Changes in amino acids composition accompanied with senescence of detached leaves
6
0
0
全文
(2) 88. proteolytic amino acids accumulation and protein turnover in senescent leaves are still not clear (Beevers, 1976). Therefore, it is interesting that which amino acids are used as nitrogen carriers in the nitrogen recovering process and whether contents of specific ammo acids increase or not in the process. The author examined the changes in composition of seventeen soluble amino acids, which are commonly found in proteins, in the process of bean plant leaf senescence.. Materials and Methods. French dwarf beans (Phaseolus vulgaris L. cv. Shin-edogawa) purchased locally were grown in a green house. Four weeks after sowing, the first trifohated leaves of bean plants were fully expanded and excised at their axils, and after recutting their periols in water the leaves were dipped their petioles in. distilled water. The leaves were placed in the dark room maintained at 26土1 ℃ during the experimental period.. Ca. 2 grams of the leaves were homogenized in ethanol (80^v/v) at an interval of two days. The homogenates were boiled for five minutes to extract soluble amino acids with a reflux condencer. Then the boiled homogenates were filtrated and additional extraction was performed on the residues for three times in the same manner. The combined filtrates were assayed their contents of ammo acids as ninhydrin positive substances, and applied to a column of H form Amberlite IR-120A equilibriated with 80^ ethanol (one ml resine used for 0.1 mmole leucine equivalence). The absorbed amino acids were eluted with 2n NH40H using ten times volume of resine, and after concentration in vacuo analyzed with automatic amino acid analyzer (KLA'5, Hitachi Ltd., Tokyo, Japan).. Results and Discussions. After four days of incubation excised leaves became etiolated as a result of senescence, and even abscission layer was formed on each petiolule. Although excised leaves did not dehydrate during the experimental periods, losses in the weight of leaves could be imputed to neither dehydration nor respiration of leaves. Therefore, the contents of amino acids in leaves were expressed on the basis of mmole per thousand leaves. Amino acid analysis was carried on the seventeen amino acids commonly found in proteins. These amino acids could be classified into five families on the origin of their C skeletons, that is, a serine family whose C skeleton is derived from 3-phospoglyceric acid in Calvin cycle, a pyruvate family whose C skeleton is derived from pyruvate in glycolysis path way, a shikimic acid family whose C skeleton is derived from shikimic acid, and an aspartate family and a glutamate family whose C skeletons are derived from oxalacetate and ot-ketoglutarate.
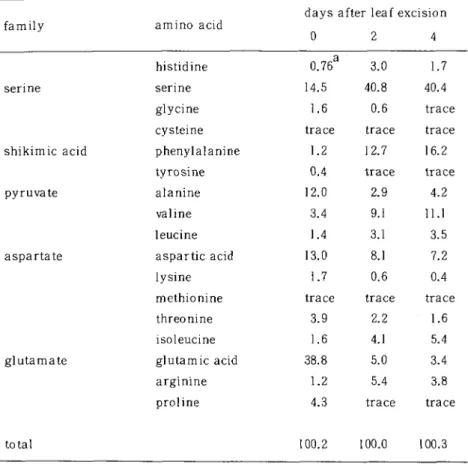
(3) Changes in amino acids composition accompanied with senescence of detached leaves. in the citric acid cycle, respectively. Sixteen ammo acids belong to these five families and histidine were assayed. In these five families, two of the glutamate family except argimne did not increased in their contents during the senescence (Fig- 4). In the other four families, the most amino acids were increased their contents in leaves, except glycine and cysteine of the serine family (Fig. 1), tyrosine of the shikimic acid family (Fig. 2), and methionine in the aspartate family (Fig. 3). Especially all of the pyruvic acid family were increased in their contents (Fig. 2). Serine, however, became to form forty per cent or more of the total amino acids detected, while valine and phenylalanine reached to form up to ten per cent during the senescence (Table 1). After four days of incubation, the contents of serine increased 76 times (Fig. 1) and those of valine and phenylalanine increased 34 times and 119 times, respectively (Fig. 2). In the rest amino acids of which contents were increased during the senescence, the amounts were increased from 1.3 times (lysine) to 31 times (isoleucine) of their initial contents. The author and a co-worker previously reported the increase in the contents of tryptophan in senescent bean leaves (Atsumi