mechanism underlying the repression of uvrA and ssb transcription by ArcA, we tried to identify the specific ArcA-binding site on these promoters by using DNase I footprinting assay. ArcA was found to bind to the uvrA promoter between -19 and +16 (AI: ArcA site-I; -56 and -90 with respect to ssb P3 promoter) (Fig. 7A). These ArcA binding regions include the ArcA-binding motif 5'-NTGTTAATTAA/T-3' (Lynch & Lin, 1996). Detailed footprinting analysis of ArcA on the ssb promoters indicated additional ArcA-binding sites at -80 and -95 (A2: ArcA site-2; between - 61 and - 76 with respect to the uvrA promoter), between -41 and -54 (A3: ArcA site-3; -102 and - 115 with respect to the uvrA promoter), and between -1 and -10 (A4: ArcA site-4; -146 and - 155. 40.
(46) with respect to the uvrA promoter) upstream from the ssb p \ transcriptional start position (Fig. 7B). All of these regions include the ArcA-binding motif 5' -NTGTTAATTAAlT3' (Lynch & Lin, 1996), showing that the divergently transcribed ssb and uvrA genes are controlled by these ArcA-binding sites.. (C). (B). (A). .1< so-. 1<. I~. -70-. .'0-so-. -teo-. . .. 0 -lO-. -!'O80-. 10-. -70-. -to-. '0-. ' I~. I~. . .. 0-. - ~o-. .lo-i-..... it-'.. I~. + 10- ... . t!I I 2 3 ~. <. t!I I 2 3. <. Fig. 7. AreA-binding sites on the uvrA and ssb promoter region. (A) DNase I footprinting assay of the ArcA to the uvrA-ssb promoter region . Probe A (Panel A) or probe B (panel B) was incubated with variou s amounts of the purified ArcA (lane I , 0 pmol; lane 2, 20 pmol ; lane 3, 40 pmol; lan e 4, 80 pmol) and subjected to DNase I footprinting assays. Lane AG represents the Maxam-Gilbert sequence ladder. The black boxes indicate ArcA binding region. (C) Transcription start sites are marked with asterisks and arrows . The nucleotide number represents the distance from the transcription initiation sites of the uvrA, ssbP I promoters. The LexA binding region is boxed with the dotted line. The ArcA binding region is also boxed; the bold line indicates the ArcA binding consensus region (5'-AlTGTTAATTANT-3').. 41.
(47) Using DNA mlcroarrays (http://ecoli.aist-nara.ac.jp/xp_ analusis/2 _ components/), many DNA repair and replication genes regulated by two-component systems containing CpxRlCpxA and ArcA/ArcB were found (Oshima et at., 2002). Among them, we focused on CpxRlCpxA (CHAPTER II) and ArcAlArcB two-component system to clarify the transcriptional regulation of DNA repair and replication gene expressions. First, we found that ung (Uracil-DNA glycosylase) is expressed in the exponential growth phase but decreased in the stationary phase (CHAPTER II). These indicated that ung is a target of negative control by CpxRlCpxA. In this paper, both uvrA and ssb. genes were also negatively regulated dependent on reducing conditions by ArcA/ArcB two-component system. Thus, DNA repair and replication gene expressions (ung, uvrA and ssb) were negatively controlled by the two-component systems CpxRlCpxA and. ArcAiArcB under anaerobic or stationary growth conditions. Recent understanding of stress-induced mutagenesis or adaptive mutagenesis in microbes indicates that mutational rates can be elevated in response to stress, producing beneficial and other mutations (Biedov et. at., 2003, Hersh et at.,. 2004).. However such a molecular mechanism still remained obscure. Two-component systems including CpxRlCpxA (CHAPTER II) and ArcAlArcB may playa important role of the molecular mechanisms by which a specifc stress under anaerobic or stationary growth conditions affects such a mutation rate.. 42.
(48) ,. (a). (b). Anaerobiosis. Aerobiosis .MmiIiiQililJi!iii,. .rniQliliGlliliRiI!' ,. Periplasm. Periplasm. AreA. Cytoplasm. Cytoplasm. AreB. ADP. p. 1 ssb. 1 PiuvrAI--I--. ---1. p. ~. ssb. H. uvrA I~-. ,. Fig. 8. A model of negative regulation of uvrA and ssb gene expression by A rcA/A reB two-component system.. 43.
