動物のアクチンの分子進化
Molecular Evolution of the Animal Actin
大堀 兼男
*OHORI Kaneo
Ⅰ.はじめに Ⅱ.アクチンの検索 Ⅲ.各種生物のアクチン Ⅳ.アクチンの進化 Ⅴ.まとめ 要約 アクチンは広く様々な生物に存在している。このアクチンは細胞骨格として細胞内に存在し、細 胞の形態維持や細胞内輸送において重要な役割を果たしている。また、動物では筋肉の構成成分と して、運動で重要な役割を果たしている。このようなアクチンについて、主に動物を対象に分子レ ベルでの進化を検討した。 キーワード:アクチン、進化、動物 Ⅰ.はじめに 運動は動物の最も重要な特徴であり、この 機能を担っているのが筋肉組織1)である。筋 肉組織の主成分としてアクチン2)とミオシン からなる収縮タンパク質のグループが存在す る。アクチンは筋収縮に働くタンパク質とし て発見されたが、その後、細胞内に存在する 細胞骨格成分の一つであることがわかった。 アクチンは細胞内において、細胞の形態維持、 細胞内物質輸送、アメーバ運動、細胞分裂、 接着、膜胞の代謝などにおいて重要な役割を 果たしていることが明らかにされた。 筋肉組織や細胞骨格では、アクチンが直線 的に重合して2本の繊維を形成していること が一般的である。アクチン繊維が主成分であ る筋肉繊維では、アクチン繊維にモータとし て働くミオシンが結合しており、ミオシンの 移動に伴い筋肉繊維の収縮が起こる。また、 アクチン繊維が細胞骨格の一つとなり、細胞 の形態を決定している。このとき、アクチン 繊維の重合が起きている。また、このアクチ ン繊維を通して物質の移動も起きている。 アクチンは菌類、植物、動物などを含む真 核生物に広く分布しているが、細菌にも見つ かっている。細菌ではアクチンの類似度は低 いが、真核生物ではその類似度が高く80%も あり、進化的には保守的である。このような アクチンが主に動物でどのように進化してき たのか検討する。研究ノート
* 本学経営学部 特任教授 1) 動物の筋肉系の進化については、ウィルマー (1990)、Schmidt-Rhaesa(2007)を参照のこと。2) ア ク チ ン に つ い て は、Pollard & Cooper (2009)、
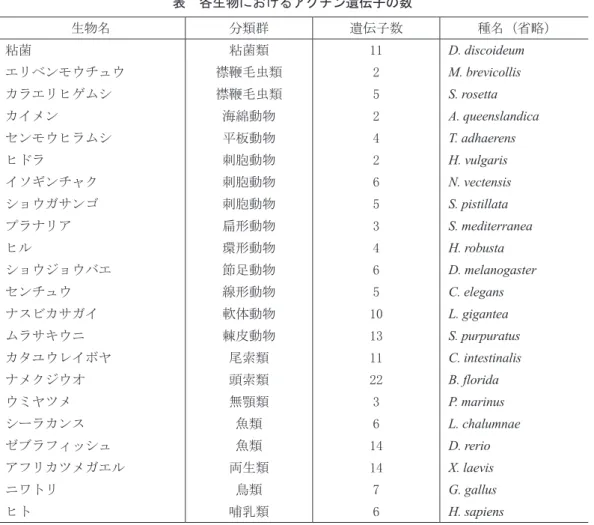
Ⅱ.アクチンの検索 アクチンの検索を行なった動物は、カイメ ン、刺胞動物のヒドラ、イソギンチャク、ショ ウガサンゴ、平板動物のセンモウヒラムシ、 環形動物のヒル、節足動物のショウジョウバ エ、線形動物のセンチュウ、頭索動物のナメ クジウオ、無顎類のウミヤツメ、魚類のゼブ ラフィッシュとシーラカンス、哺乳類のヒト などであり、表に示した。なお、刺胞動物は、 花虫綱、鉢虫綱、箱虫綱、ヒドロ虫綱の4つ の綱に分類されている。ヒドラはヒドロ虫綱、 イソギンチャクとショウガサンゴは花虫綱に 属する。扁形動物のプラナリアは、SmedGD 2.03)のデータベースを利用した。また、表に 示してある通り、関連の生物で襟鞭毛虫、粘 菌などのアクチンも検索した。 各生物のアクチンはUnitProt4)のデータベー スから検索し、ドメイン構造はInterPro5)およ びSMART6)によって確認した。さらに、各生 物のアクチンの相同検索により、類似のアク チンを広く検索した。プラナリアの検索には SmedGD 2.0を使用した。 タンパク質の相同検索には、UnitProt上で BLASTを使用した。アミノ酸配列の類似度 はBLASTの数値を参考にした。多重配列は UnitProt上ではClustal Omegaを使用し、また 単 独 で はMac上 でClustal X7)を 使 用 し た。 多 重配列より系統樹を作成し、類縁関係の推 定に利用した。系統樹の表示には、Mac版の Njplot8)とFigTree9)を使用した。 Ⅲ.各種生物のアクチン 動物および関連の生物のアクチンを検索 し、その数について表にまとめた。進化的に 新しい生物ほど遺伝子数が多くなっていると は言えない。例えば、ナメクジウオの場合、 アクチンの数は22と著しく多い。また、ゼブ ラフィッシュや両生類でも多いことが認めら れた。一方、同じ魚類でもシーラカンスは少 なく、四足類と同じような数であった。表に 載せていないが、同じ四足類のアメリカアリ ゲーター(爬虫類)では、アクチンは6種類 であった。 哺乳類では、6種類のアクチンのタイプが 知られている。2種類は細胞質骨格由来のも の(βとγ)、平滑筋に存在するアクチンが 2種類(腸管と動脈由来)、残りは骨格筋に存 在するものと心筋に存在するものである。す なわち、哺乳類では、細胞質タイプが2種類 で筋肉タイプが4種類存在する。 Ⅳ.アクチンの分子進化 動物と近縁の生物として、襟鞭毛虫、酵母、 粘菌、そして動物のカイメンを含めてアクチ ンの系統樹を作成した(図1)。