Ef f ect of Lys os t aphi n on Es t abl i s hment of St aphyl ococcal I nf ect i ous Foci i n Mous e Ki dney
Kei ko S
EKI,Hi t omi S
HINJI,Mi yo M
URAI,Aki ko T
AJIMA, and Shogo M
ASUDADepar tment of Micr obiology (II), The Jikei Univer sity School of Medicine Depar tment of Medical Technology, Saitama Pr efectur al Univer sity Junior College
ABSTRACT
To clarify the mechanis
ms how Staphylococcus aureus organisms proliferate and how the infectious foci are established in mouse kidney,we carried out the in vivo experiments using mice and the in vitro experiments using kidney homogenat e of mice and lysostaphin.
We demonstrated that the number of S. aureus organisms in the kidneys of mice treated with lysostaphin 1 hr after injection of the bacteria was one‑third of that without lysostaphin treatment. The major reason of the diminution seemed that the bacteria had been directly killed by lysostaphin in the bloodstream before they came to the kidney. The bacteria that could escape from lyso- staphin‑attack were thought to exist inside leukocytes or some renal cells,or the very narrow intracellular space of the kidney. We also demons trated that the leukocytes got a higher phagocytic activity according to the injection of heat‑killed bacteria. In the formation of infec- tious foci,leukocytes were suspected to play two conflicting roles,killing and protecting bacteria, and renal cells might be a hiding place for S. aureus organisms.(Jikeikai Med J 2005;52:21‑9) Key words:Staphylococcus aureus,lysostaphin,infectious foci,leukocyte,kidney cell
I
NTRODUCTION
Staphylococcus aur eus i s known t o be a pat h- ogen of dermat i t i s , endocardi t i s , and os t eomyel i t i s i n human,and has been exper i men- t al l y demons t r at ed t o pr ol i f er at e i n var i ous mur i ne t i s s ues . Our pr evi ous r epor t s s t at ed t hat about 2×10 CFU of S. aur eus Cowan I adher ed t o one ki dney of t he mous e i nt r avenous l y i nnocul at ed wi t h 1×10 CFU. Kondo et al .r epor t ed t hat a l ar ge number of t he S. aur eus or gani s ms r emai ned i n t he mous e ki dney mor e t han 7 days af t er t hei r i nt r a-
venous i nocul at i on,al t hough t hei r number s had de- cr eas ed wi t hi n a f ew days i n ot her or gans s uch as t he l i ver ,s pl een,and l ung. Why can S. aur eus pr ol i f er at e
s o s t r ongl y i n t he ki dney? One r eas on may be t hat t her e ar e onl y a l i t t l e r es i dent phagocyt es i n t he ki dney,and s o bact er i ci dal act i vi t y i n t he ki dney mi ght be l ow. On t he ot her hand s ome or gans have s ome s peci al i zed phagocyt i c cel l s s uch as t he Kupf f er cel l s i n t he l i ver ,t he al veol ar macr ophages i n t he l ung, or t he s pl enocyt es i n t he s pl een. Anot her r eas on f or t he abi l i t y of S. aur eus t o pr ol i f er at e s t r ongl y i n t he ki dney may be t he i nvol vement of s t aphyl ococcal cel l s ur f ace‑as s oci at ed s ubs t ances . The ki dney i s com- pos ed of var i ous t ypes of t i s s ues ,and t he cel l s t hat f or m t hes e t i s s ues s ecr et e s ome of t hei r ext r acel l ul ar mat r i ces . The ext r acel l ul ar mat r i x‑bi ndi ng pr ot ei ns of S. aur eus i ncl ude l ami ni n‑bi ndi ng pr ot ei n vi t r onect i n‑bi ndi ng pr ot ei n ,f i br i nogen‑bi ndi ng pr o-
Received for publication,October 19,2004
関 啓子,進士ひとみ,村井 美代,田嶌亜紀子,益田 昭吾
Mailing address:Keiko SEKI,Department of Microbiology(II),The Jikei University School of Medicine,3‑25‑8,Nishi‑Shimbashi, Minato‑ku,Tokyo 105‑8461,Japan.
E‑mail address:kseki@jikei.ac.jp
21
t ei n , and col l agen‑bi ndi ng pr ot ei n . Ot her wel l ‑known at t achment f act or s i ncl ude s t aphyl ococ- cal pr ot ei n A , cl umpi ng f act or , and f i br onect i n‑bi ndi ng pr ot ei n . However ,i t has not yet been cl ar i f i ed why or how S. aur eus can mul t i pl y i n t he ki dney t i s s ue or t he r enal cel l s of t he mous e.