(49) Summary. Chapter II In Escherichia coli K-12 overexpressing CpxR, transcription of the ung gene for uracil-DNA glycosylase was repressed, ultimately leading to the induction of mutation. Gel shift, DNase I footprinting, and in vitro transcription assays all indicated negative regulation of ung transcription by phosphorylated CpxR. Based on the accumulated results, we conclude that ung gene expression is negatively regulated by the two-component system of CpxRlCpxA signal transduction (Fig. 1).. Chapter III We first selected the uvrA gene as an up-regulated gene in arcB-defective strains using. DNA. microarray. analysis. data. (http://ecoli.aist-nara.ac.jp/xp analysis/2. components). To confirm this microarray data, we performed S 1 nuclease assays of RNAs prepared from BW27422 (BW25113 LiarcA), BW26422 (BW25113 8arcB), and the wild-type strain (BW25113) under an aerobic or anaerobic growth condition. As a result, transcription of uvrA increased in BW27422 and BW26422 more than in BW25113, which was remarkable under the anaerobic condition. In BW25113 grown in the presence of DTT (10 mM) or containing overexpressed ArcA, the expressions of uvrA gene were repressed. Furthermore, the results of in vitro transcription, gel shift. assay and footprinting experiments indicate that phosphorylated ArcA specifically binds to the ArcA box [(AlT)GTTAATTA(AlT)] in the uvrA promoter and represses its transcription. These results strongly suggest that uvrA gene expression is negatively regulated by an AreAlArcB two-component system under reducing conditions. (Fig. 1). 44.
(50) G:U--+G:C. ung '\fIIt./WN\N. .. ~. repair. ~. t uvr.. ~ Stress (stationary or. Stationary phase. reducing condition). y....•. ••... 7'X. .tepreSSion :. •. •. ~. mutagenesis .-------~. ung. utagenesis uvrA. T. ~. •. reducing condition. Fig. 1 Stress-induced mutagenesis controlled by TCS 45.
(51) ACKNOWLEGMENTS. I wish to express my sincerest gratitude to Professor Ryutaro Utsumi, Graduate School of Agriculture, Kinki University, for kind guidance, valuable suggestion and discussions, and continuous encouragement throughout the course of this work and critical reading of the manuscript. Special thanks are due to Professors Yukio Kawamura and Shigeru Shigeoka, Graduate School of Agriculture, Kinki University, for reading the entire text in its original form. I also wish to express my sincerest acknowledgment to Assosiate Professor Hiroyuki Tanabe, and Assistant Professor Kaneyoshi Yamamoto, for their useful discussions and technical advices. I am also deeply indebted to Dr. Akira Ishihama, Nippon Institute for Biological Science, for valuable suggestions and discussions thoughout the work of transcription regulation of ung, uvrA, and ssb. I would also like to thanks to Dr. R. W. Simons, for supplying plasmid, pRS551; Dr Hirotada Mori, for providing the cpxR gene,. p41-5~GFP;. Dr. Akiko Nishimura, for. supplying plasmid, pBAD 18; Dr. Hirofumi Aiba, for providing the TCS mutants, BW26422. (~arcB),. BW27422. (~arcA),. BW27559. (~cpxRA);. Dr. B. K. Duncan, for. providing the ung mutant, BD2314; Dr. l. H. Miller, for supplying. G:C~ A:T. tester. strain, CC 102. Many thanks are due to Dr. Akinori Kato, Mr. lun Teramoto, Miss Kiyo Hirao, Mr. Satoshi Yamamoto, Miss lunko Sasaki, Mr Yusuke Sato, Miss Mayumi Kubo, Miss Risa Hayashi, Dr. Takafumi Watanabe, Mr. Shu Minagawa, Mr. Eiji Furuta, Mr. Tomoki. 46.
(52) Nakanishi and other members of Department of Agricultural Chemistry, Kinki University. Finally, very special thanks are forwarded to my parents for their continuous support and cooperation, without which this work was never possible.. 47.
(53) REFFERENCES. Backendorf, C., J. A. Brandsma, T. Kartasova, and P. van de Putte. 1983. In vivo regulation of the uvrA gene: role of the "-10" and "-35" promoter regions.. Nucleic. Acids Res. 11: 5795-5810. Backman, K., M. Ptashne, and W. Gilbert. 1976. Construction of plasmids carrying the cI gene of bacteriophage A. Proc. Natl. Acad. Sci. USA 73: 4174-4178. Bjedov, I., O. Tenaillon, B. Gerard, V. Souza, E. Denamur, M. Radman, F. Taddei, and I. Matic. 2003. Stress-Induced mutagenesis in Bacteria. Science 300: 1404-1409. Brandsma, J. A., D. Bosch, C. Backendorf, and P. van de Putte. 1983. A common regulatory region shared by divergently transcribed genes of the Escherichia coli SOS system. Nature 305: 243-245. Chao, G, J. Shen, C. P. Tseng, S. J. Park, and R. P. Gunsalus. 1997. Aerobic regulation of isocitrate dehydrogenase gene (icd) expression in Escherichia coli by the. arcA andfnr gene products. J. Bacteriol. 179: 4299-4304 48.