襟鞭毛虫は単 細胞生物であるが、この襟鞭毛虫が複数集合 して多細胞生物の動物が進化してきたと言わ れている。酵母、粘菌は真核生物の中ではよ り動物に近いものとされている。系統樹では、 アクチンは3つのグループに分けることがで きた。Ⅰのグループは、エリベンモウチュウ とカラエリヒゲムシの一部である。Ⅱのグ ループは酵母(S.cerevisiae)と粘菌の一部の グループであった。Ⅲのグループには、カラ エリヒゲムシの一部と粘菌の一部とカイメン が存在した。この系統樹により、襟鞭毛虫類 の中で、酵母や粘菌の祖先となる生物から動 物が発生したことが推定される。 カイメンは襟鞭毛虫類よりも酵母や粘菌に 近いことがわかったので、酵母や粘菌と動物 のアクチンの系統樹を作成した(図2)。Ⅰの グループは平板動物であるセンモウヒラムシ のみであった。Ⅱのグループは刺胞動物、環 形動物のヒル、棘皮動物のウニ、尾索類のホ ヤ、頭索類のナメクジウオ、両生類のアフリ カツメガエル、哺乳類のヒトを含む。Ⅲのグ ループには、棘皮動物のヒル、両生類のアフ リカツメガエル、哺乳類のヒトが存在する
3) Robb et al.(2015) http://smedgd.stowers.org/ 4) http://www.uniprot.org/
5) Mitchell, et al.(2015)
6) Letunic et.al. (2012) https://metazome.jgi.doe.gov/pz/
7) Larkin et.al.(2007) 8) Perrière & Gouy (1996)
が、これらは筋肉タイプのアクチンである。 さらにセンチュウ、軟体動物のナスビカサ ガイ、ショウジョウバエのアクチンも含まれ る。ショウジョウバエの場合は、細胞質タイ プ(P10987、P02572)と筋肉タイプ(P10981、 P53501)の2種類がある。このグループには、 カ イ コ(B. mori)と線形動物の旋毛虫(T. murrelli)も存在する。Ⅳのグループは粘菌と 刺胞動物の一部とカイメンが存在する。この ことから、Ⅳのグループが進化的には古いグ ループと言える。また、カイメンのアクチン は他の動物とは異なった進化を辿ったと言え る。 動物は側生動物と真正後生動物と分類さ れ、側生動物として海綿動物と平板動物が含 まれる。さらに、真正後生動物は放射相称動 物と左右相称動物とに分類される。放射相称 動物には刺胞動物が含まれる。左右相称動物 の方は、前口動物と後口動物とに分けられる。 後口動物からは脊椎動物が進化してきた。ま ず、前口動物について、アクチンの系統樹 を作成し、図3に表示した。この系統樹では、 アクチンは3つのグループに分類される。Ⅰ のグループには海綿動物、軟体動物、刺胞動 物が含まれる。Ⅱのグループは平板動物、刺 胞動物、扁形動物、環形動物、節足動物、線 形動物が含まれる。Ⅲのグループには、刺胞 動物と軟体動物が含まれる。前口動物の中で 特異なのは軟体動物のナスビカサガイで、遺 伝子の数が多く多様化しており、3つのグルー プにそれぞれ属している。 次に、後口動物のアクチンの系統樹を作成 し、図4に示した。ここでは、4つのグループ に分けることができる。Ⅰのグループには棘 皮動物、頭索類などが存在する。Ⅱのグルー プをみると、ヒトのアクチンP68133は骨格筋 のアクチンである。P68032は心筋のアクチン である。P63267とP62736は平滑筋のアクチン である。C3XZ13はナメクジウオの脊索に存 在するアクチンである。W4XTP5はムラサキ ウニが持つ唯一の筋肉アクチンである。この グループは筋肉タイプアクチンのグループと 言える。このグループでは、頭索類のナメク ジウオ(B. floridae)のアクチンの多様化が顕 著である。 グループⅢの中の2つのサブグループのう ちの1つにヒトのアクチンのP60709とP3261と が入っており、これらは細胞質タイプアクチ ンである。また、このサブグループは無顎類 と脊椎動物のアクチンが含まれており、もう 一つのアクチンのグループと明らかに異なる 動物のアクチンである。このグループは細胞 質タイプアクチンといえる。また、このグルー プでは、棘皮動物のウニ(S. purpuratus)の アクチン分子の多様化が注目される。Ⅳのグ ループには、棘皮動物、尾索類、無顎類、魚 類が含まれる。後口動物の初期の動物が含ま れており、進化的に古いアクチンのグループ と推定される。 Ⅴ.まとめ アクチンは広い範囲の生物に分布してい る。特に、真核生物では進化上保守的で類似 性は80%もある。しかし、進化的にもっとも 古いカイメンのアクチンは、他の動物とは異 なっている。カイメンには収縮機能を持つ細 胞が存在するが10)、この細胞に有するアクチ ンは筋肉タイプのアクチンとは異なると思わ れる。このアクチンは、粘菌やカラエリヒゲ ムシが持っているアクチンに似ていると言え る。筋肉タイプのアクチンの出現時期は、1 つは前口動物と後口動物が分かれたころでこ のタイプは脊椎動物の筋肉タイプのアクチン となったと推定される。他の1つは節足動物 の筋肉タイプで、出現の時期は前者よりも新 しいと思われる。このように、筋肉タイプの アクチンは動物の進化の過程で何回か出現し たと考えられる。刺胞動物の横紋筋は左右相 称動物のものとは独立に出現したと言われて おり、同様の現象と思われる11)。 10) 大堀兼男(2018) 11) Steinmetz et. al (2012)