I t has been r epor t ed t hat abs ces s es wer e f or med when a ki dney was s t abbed wi t h a needl e car r yi ng onl y a f ew s t aphyl ococcal cel l s ,al t hough no abs ces s was obs er ved i n t he ki dney af t er t he i nt r avenous i nj ect i on of t he s ame amount of S. aur eus ,s ugges t i ng t hat a ver y s mal l number of S. aur eus or gani s ms was s uf f i ci ent t o f or m an i nf ect i ous f ocus . Mos t of t he i nt r avenous l y‑i nocul at ed bact er i al cel l s may have been t r apped by t i s s ue‑r es i dent phagocyt es s uch as s pl enocyt es and Kupf f er cel l s ,or ki l l ed by ci r cul at i ng l eukocyt es and s er um f act or s . Thi s s pecul at i on s eemed t o have a r el at i on t o one of our f i ndi ngs : S.
aur eus or gani s ms di s appear ed i n a l i near f as hi on f r om t he bl oods t r eam dur i ng t he f i r s t 30 mi n af t er t he i nj ect i on vi a i nt r avenous r out es ,but t he di s appear - ance of t he or gani s ms f ol l owed a gent l e s l ope i n mi ce whi ch had pr evi ous l y r ecei ved a l ar ge number of heat ‑
ki l l ed bact er i a. That r es ul t s ugges t ed t hat t he mas - s i ve admi ni s t r at i on of ki l l ed or gani s ms may have i nf l uenced t he phagocyt i c f unct i on of t he l eukocyt es . I n or der t o cl ar i f y t he mechani s ms t hr ough whi ch S. aur eus or gani s ms adher e t o and pr ol i f er at e i n t he ki dney cel l s ,we des i gned an in vivo exper i ment us i ng l ys os t aphi n. Lys os t aphi n i s a pr oduct of Staphylococ- cus simulans ,and i s known t o enzymat i cal l y br eak- down pept i dogl ycan,t he maj or component of t he bact er i al cel l wal l . We demons t r at e t hat s t aph-
yl ococcal pr ol i f er at i on i n t he mous e ki dney was i nhi b- i t ed by l ys os t aphi n chal l enge,and di s cus s t he pos s i bi l - i t y t hat l eukocyt es may pl ay an i mpor t ant r ol e i n hel pi ng S. aur eus or gani s ms s ur vi ve and col oni ze by gi vi ng r ef uge t o t he bact er i a.
M
ATERIALS ANDM
ETHODSBacter ia
S. aur eus Cowan I or gani s ms wer e cul t ur ed over -
ni ght on hear t i nf us i on agar( HI A,Di f co Co.Lt d. , Det r oi t ,Mi ch. ,USA)pl at es at 37° C. Bact er i a wer e
col l ect ed,was hed t wi ce and s us pended i n phys i ol ogi - cal s al i ne. The bact er i al concent r at i on was es t i - mat ed by t ur bi di met r y at 660 nm us i ng a Juni or I I I s pect r ophot omet er Model 6/8( Per ki n‑El mer I ns t r u-
ment s ,Oak Br ook,I l l . ,USA) . Heat ‑ki l l ed bact er i a ( KB)wer e pr epar ed by boi l i ng t he l i ve ones f or 30 mi n and was hi ng wi t h s al i ne. The concent r at i on of KB was t hen adj us t ed.
Mice
Femal e I CR mi ce( 5 weeks ol d)wer e pur chas ed f r om Char l es Ri ver Japan I nc.( Yokohama,Japan) , and wer e al l owed f ood ad libitum. Leukopeni c mi ce wer e cr eat ed by i nj ect i ng 4 mg of cycl ophos phami de ( Endoxan ,Shi onogi Co.Lt d. ,Os aka,Japan)ever y ot her day f or 5 days ,af t er whi ch t he aver age number of l eukocyt es per ml of t he per i pher al bl ood of t he l eukopeni c mi ce was 3. 7×10 cel l s .
Estimation of the number of intr ar enal bacter ia Each mous e r ecei ved 0. 1 ml of bact er i al s us pen-
s i on vi a t he t ai l vei n. The i nocul at i on l evel of l i ve bact er i a was 1×10 CFU per mous e. One hour bef or e r ecei vi ng t he l i ve bact er i a,t he mous e was i nj ect ed wi t h 1×10 cel l s of KB,i f neces s ar y. Mous e was al s o i nt r avenous l y gi ven l ys os t aphi n der i ved f r om S. simulans ( Si gma‑Al dr i ch Co. ,St .Loui s ,Mo. ,USA) 1 hr af t er r ecei vi ng t he l i ve bact er i a,i f neces s ar y.