(54) Colloms, S. D., C. Alen, and D. J. Sherratt. 1998. The ArcAiArcB two-component regulatory system of Escherichia coli is essential for Xer site-specific recombination at psi. Mol. Microbiol. 28: 521-530. Compan, I. and D. Touati. 1994. Anaerobic activation of arcA transcription m. Escherichia coli: roles ofFnr and ArcA. Mol. Microbiol. 11: 955-964. Cupples, C. G and J. H. Miller. 1989. A set oflacZ mutations in Escherichia coli that allow rapid detection of each of the six base substitutions. Proc. Natl. Acad. Sci. USA 86: 5345-5349. Danese, P. N. and T. J. Silhavy. 1997. The a E and the Cpx signal transduction systems control the synthesis of periplasmic protein-folding enzymes in Escherichia. coli. Genes Dev. 11: 1183-1193. Datsenko, K. A. and B. L. Wanner. 2000. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 97: 6640-6645. 49.
(55) DeWulf, P., O. Kwon, and E. C. C. Lin. 1999. The CpxRA signal transduction system of Escherichia coli: growth-related auto activation and control of unanticipated target operons. J. Bacteriol. 181: 6772-6778. DeWulf, P., A. M. McGuire, X. Liu, and E. C. C. Lin. 2002. Genome-wide profiling of promoter recognition by the two-component response regulator CpxR-P in. Escherichia coli. J. BioI. Chern. 277: 26652-26661. DiGiuseppe, P. A. and T. J. Silhavy. 2003. Signal detection and target gene induction by the CpxRA two-component system. J. Bacteriol. 185: 2432-2440. Dorel, C., O. Vidal, C. Prigent-Combaret, I. Vallet, and P. Lejeune. 1999. Involvement of the Cpx signal transduction pathway of E.coli in biofilm formation. FEMS Microbiol. Lett. 178: 169-175. Drake, J. W. 1991. Spontaneous mutation. Annu. Rev. Genet. 25: 125-146. 50.
(56) Drapal, N. and G Sawers. 1995. Purification of ArcA and analysis of its specific interaction with the pjl promoter-regulatory region. Mol. Microbiol. 16: 597-607. Duncan, B. K. and B. Weiss. 1982. Specific mutator effects of ung (uracil-DNA glycosylase) mutations in Escherichia coli. J. Bacteriol. 151: 750-755. Duncan, B. K. 1985. Isolation of insertion, deletion, and nonsense mutations of the uracil-DNA glycosylase (ung) gene of Escherichia coli K-12. J. Bacteriol. 164: 689-695. Foster, P. L. 1999. Mechanisms of stationary phase mutation: a decade of adaptive mutation. Annu. Rev. Genet. 33: 57-88. Gallinali, P. and J. Jiricny. 1996. A new class of uracil-DNA glycosylases related to human thymine-DNA glycosylase. Nature (London) 383: 735-738. Georgellis, D., A. S. Lynch, and E. C. C. Lin. 1997. In vitro phosphorylation study of the ArcA two-component signal transduction system of Escherichia coli. J. Bacteriol. 179: 5429-5435. 51.
(57) Georgellis, D., O. Kwon, and E. C. C. Lin. 1999. Amplification of Signaling activity of the Arc two-component system of Escherichia coli by anaerobic metabolites. An in vitro study with different protein modules. J. BioI. Chern. 274: 35950-35954. Gifford, C. M., J. O. Blaisdell, and S. S. Wallace. 2000. Multiprobe RNase protection assay analysis of mRNA levels for the Escherichia coli oxidative DNA glycosylase genes under conditions of oxidative stress. J. Bacteriol. 182: 5416-5424. Govantes, F., A. V. Orjalo and R. P. Gunsalus. 2000. Interplay between three global regulatory proteins mediates oxygen regulation of the Escherichia coli cytochrome d oxidase (cydA) operon. Mol. Microbiol. 38: 1061-1073. Guzman, L.-M., D. Belin, M. J. Carson, and J. Beckwith. 1995. Tight regulation, modulation, and high-level expression by vectors containing the arabinose PBAD promoter. J.Bacterioi. 177: 4121-4130. 52.