アクチンは筋肉組織の主要なタンパク質で はあるが、筋肉組織にはミオシンを含め多様 なタンパク質が構成成分として存在する。し たがって、アクチンの分子進化だけで筋肉組 織の進化を説明することは困難である。筋肉 は脊椎動物と無脊椎動物で大きく異なる。脊 椎動物の場合は大きく2種類、横紋筋と平滑 筋という筋肉がある。横紋筋はさらに骨格筋 と心筋がある。一方、無脊椎動物では横紋 筋、斜紋筋、平滑筋の3種類がある。OOtaと Saitou12)は、アクチンだけでなく、トロポニ ンC、ミオシン必須短鎖、ミオシン制御短鎖、 ミオシン重鎖の系統樹を重ね合わせて、筋肉 組織の進化を推定した。これによると、節足 動物の非筋、脊椎動物の非筋、脊椎動物の平 滑筋が1つのグループを構成している。他の グループは、節足動物の横紋筋、尾索類の平 滑筋、脊椎動物の骨格筋と心筋を含むグルー プである。今回の結果は単一の分子の進化を 扱ったものであり、OOtaとSaitouとは異なる 方法に基づくものであるが、その違いについ ては今後の検討が必要である。 Gunningら13)は、脊椎動物の筋肉タイプのア クチンを2つのグループに分けた。1つは骨格 筋と心臓平滑筋のグループ、他の1つのグルー プは心筋と平滑筋である。また、トロポミオ シンが菌類と後生動物とのアクチン繊維の機 能の多様化につながったと推測している。こ のように筋肉組織の進化については、アクチ ンだけでなく他の主要なタンパク質の進化の 検討も必要であると考えられる。 文献 ウィルマー、P.『無脊椎動物の進化』蒼樹書房、 1990年 大堀兼男『免疫グロブリンスーパーファミ リーと動物の進化』 環境と経営、第24巻第 2号、2018年101〜106ページ
Gunning, P. W., Ghoshdastider, U., Whitaker, S., Popp, D., & Robinson, R. C. (2015). The
evolution of compositionally and functionally distinct actin filaments. Journal of Cell Science, 128(11), 2009 - 2019.
Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA, McWilliam H, Valentin F, Wallace IM, Wilm A, Lopez R, Thompson JD, Gibson TJ, Higgins DG. (2007). Clustal W and Clustal X version 2.0. Bioinformatics, 23, 2947-2948.
Letunic, I., Doerks, T. and Bork, P. “SMART 7: recent updates to the protein domain annotation resource”, Nucleic Acids Res. Vol. 40, 2012, pp. D302-D305
Mitchell, A., Chang, H.-Y., Daugherty, L., Fraser, M., Hunter, S., Lopez, R., McAnulla, C., McMenamin, C., Nuka, G., Pesseat, S., Sangrador-Vegas, A., Scheremetjew, M., Rato, C., Yong, S.-Y., Bateman, A., Punta, M.,Attwood, T. K., Sigrist, C. J.A., Redaschi, N., Rivoire, C., Xenarios, I., Kahn, D., Guyot, D., Bork, P., Letunic, I., Gough, J., Oates, M., Haft, D., Huang, H., Natale, D. A., Wu, C., H., Orengo, C., Sillitoe, I., Mi, H., Thomas, P.D. and Finn, R. D. “The InterPro protein families database”, Nucleic Acids Res., Vol. 43, 2015, pp. D213 - 221