Phos phat e buf f er ed s al i ne( PBS)was us ed i ns t ead of ei t her KB or l ys os t aphi n.
The es t i mat i on of t he bact er i al number s i n t he ki dney was car r i ed out i n al mos t t he s ame way de-
s cr i bed pr evi ous l y . I n s hor t ,2 hr af t er t he i nj ec-
t i on of l i ve bact er i a,bot h ki dneys wer e as ept i cal l y
r emoved f r om each mous e under et her anes t hes i a,and
homogeni zed wi t h a s t er i l e mor t ar and pes t l e i n 2 ml
of nut r i ent br ot h. Appr opr i at el y di l ut ed homogenat e
was s pr ead on a nut r i ent agar pl at e( Di f co) ,and i n-
cubat ed at 37° C over ni ght f or col ony count i ng. The
number of bact er i a per ki dney was cal cul at ed.
Monitor ing bacter ia in kidney homogenate mixed with lysostaphin
Bot h ki dneys of t he mous e gi ven l i ve bact er i a 1 hr ear l i er wer e homogeni zed i n 2 ml of nut r i ent br ot h
( Di f co) . The homogenat e was mi xed wi t h 1. 1 uni t s / ml of l ys os t aphi n i n a t es t t ube,and i ncubat ed at 37° C.
The bact er i al number s wer e count ed af t er 0,1 and 3 hr of t he i ncubat i on t i me by col ony count i ng met hod.
Phagocytic plaque method
Changes i n t he phagocyt i c act i vi t i es of l eukocyt es af t er t he i nj ect i on of KB wer e exami ned us i ng t he phagocyt i c pl aque met hod devel oped by Seki et al .,
wi t h s l i ght modi f i cat i on. I n s hor t ,a 0. 8‑ml hepar i n- i zed bl ood s ampl e was put i nt o a pl as t i c di s h cover ed wi t h an S. aur eus ‑t hi n l ayer . Af t er 1 hr of i ncubat i on at 37° C,t he di s h was was hed t wi ce wi t h phys i ol ogi cal s al i ne,dr i ed,f i xed wi t h met hanol and s t ai ned wi t h Gi ems aʼ s s ol ut i on. Phagocyt i c f unct i on was expr es -
s ed i n t er ms of t he number of phagocyt i c pl aques ( pl aques )whi ch wer e empt y s paces f or med by l euko- cyt es af t er i nges t i ng t he bact er i al l ayer . I mage ana- l ys es wer e car r i ed out us i ng t he FRM Tool ‑Ki t com- put er pr ogr am ( ver 2. 1,Phot or on Co. ,Tokyo,Japan) wi t h a l i ght mi cr os cope under a 10× obj ect i ve l ens . I n t hi s met hod,t he s i ze was r epr es ent ed as t he num- ber s of pi xel s . The s um of t he ar eas of pl aques wi t hi n a cer t ai n mi cr os copi c f i el d was expr es s ed as t he“t ot al ar ea”,and t he mean was bas ed on t he val ues obt ai ned f r om f i ve l ocat i ons per di s h. The s i ze of pl aque was s epar at el y meas ur ed about 100 pl aques ,each of whi ch r epr es ent ed t he ar ea cover ed by t he movement of one l eukocyt e dur i ng phagocyt os i s .
Deter mination of the types of neutr ophils
A s mear was pr epar ed f r om t he bl ood of each mous e, f i xed wi t h met hanol , and s t ai ned wi t h Gi ems aʼ s s ol ut i on. The per cent ages of bot h s egment - ed and s t ab cel l s wer e cal cul at ed;and t he number of l eukocyt es was count ed wi t h a hemocyt omet er . Then,bot h s egment ed and s t ab cel l number s per ml of bl ood wer e det er mi ned.
Statistics
Dat a wer e anal yzed f or s t at i s t i cal s i gni f i cance by St udent ʼ s t t es t f or each pai r ed s ampl e. P val ues l es s t han 0. 05 wer e r egar ded as s i gni f i cant .