(58) Hersh, M. N., R. G Ponder, P. J. Hastings, and S. M. Rosenberg. 2004. Adaptive mutation and amplification in Escherichia coli: two pathways of genome adaptation under stress. Res. Microbiol. 155: 352-359. Hoch, J. A., and T. J. Silhavy. 1995. Two-component signal transduction (Am. Soc. Microbiol., Washington, DC). Hung, D. L., T. L. Raivio, C. H. Jones, T. J. Silhavy, and S. J. Hultgren. 2001. Cpx signaling pathway monitors biogenesis and affects assembly and expression of P pili. EMBO J. 20: 1508-1518. Iuchi, Sand E. C. C. Lin. 1995. Signal transduction in the Arc system for control of operons encoding aerobic respiratory enzymes. Two-component signal transduction system (Am. Soc. Microbiol., Washington, DC). 223-231. Iuchi, S., and E. C. C. Lin. 1998. arcA (dye), a global regulatory gene in Escherichia coli mediating repression of enzymes in aerobic pathways. Proc. Natl. Acad. Sci. USA. 85: 1888-1892. 53.
(59) Iuchi, S., A. Aristarkhov, J. M. Dong, J. S. Taylor, and E. C. C. Lin. 1994. Effect of nitrate respiration on expression of the Arc-controlled operons encoding succinate dehydrogenase and flavin-linked L-Lactate dehydrogenase. J. Bacteriol. 176: 1695-1701. Kajitani, M. and A. Ishihama. 1983. Determination of the promoter strength in the mixed transcription system: promoters of lactose, tryptophan and ribosomal protein L 10 operons from Escherichia coli. Nucleic Acids Res. 11: 671-686. Kato, A., T. Latifi, and E. A. Groisman. 2003. Closing the loop: The PmrAlPmrB two-component system negatively controls expression of its posttranscriptional activator PmrD. Proc. Natl. Acad. Sci. USA 100: 4706-4711. Kwon, 0., D. Georgellis, A. S. Lynch, D. Boyd, and E. C. C. Lin. 2000. The ArcB sensor kinase of Escherichia coli: genetic exploration of the transmembrane region. J. Bacteriol. 182: 2960-2966. 54.
(60) Lee, Y. S., J. S. Han, Y. Jeon, and D. S. Hwang. 2001. The Arc two-component signal transduction system inhibits in vitro Escherichia coli chromosomal initiation. 1. BioI. Chern. 276: 9917-9923. Liu, X. and P. De Wulf. 2004. Probing the ArcA-P modulon of Escherichia coli by whole-genome transcriptional analysis and sequence-recognition profiling. J. BioI. Chern. 279: 12588-12597. Lombardo, M-J. I. Aponyi, and S. M. Rosenberg. 2004. General stress response regulator RpoS in adaptive mutation and amplification in Escherichia coli. Genetics 166: 669-680. Lutsenko, E. and A. S. Bhagwat. 1999. The role of the Escherichia coli Mug protein in the removal of uracil and 3,N4-ethenocytosine from DNA. J. BioI. Chern. 274: 31034-31038. Luria, S. E., and M. Delbruck, 1943. Mutations of bacteria from virus sensitivity to virus resistance. Genetics 28: 491-511. 55.
(61) Lynch, A. S. and E.C.c. Lin. 1996. Transcriptional control mediated by the ArcA two-component response regulator protein of Escherichia coli: Characterization of DNA binding at the target promoters. J. Bacteriol. 178: 6238-6249. Mackay, W. J., S. Han, and L. D. Samson. 1994. DNA alkylation repair limits spontaneous base substitution mutation in Escherichia coli. 1. Bacteriol. 176: 3224-3230. Makinoshima, H., A. Nishimura, and A. Ishihama. 2002. Fractionation of Escherichia coli cell populations at different stages during growth transition to stationary phase. Mol. Microbiol. 43: 269-279. Malpica, R., B. Franco, C. Rodriguez, O. Kwon, and D. Georgellis. 2004. Identification of a quinone-sensitive redox switch in the ArcB sensor kinase. Proc. Natl. Acad. Sci. USA 101: 13318-13323. Mckenzie, G J., R. S. Harris, P. L. Lee, and S. M. Rosenberg. 2000. The SOS response regulates adaptive mutation. 97: 6646-6651. 56.