OOta, S. and Saitou, N. (1999) Phylogenetic Relationship of Muscle Tissues Deduced from Superimposition of Gene Trees. Mol.Biol.Evol. 16(6):856-867
Perrière, G. and Gouy, M. (1996) WWW-Query: An on-line retrieval system for biological sequence banks. Biochimie, 78, 364-369. Pollard, T. D., & Cooper, J. A. (2009). Actin, a
Central Player in Cell Shape and Movement. Science 27:326(5957):1208-1212
Pollard, T. D., & Goldman, R. D. (2018). Overview of the Cytoskeleton from an Evolutionary Perspective. Cold Spring Harbor Perspectives in Biology, 10(7). 1-7
12) OOta & Saitou(1999) 13) Gunning et.al.(2015)
Robb, S.M., Gotting K., Ross E., and Sánchez Alvarado A., “ SmedGD 2.0: The Schmidtea mediterranea genome database., Genesis Vol. 53, 2015 , pp.535-546
Ruiz-Gómez, M., Coutts, N., Price, A., Taylor, M. V., and Bate, M. “Drosophila Dumbfounded: A Myoblast Attractant Essential for Fusion”, Cell, Vol. 102, Issue 2, 2000, pp. 189-198
Schmidt-Rhaesa, A., The Evolution of Organ Systems, New York, Oxford University Press, 2007
Srinivas, B. P., Woo, J., Wan, Y. L., and Roy, S. “A conserved molecular pathway mediates myoblast fusion in insects and vertebrates”, Nature Genetics, Vol.39, 2007, pp.781-786. Steinmetz, P. R. H., Kraus, J. E. M., Larroux,
C., Hammel, J. U., Amon-Hassenzahl, A., Houliston, E., Wörheide, G., Nickel, M., Degnan, B. M., Technau, U. “Independent evolution of striated muscles in cnidarians and bilaterians”, Nature Vol. 487, 2012, pp. 231–234 Wu, C.H., Apweiler, R., Bairoch, A., Natale,
D.A., Barker, W.C., Boeckmann, B., Ferro, S., Gasteiger, E., Huang, H., Lopez, R., Magrane, M., Martin, M.J., Mazumder, R., O'Donovan, C., Redaschi, N. and Suzek, B. “The Universal Protein Resource (UniProt): an expanding universe of protein information”, Nucleic Acids Res., Vol. 34, 2006, pp. D187 - 191