R
ESULTS
Inhibition of staphylococcal adhesion to the kidney by lysostaphin
At f i r s t ,t he appr opr i at e concent r at i on of l ys o-
s t aphi n t o admi ni s t er t o mi ce f or t he f ol l owi ng exper i - ment s was det er mi ned. Mi ce r ecei ved var i ous con- cent r at i ons of l ys os t aphi n 1 hr af t er t he i nj ect i on of l i ve bact er i a,and anot her hour l at er ,bot h ki dneys wer e r emoved and t he number of bact er i a was count - ed t o eval uat e t he act i on of l ys os t aphi n. As s hown i n Fi g.1,t he number of S. aur eus or gani s ms i n t he ki dney decr eas ed wi t h t he i nj ect i on of l ys os t aphi n.
The degr ee of i nhi bi t i on depended on t he dos e of l ys os t aphi n wi t hi n t he r ange of 0 t o 0. 88 uni t s per mous e. Ther e was not a s i gni f i cant di f f er ence bet ween t he number of bact er i a r ecover ed f r om mi ce i nj ect ed wi t h 0. 88 and 1. 76 uni t s of l ys os t aphi n.
Accor di ng t o t hi s r es ul t ,1. 76 uni t s of l ys os t aphi n per mous e wer e us ed about t he f ol l owi ng exper i ment s .
A l ar ge amount of l i ve bact er i a wer e r ecover ed f r om t he ki dneys of mi ce whi ch had r ecei ved KB i n advance,and t hi s r es ul t was i ndependent of l ys o-
Fig.1. Effect of lysostaphin on the staphylococcal prolif- eration in the kidney. One hour after the injec- tion of live S. aureus organisms,various concen- trations of lysostaphin were intravenously inject- ed into mice. Details are described in MATERIALS AND METHODS. Values ar e mean±SD (n=5).
s t aphi n t r eat ment( Fi g.2) . Wi t hout t he pr e‑i nj ect i on of KB,s mal l er number of bact er i a was f ound i n t he ki dneys of l ys os t aphi n‑i nj ect ed mi ce t han t hos e i n t he cont r ol mi ce. The l evel i n l ys os t aphi n‑i nj ect ed mi ce was about one‑t hi r d of t hat i n t he cont r ol .
Action of lysostaphin on the bacter ia in kidney homogenate
Thi s l ys os t aphi n concent r at i on us ed i n t hi s exper -
i ment ,1. 1 uni t s /ml ,was det er mi ned on t he bas i s of t he quant i t y i n t he bl oods t r eam of a mous e i nt r avenous l y gi ven 1. 76 uni t s . The bact er i al number s i n ki dney homogenat e wer e about 1. 5×10 CFU/ml f or l i ve bact er i a‑i nj ect ed mous e,and 6. 3×10 CFU/ml f or KB‑and l i ve bact er i a‑i nj ect ed mous e,r es pect i vel y.
Wi t h t he addi t i on of l ys os t aphi n t o t he homogenat e, t he number s decr eas ed dur i ng t he f i r s t hour and r emai ned s t eady f r om 1 t o 3 hr ,al t hough bact er i a wer e pr ol i f er at i ng t hr oughout t he i ncubat i on t i me wi t hout l ys os t aphi n. When t he ki dney homogenat e of an uni nf ect ed mous e was mi xed wi t h 2. 7×10 CFU/
ml of bact er i a and 1. 1 uni t s /ml of l ys os t aphi n,al mos t al l t he bact er i a di s appear ed dr amat i cal l y dur i ng t he f i r s t hour and keep t he s ame l evel f or anot her 2 hr . Tabl e 1 s hows t he bact er i al number s at 0 and 3 hr
af t er t he i ncubat i on and t he s ur vi val r at e of t he bact er i a dur i ng t he i ncubat i on. The di f f er ences of t he s ur vi val r at e bet ween t hr ee gr oups wer e s i gni f i - cant( p <0. 05) . I n cas e 1×10 CFU/ml of bact er i a wer e mi xed wi t h 1. 1 uni t s /ml of l ys os t aphi n,bact er i a qui ckl y di s appear ed t o be undet ect abl e ( dat a not s hown) .