(62) Miller, J. H. 1992. In J. H. Miller (ed.), A short course in bacterial genetics. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N. Y.. Minagawa, S., H. Ogasawara, A. Kato, K. Yamamoto, Y. Eguchi, T. Oshima, H. Mori,. A.. Ishihama,. and. R.. Utsumi.. 2003.. Identification. and. molecular. characterization of the Mg2+-stimulon of Escherichia coli. J. Bacteriol. 185: 3696-3702. Mokkapati, S. K., A. R. Fernandez de Henestrosa, and A. S. Bhagwat. 2001.. Escherichia coli DNA glycosylase Mug: a growth-regulated enzyme required for mutation avoidance in stationary-phase cells. Mol. Microbiol. 41: 11 0 1-1111. Nystrom, T., C. Larsson, and L. Gustafsson. 1996. Bacterial defense against aging: role of the Escherichia coli ArcA regulator in gene expression, readjusted energy flux and survival during stasis. EMBO J. 15: 3219-3228. Otto, K. and T. J. Silhavy. 2002. Surface sensing and adhesion of Escherichia coli controlled by the Cpx-signaling pathway. Proc. Natl. Acad. Sci. USA. 99: 2287-2292. 57.
(63) Mori, H., K. Isono, T. Horiuchi, and T. Miki. 2000.. Functional genomlcs of. Escherichia coli in Japan. Res. Microbiol. 151: 121-128. Oshima, T., H. Aiba, Y. Masuda, S. Kanaya, M. Sugiura, B.L. Wanner, H. Mori, and T. Mizuno. 2002. Transcriptome analysis of all two-component regulatory system mutants of Escherichia coli K-12. Mol. Microbiol. 46: 281-291. Pellicer, M. T., C. Fernandez, J. Badia, J. Aguilar, E. C. C. Lin, and L. Baldom. 1999. Cross-induction of glc and ace operons of Escherichia coli attributable to pathway intersection. J. BioI. Chern. 274: 1745-1752. Rodriguez, C., O. Kwon, and D. Georgellis. 2004. Effect of D-Iactate on the physiological activity of the ArcB sensor kinase in Escherichia coli. J. Bacteriol. 186: 2085-2090. Rosenberg, S. M. 2001 Evolving responsively: adaptive mutation. Nat. Rev. Genet. 2: 504-515. 58.
(64) Ross, W., K. K. Gosink, J. Salomon, K. Igarashi, C. Zou, A. Ishihama, K. Severinov, and R. L. Gourse. 1993. A third recognition element in bacterial promoter: DNA binding by the a subunit of RNA polymerase. Science 262: 1407-1413. Shimada, T., H. Makinoshima, Y. Ogawa, T. Miki, M. Maeda, and A. Ishihama. 2004. Classification and strength measurement of stationary-phase promoters by use of a newly developed promoter cloning vector. J. Bacteriol. 186: 7112-7122. Simons, R. W., F. Howman, and N. Kleckner. 1987. Improved single and multicopy lac-based cloning vectors for protein and operon fusions. Gene 53: 85-96. Snyder, W. B., L. J. B. Davis, P. N. Danese, C. L. Cosma, and T. J. Silhavy. 1995. Overproduction of NlpE, a new outer membrane lipoprotein, suppresses the toxicity of the Cpx signal transduction pathway. J. Bacteriol. 177: 4216-4223. 59.
(65) Strohmaier, H., R. Noiges, S. Kotschan, G Sawers, G Hogenauer, E. L. Zechner, and. G. Koraimann.. 1998.. Signal. transduction. and. bacterial. conjugation:. Characterization of the role of ArcA in regulating conjugative transfer of the resistance plasmid Rl. J. Mol BioI. 277: 309-316. Sugiura, M., H. Aiba, and T. Mizuno. 2003. Identification and classification of two-component systems that affect rpoS expression in Escherichia coli. Biosci. Biotechnol. Biochem. 67: 1612-1615. Taddei. F., I. Matic, and M. Radman. 1995. cAMP-dependent SOS induction and mutagenesis in resting bacterial populations. Proc. Natl. Acad. Sci. USA. 92: 11736-11740. Tardat, B. and D. Touati. 1993. Iron and oxygen regulation of Escherichia coli MnSOD expression: competition between the global regulators Fur and ArcA for binding to DNA. Mol. Microbiol. 9: 53-63. 60.