図1 襟鞭毛虫類、粘菌などのアクチンの系統樹 0. 05 P0 78 30 _D .d is co id eu m Q 54 G X7 _D .d is co id eu m F2 U C K1 _S .ro se tta F2 U 1I 1_ S. ro se tta P0 78 29 _D .d is co id eu m F2 U C J6 _S .ro se tta A9 V4 89 _M .b re vi co llis Q 54 H F1 _D .d is co id eu m Q 95 U U 0_ M .b re vi co llis Q 54 H E9 _D .d is co id eu m F2 U 0P 3_ S. ro se tta P0 78 28 _D .d is co id eu m Q 54 H F0 _D .d is co id eu m A0 A1 X7 VV X1 _A .q ue en sl an di ca P6 00 10 _S .c er ev is ia e Q 55 3U 6_ D .d is co id eu m Q 55 4S 6_ D .d is co id eu m Q 55 C U2 _D .d is co id eu m Q 55 EU 6_ D .d is co id eu m Q 54 L5 4_ D .d is co id eu m A0 A1 X7 VG Y7 _A .q ue en sl an di ca F2 U C J7 _S .ro se tta Ⅰ Ⅱ Ⅲ
図2 動物のアクチンの系統樹 0. 05 A0 A2 B4 S2 83 _S .p is till at a A7 SC N 8_ N .v ec te ns is P1 09 87 _D .m el an og as te r P0 78 30 _D .d is co id eu m Q 54 L5 4_ D .d is co id eu m W 4X TP 5_ S. pu rp ur at us A7 R U 31 _N .v ec te ns is A0 A2 B4 S2 02 _S t.p is til la ta P6 32 61 _H .s ap ie ns V4 BI V6 _L .g ig an te a T2 M EG 4_ H .v ul ga ris A7 S0 V7 _N .v ec te ns is B3 S5 E4 _T .a dh ae re ns Q 93 13 1_ B. flo rid ae P0 D M 41 _C .e le ga ns Q 55 4S 6_ D .d is co id eu m P1 09 81 _D .m el an og as te r P1 71 26 _H .v ul ga ris B3 R X7 7_ T. ad ha er en s A2 SY 09 _C .in te st in al is V3 Z1 37 _L .g ig an te a P0 25 72 _D .m el an og as te r T1 FQ Q 4_ H .ro bu st a A7 SE A7 _N .v ec te ns is A0 A1 X7 VG Y7 _A .q ue en sl an di ca P1 09 84 _C .e le ga ns A0 A1 X7 VV X1 _A .q ue en sl an di ca Q 55 EU 6_ D .d is co id eu m A7 SE B1 _N .v ec te ns is Q 54 G X7 _D .d is co id eu m T1 G 9A 8_ H .ro bu st a Q 55 C U 2_ D .d is co id eu m P5 34 72 _S .p ur pu ra tu s Q 55 3U 6_ D .d is co id eu m Q 54 H F0 _D .d is co id eu m A7 R U 30 _N .v ec te ns is A0 A0 V0 U H V8 _T .m ur re lli A0 A2 B4 R 4C 5_ S. pi st illa ta P6 81 33 _H .s ap ie ns A0 A2 B4 ST 10 _S .p is till at a P0 48 29 _B .m or i P6 00 10 _S .c er ev is ia e P6 07 09 _H .s ap ie ns P1 09 86 _C .e le ga ns P6 27 36 _H .s ap ie ns Q 7Y TV 5_ T. ad ha er en s O 93 40 0_ X. la ev is B3 R KR 4_ T. ad ha er en s P0 78 29 _D .d is co id eu m P1 09 95 _X .la ev is Q 54 H E9 _D .d is co id eu m Q 54 H F1 _D .d is co id eu m P5 35 01 _D .m el an og as te r P0 78 28 _D .d is co id eu m Ⅰ Ⅱ Ⅲ Ⅳ
