つくばリポジトリ SR 8 1 1282
全文
図
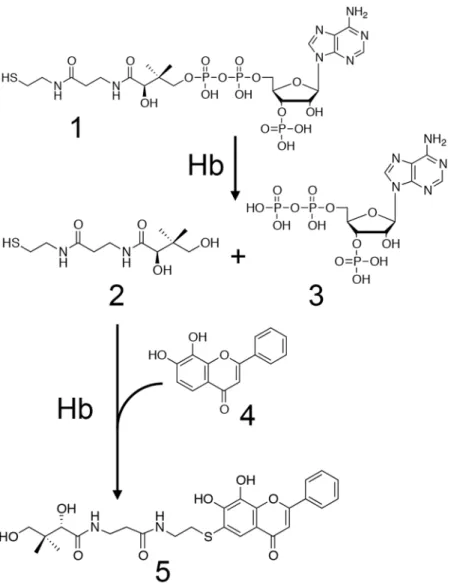
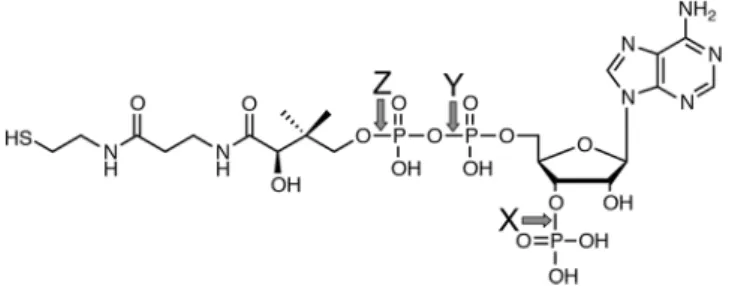
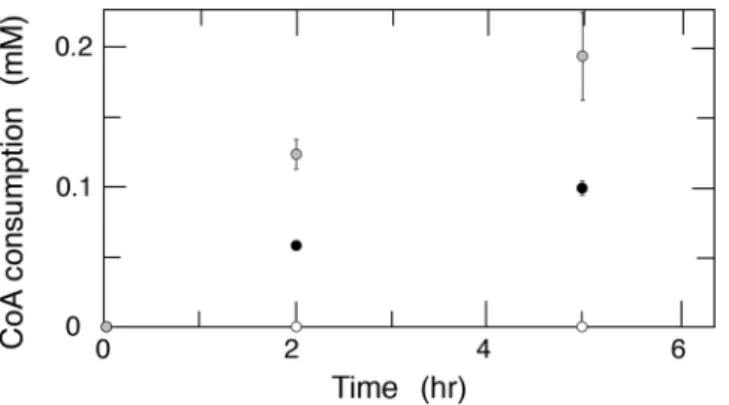
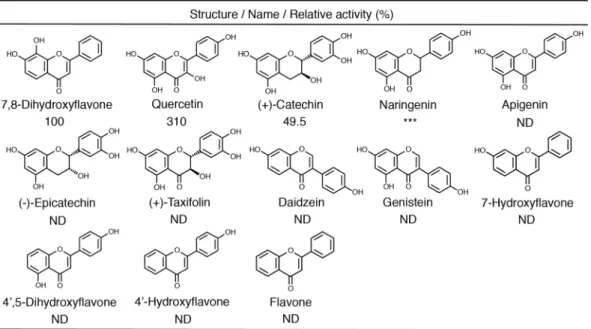
関連したドキュメント
Expression plasmids for JunB, c-Jun and JunD were each co-transfected with c-Fos expression plasmids as well as a luciferase reporter plasmid containing the wild-type LCR of
In immunostaining of cytokeratin using monoclonal antibodies, the gold particles were scattered in the cytoplasm of the hepatocytes and biliary epithelial cells
Determination of the Levels of Phosphorylated MAPK and GTP-bound Rac1—J774[SR-BI] cells or HEK293 cells forcedly expressing rat SR-BI and FLAG-tagged human GULP were incubated in
But the only places that carets might need to be added in order for multiplication by a generator to take place are as follows (each of these added carets are described by
Standard domino tableaux have already been considered by many authors [33], [6], [34], [8], [1], but, to the best of our knowledge, the expression of the
The approach based on the strangeness index includes un- determined solution components but requires a number of constant rank conditions, whereas the approach based on
In this work we give definitions of the notions of superior limit and inferior limit of a real distribution of n variables at a point of its domain and study some properties of
If in the infinite dimensional case we have a family of holomorphic mappings which satisfies in some sense an approximate semigroup property (see Definition 1), and converges to