and Hayashi, 1979), but tryptophan did form less than 3 per cent of total amino acids and the contents were increase ca. 34 times of the initial amounts. Leaf segments of oat was reported to accumulate serine and not to accumulate glutamine during their senescence in the dark (Malik, 1982). Also the accumulation of sei;ine in the senescent bean leaves (Fig. 1 and Table 1) might be due to the incubation in the dark and the absence of photorespiration as same as. in oat (Malik, 1982) and wheat (Kar and Feierabend, 1984). On the other hand, glutamine and glutamic acid synthase activities were reported to decline rapidly in the dark (Kar and Feierabend, 1984). In the present works glutamme and asparagine were not analyzed, because convensional ammo acid analyzers such as used in this work could not sharply separate these two basic amino acids from threonine. However, the contents of threonine were 149 /(moles per 1000 leaves and occupied only 1.6 per cent of total amino acids at days four (Fig. 3 and Table 1). And the chromatograms of threonine had no shoulders on its peak. Accordingly it might be approved that there were few amounts of glutamine and asparagine in senescent been leaves as a result of the senescence in the dark. Serine was also found to be the most active amino acid to promote senescence of oat leaves, while alanine, glycine, cystein and phenylalanine had semilar but. less effect (Shibaoka and Thimann, 1970, Martin and Thimann, 1972). In the excised leaves of bean plants accumulated serine and phenylalanine might accelerate the senescence of the leaves incubated in the dark. However, it seemed to be adequent that the lysis of chloroplasts in the dark evoked leaf senescence and in the process of senescence serine was accumulated as a products of proteolysis and/or the other metabolisms..
(4) 90. 8 6 ′ -. saADairOLJadb¥o∈∈. 2. U. days after excision. Fig. 1 Time course of changes in the contents of serine (-「▲i-), cysteine ) andglycine ( ○ ) belong to the serine family amino acids during the senescence of excised bean leaves. days after excision Fig. 2 Time course of changes in the contents of phenylalanine (一・-) and tyrosine ( △ ) of the shikimic acid. placed in the dark.. family, and alanine (-・▲・」, valine (-O-) and leucine (・蝣一--) of the. pyruvic acid family during the senescence of excised bean leaves placed in the dark.. 5. 0 0. 1 0. 3. 2. CO. O O o O s9AD3│OLJ9d8│0∈∈. /. S8AD8│﹂OLJ9d910UIUJ. 0. U. days after excision. Fig. 3 Time course of changes in the. Fig. 4 Time course of changes m the. contents of aspartic acid (」トー),. contents of glutamic年cid (-Al一),. lysineト・◆--), methionine ( △ ),. argmme C・書・⇒ and proline (・-・題・-・-) of. threonine. (-lm・-蝣). and. insoleucine. glutamate familyduringthe senescence of. 〔-{ゝ-〕 of the aspartate family during the senescence of excised bean leaves. excised bean leaves placed in the dark.. placed in the dark.. histidine C--0--) was also shown.. In addition with the glutamate family,.
(5) Changes in amino acids composition accompanied with senescence of detached leaves 91. Table 1 Changes in thecomposision ofamino acids in excised bean leaves during their senescence. days after leaf excision. amino acid. family. 0. histidine. 0.76 14.5 1.6 trace. cysteine. acid. phenylalanine tyrosine. pyruvate. alanine. 0.4. 1. valine. 2.0 3.4. leucine aspartate. aspartic. 1.4. acid. lysine. 13.0. 1.7. methionine. trace. threonine. glutamate. 1.2. 3.9. isoleucine. 1.6. glutamic. 38.8. acid. arginine. 1.2. proline. 4.3. QJQJ<D<V(D 。。cm。c^Uifi(N c・ ' o・ t c・ u -C O3 i_,川LOcOCO 4ttLltltt. serine glycine. shikimic. -1. QJCDCDCD 。ocD ooC--0 c0 m5 '2csjc ao ic <oo I T*+J^_^J+->+J. senne. 2. 100.2 100.0 100.3. total. Values were expressed on the basis of percentage of each amino acid to total amino acids detected.. Acknowledgement. The author makes a greatiful acknowledgement for Prof. T. Hayashi (Univ. Tokyo) to allow an opportunity to study on this theme. The author also wishes to thank Prof. T. Isobe (Tokyo Metropolitan Univ.) for the amino acid analysis, and Prof. T. Yamada (Hyogo Univ. Teacher Education) for a critical reading of this manuscript..