(66) Taylor, B. L., and I. B. Zhulin. 1999. PAS Domains: Internal Sensors of Oxygen, Redox Potential, and Light. Microbioi. Mol. BioI. Rev. 63: 479-506. Tsui, H.. c., G. Feng, and M. F. E. Winkler, 1997. Negative regulation of mutS and. mutH repair gene expression by the Hfq and RpoS global regulators of Escherichia coli. K-12. J. Bacteriol. 179: 7476-7487. Varshney, U., T. Hutcheon, and J. H. van de Sande. 1988. Sequence analysis, expression, and conservation of Escherichia coli uracil DNA glycosylase and its gene (ung). 1. BioI. Chern. 263: 7776-7784. Wright, B. E. 2004. Stress-directed adaptive mutations and evolution. Mol. Microbiol. 52: 643-650. Wyszynski, M., S. Gabbara, and A. S. Bhagwat. 1994. Cytosine deaminations catalyzed by DNA cytosine methyltransferases are unlikely to be the major cause of mutational hot spots at sites of cytosine methylation in Escherichia coli. Proc. Natl. Acad. Sci. USA. 91: 1574-1578. 61.
(67) Yamamoto, K., H. Ogasawara, N. Fujita, R. Utsumi, and A. Ishihama. 2002. Novel mode of transcription regulation of divergently overlapping promoters by PhoP, the regulator of two-component system sensing external magnesium availability. Mol. Microbiol. 45: 423-438. Yamamoto K, K. Hirao, T. Oshima, H. Aiba, R. Utsumi, and Ishihama A. 2005. Functional characterization in vitro of all two-component signal transduction systems from Escherichia coli. J. BioI. Chern. In press. 62.
(68) PUBLICATIONS. Ogasawara, H., Teramoto, J., Hirao, K., Yamamoto, K., Ishihama, A., Utsumi, R., "Negative regulation of DNA repair gene (ung). expression by CpxRlCpxA. two-component system in Escherichia coli K-12 and induction of mutations by increased expression ofCpxR." ,J. Bacteriol., 186,8317-8325,2004. Ogasawara, H., Teramoto, J., Yamamoto, S., Hirao, K., Yamamoto, K., Ishihama, A., Utsumi, R., "uvrA and priC genes involved in DNA repair and replication are negatively regulated by ArcA in Escherichia coli K-12." , submitted to 1. Bacteriol.. Yamamoto, K., Ogasawara, H., Fujita, H., Utsumi, R., Ishihama, A., "Novel mode of transcription regulation of divergently overlapping promoters by PhoP, the regulator of two-component system sensing external magnesium availability.", Mol. Microbiol., 45, 423-438, 2002.. Minagawa, S., Ogasawara, H., Kato, A., Yamamoto, K., Eguchi, Y., Oshima, T., Mori, H., Ishihama, A., Utsumi, R., "Identification and molecular characterization of the Mg2+ stimulon of Escherichia coli.", J. Bacteriol., 185,3696-3702,2003.. 63.
(69)
図



+7




Outline
関連したドキュメント
Second, it was revealed that ADAR1-mediated RNA editing positively regulates DHFR expression in human breast cancer-derived MCF-7 cells by destroying miR- 25-3p and miR-125a-3p
PD-L1 expression was upregulated in macrophages and dendritic cells (DCs) in high-grade invasive human OSCC tissues or co-cultured with mesenchymal-phenotype OSCC cells in
After the precursor for miR-223 was transfected into HepG2 cells, the miR-223 levels were strikingly increased (40-Ct value: 21.1), similar to the levels observed with HeLa cells,
Gene expression levels and promoter usage of NR4A family NGFIB, NURR1, NOR1, NR5A1 and CYP11B2 in human cardiovascular and adrenal tissues.. a mRNA expression levels
Consistent with this, the knockdown of ASC expression by RNA interference in human monocytic/macrophagic cell lines results in reduced NF-κB activation as well as diminished IL-8
In humans, three types of TFF (TFF1–3) and their characteristic and co- ordinated distribution together with MUC mucin have been reported. That is, a combination of TFF1 with MUC5AC
Two novel mutations, both affecting highly conserved amino acids and one previously reported mutation responsible for PXE have been identified. To the best of our knowledge, this is
Treatment with ONO-1301 increased hepatic HGF mRNA expression, but decreased the expressions of TGF-β1, connective tissue growth factor, α-smooth muscle actin, and type-I and