図3 前口動物のアクチンの系統樹 0. 04 V4 BI V6 _L .g ig an te a V4 B6 R 7_ L. gi ga nt ea V3 ZN H 0_ L. gi ga nt ea P1 09 84 _C .e le ga ns SM U 15 02 25 32 _S .m ed ite rra ne a A7 R U 30 _N .v ec te ns is T1 G 9A 8_ H .ro bu st a A7 S0 V7 _N .v ec te ns is V4 BH 33 _L .g ig an te a V3 Z1 37 _L .g ig an te a A0 A2 B4 R 4C 5_ S. pi st illa ta 0D M 41 _C .e le ga ns V4 BC 69 _L .g ig an te a A7 R U 31 _N .v ec te ns is P1 09 81 _D .m el an og as te r A0 A1 X7 VG Y7 _A .q ue en sl an di ca A7 SC N 8_ N .v ec te ns is SM U 15 03 16 10 _S .m ed ite rra ne a A0 A2 B4 ST 10 _S .p is til la ta P1 09 86 _C .e le ga ns A0 A2 B4 S2 83 _S .p is til la ta O 45 81 5_ C .e le ga ns P5 35 01 _D .m el an og as te r B3 S5 E4 _T .a dh ae re ns T1FQ Q 4_ H .ro bu st a P0 25 74 _D .m el an og as te r Q 7Y TV 5_ T. ad ha er en s T1 FM E7 _H .ro bu st a V4 B4 D 3_ L. gi ga nt ea A7 SE B1 _N .v ec te ns is P1 71 26 _H .v ul ga ris P0 25 72 _D .m el an og as te r A0 A2 B4 S2 02 _S .p is til la ta V4 BV 77 _L .g ig an te a T2 M EG 4_ H .v ul ga ris P8 39 67 _D .m el an og as te r V3 ZS W 2_ L. gi ga nt ea V4 B4 76 _L .g ig an te a P1 09 87 _D .m el an og as te r B3 R X7 7_ T. ad ha er en s A7 SE A7 _N .v ec te ns is B3 R KR 4_ T. ad ha er en s SM U 15 01 76 01 _S .m ed ite rra ne a T1 FM D 2_ H .ro bu st a A0 A1 X7 VV X1 _A .q ue en sl an di ca Ⅰ Ⅱ Ⅲ
図4 後口動物のアクチンの系統樹 0. 06 C 3Z M R 5_ B. flo rid ae W 4Y 96 9_ S. pu rp ur at us W 4Z 0B 1_ S. pu rp ur at us F1 R C B6 _D .re rio P1 09 95 _X .la ev is C 3X YD 5_ B. flo rid ae H 3A 44 0_ L. ch al um na e Q 07 90 3_ S. pu rp ur at us P6 81 33 _H .s ap ie ns F6 R 0S 4_ C .in te st in al is C 3X YD 3_ B. flo rid ae C 3X YC 8_ B. flo rid ae C 3X YD 2_ B. flo rid ae P5 35 05 _X .la ev is W 4X TP 5_ S. pu rp ur at us W 4Y 1A 3_ S. pu rp ur at us P6 07 09 _H .s ap ie ns P6 81 39 _G .g al lu s P5 34 73 _S .p ur pu ra tu s C 3Z H N 2_ B. flo rid ae W 4Y KQ 1_ S. pu rp ur at us W 4Y 15 7_ S. pu rp ur at us O 93 40 0_ X. la ev is C 3Z H N 4_ B. flo rid ae P6 90 05 _S .p ur pu ra tu s F6 Q BJ 1_ C .in te st in al is H 3B G S4 _L .c ha lu m na e F6 Q 66 2_ C .in te st in al is A2 BD B0 _X .la ev is W 4Y H N 2_ S. pu rp ur at us S4 R