(6) 92. References. (1). Addicott, F. T∴ Plant hormones in the control of abscission. Biol. Rev. 45: 485-524 (1970). (2). Atsumi, S. and T. Hayashi: Examination of the pronounced increase in auxin content of senescent leaves. Plant & Cell Physiol. 20: 86ト865 (1979). (3). Beevers, L∴ Senescence. In Plant Biochemistry, pp. 771-794, Bonner, J.. and J. E. Varner, eds. New York: Academic Press (1976). (4). Kar, M. and J. Feierabend: Changes in the activities of enzymes involved in amino acid metabolism during the senescence of detached wheat leaves. Physiol. Plant. 62: 39-44 (1984). (5). Malfatti, H., J. C. Vallee, E. Perdrizet, M. Carre, and C. Martin: Acides amines et amines libres d'explants foliaires de Nicotiana tabacum cultives in vitro sur des milieux induisant la rhizogenese ou la caulogenese. ibid. 57: 492-498 (1983). (6). Malik. N. S. A∴ Senescence in detached oat leaves I. Changes in free amino. acid levels. Plant & Cell Physiol. 23: 49-57 (1982). (7). Martin, C. and K. V. Thimann: Role of protein synthesis in the senescent leaves II. The influence of amino acids on senescence. Plant Physiol. 50: 432-437 (1972). (8). Sheldrake, A. R.: The production of hormones in higher plants. Biol. Rev. 48: 509-559 (1973). (9 ). Shibaoka, H. and K. V. Thimann: Antagonism between kinetin and amino acids. Experiments on the mode of action of cytokinins. Plant Physiol. 46: 212-220 (1970). (10). Thimann, K. V∴ The senescence of leaves. In Senescence in Plants, pp-. 85-115, Thimann, K. V., ed. Florida: CRC Press (1980). (ll). Thomas, H.: Enzymes of nitrogen mobilization in detached leaves of Lolium temulentum during senescence. Planta 142: 161-169 (1978). (12). - and Stoddart, J. L.: Leaf senescence. Ann. Rev. Plant Physiol. 31: 83-111 (1980)..
(7)
図
関連したドキュメント
Standard domino tableaux have already been considered by many authors [33], [6], [34], [8], [1], but, to the best of our knowledge, the expression of the
The edges terminating in a correspond to the generators, i.e., the south-west cor- ners of the respective Ferrers diagram, whereas the edges originating in a correspond to the
In order to obtain more precise informations of b(s) and ~ , we employ Hironaka's desingularization theorem.. In this section, as its preparation, we will study the integration
We show that a discrete fixed point theorem of Eilenberg is equivalent to the restriction of the contraction principle to the class of non-Archimedean bounded metric spaces.. We
In this work we give definitions of the notions of superior limit and inferior limit of a real distribution of n variables at a point of its domain and study some properties of
When a 4-manifold has a non-zero Seiberg-Witten invariant, a Weitzenb¨ ock argument shows that it cannot admit metrics of positive scalar curvature; and as a consequence, there are
The intent of this article is to study the Kleisli extension introduced in [20] for Sup-enriched monads, and its interaction with four other types of extensions: the initial
この chart の surface braid の closure が 2-twist spun terfoil と呼ばれている 2-knot に ambient isotopic で ある.4個の white vertex をもつ minimal chart