H 85 _P .m ar in us P6 27 36 _H .s ap ie ns Q 93 13 2_ B. flo rid ae Q 7Z VF 9_ D .re rio A0 A1 D 5N V1 7_ G .g al lu s C 3X YC 6_ B. flo rid ae P6 07 06 _G .g al lu s P6 80 32 _H .s ap ie ns S4 S0 S5 _P .m ar in us P1 84 99 _S .p ur pu ra tu s A0 A1 X7 VG Y7 _A .q ue en sl an di ca C 3Z 4K 3_ B. flo rid ae P5 34 72 _S .p ur pu ra tu s C 3X YC 5_ B. flo rid ae F6 SC P4 _C .in te st in al is A0 A0 C 3S G 04 _S .p ur pu ra tu s C 3X YC 2_ B. flo rid ae F6 SZ Q 0_ C .in te st in al is P6 32 61 _H .s ap ie ns C 3Y P5 8_ B. flo rid ae Q 93 13 1_ B. flo rid ae C 3X YC 7_ B. flo rid ae F6 SC 57 _C .in te st in al is C 3X YD 4_ B. flo rid ae C 3Z TX 6_ B. flo rid ae A0 A1 W 2W EN 5_ C .in te st in al is H 3A 7F 0_ L. ch al um na e F6 T8 C 7_ C .in te st in al is Q 5Z M Q 2_ G .g al lu s A0 A1 51 M BG 8_ A. m is si ss ip pi en si s H 3A VL 6_ L. ch al um na e H 3A 4B 1_ L. ch al um na e C 3Y KS 6_ B. flo rid ae Q 6I P2 2_ X. la ev is C 3X Z1 3_ B. flo rid ae H 3A U T1 _L .c ha lu m na e H 3A VL 5_ L. ch al um na e Q 7Z VI 7_ D .re rio P6 32 67 _H .s ap ie ns P5 34 74 _S .p ur pu ra tu s C 3Z M R 2_ B. flo rid ae H 2Z W 67 _L .c ha lu m na e Q 7Z W R 7_ X. la ev is F6 R 3E 1_ C .in te st in al is F6 SD C 9_ C .in te st in al is Q 6D H S1 _D .re rio S4 S1 42 _P .m ar in us C 3Z M 16 _B .fl or id ae A0 A1 X7 VV X1 _A .q ue en sl an di ca H 3B BJ 9_ L. ch al um na e C 3X YC 4_ B. flo rid ae F1 Q U N 8_ D .re rio A0 A0 R 4I EZ 2_ D .re rio A2 SY 09 _C .in te st in al is Ⅰ Ⅱ Ⅲ Ⅳ
表 各生物におけるアクチン遺伝子の数 生物名 分類群 遺伝子数 種名(省略) 粘菌 粘菌類 11 D. discoideum エリベンモウチュウ 襟鞭毛虫類 2 M. brevicollis カラエリヒゲムシ 襟鞭毛虫類 5 S. rosetta カイメン 海綿動物 2 A. queenslandica センモウヒラムシ 平板動物 4 T. adhaerens ヒドラ 刺胞動物 2 H. vulgaris イソギンチャク 刺胞動物 6 N. vectensis ショウガサンゴ 刺胞動物 5 S. pistillata プラナリア 扁形動物 3 S. mediterranea ヒル 環形動物 4 H. robusta ショウジョウバエ 節足動物 6 D. melanogaster センチュウ 線形動物 5 C. elegans ナスビカサガイ 軟体動物 10 L. gigantea ムラサキウニ 棘皮動物 13 S. purpuratus カタユウレイボヤ 尾索類 11 C. intestinalis ナメクジウオ 頭索類 22 B. florida ウミヤツメ 無顎類 3 P. marinus シーラカンス 魚類 6 L. chalumnae ゼブラフィッシュ 魚類 14 D. rerio アフリカツメガエル 両生類 14 X. laevis ニワトリ 鳥類 7 G. gallus ヒト 哺乳類 6 H. sapiens