Phagocytic ability of leukocytes
Fi gur e 3 s hows t he mi cr ogr aphs of pl aques dur i ng t he t i me af t er t he i nj ect i on of KB. Leukocyt es i n a nor mal condi t i on ( at 0 hr )f or med onl y ver y f ew, s mal l ,and r ound pl aques ,and t he number s and s i zes of t he pl aques i ncr eas ed wi t h t i me. Fi gur e 4 s hows t he t ot al ar ea of t he pl aques wi t hi n a cer t ai n mi cr os copi c f i el d. Bef or e t he i nj ect i on of KB( 0 hr ) ,t he val ue was 300. 5±135. 6 pi xel s . A 220‑f ol d i ncr eas e was ob- s er ved dur i ng t he f i r s t t wo hour s ,and t hen t he l evel r emai ned cons t ant f or t he next hour . Si gni f i cant di f f er ence i n val ues bet ween 2 and 3 hr was not r ecogni zed. The s i ze f r equency di s t r i but i ons of pl aques at each t i me af t er t he i nj ect i on of KB wer e i nves t i gat ed( Fi g.5) . The di s t r i but i on pat t er ns wer e di f f er ent f r om each ot her ,and t he pat t er n 2 hr af t er t he i nj ect i on of KB s hi f t ed t o t he r i ght . Some l ar ger pl aques ,whos e ar ea was gr eat er t han a r el at i ve ar ea of 600,wer e r ecogni zed 2 hr af t er t he i nj ect i on of KB.
Fig.2. Decline in lysostaphin action resulting from injec- tion of KB. One hour before the injection of live bacteria,1×10 cells of heat ‑killed counterparts were intravenously inject ed into mice. Lysosta- phin (1.76 units) was injected, if necessary.
Details are described in MATERIALS AND METHODS. Values are mean±SD (n=5). The difference between lysostaphin‑treat ed mice with and with- out pre‑injection of KB was statistically signifi- cant(,p<0.05).
Table 1. Changes in the bacterial numbers during the incubation with lysostaphi n
Live KB & Live None bacteria
(CFU/ml of homogenat e)
0 h(1.5±1.3)×10 (6.3±1.2)×10 (2.7±1.1)×10 3 h(6.7±1.3)×10 (1.3±1.3)×10 (1.4±1.5)×10 survival
rate( %) 4.55±1.63 20.98±2.42 0.55±0.21
Values are expressed as mean±SD of three experiments under each condition.
a) Mice were intravenously injected with either live,or KB and live bacteria. The ki dney homogenate of each mouse prepared as written i n MATERIALS AND METHODS, was mixed with 1.1 units/ml of lysostaphin.
b) The kidney homogenate of uninfected mouse was mixed with bacteria and 1.1 uni ts/ml of lysostaphin.
c) The bacterial numbers in each homogenate were checked at 0 and 3 hr after i ncubation.
d) The survival rate was calculated as:
bacterial numbers(3 h)×100 bacterial number s(0 h)
Mean s i zes of pl aques about each t i me wer e cal cu- l at ed( Tabl e 2) . As f or t he mean ar ea,s t at i s t i cal l y s i gni f i cant di f f er ences bet ween 0 and 3 hr ,and 1 and 2 hr wer e not .
Fi gur e 6 s hows t he t r ans i t i on of t he number s and t ypes of neut r ophi l s af t er t he KB‑i nj ect i on. The maxi mum t ot al of neut r ophi l s was s een 2 hr l at er . As f or i ndi vi dual cel l t ypes ,t he s egment ed cel l s de-
cr eas ed dr amat i cal l y wi t h t i me,whi l e t he s t ab cel l s i ncr eas ed i n number ,peaki ng dur i ng t he s ame per i od.
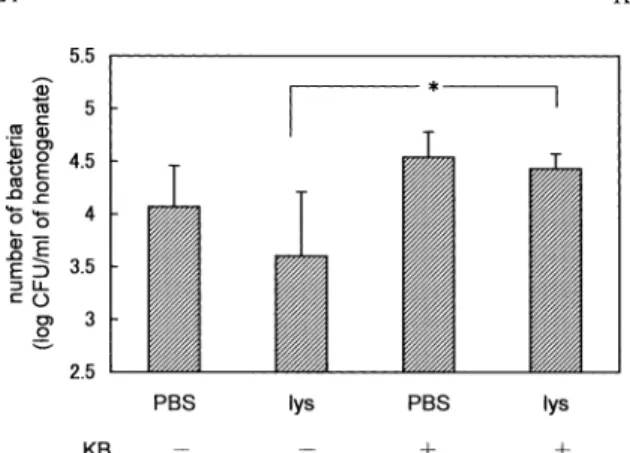
Bacter ial gr owth in kidney of leukopenic mice
The i nt r ar enal bact er i al gr owt h of nor mal and l eukopeni c mi ce was compar ed. As Fi g.7A i ndi - cat es ,mor e bact er i a wer e r ecover ed f r om l eukopeni c mi ce t han f r om nor mal mi ce. On t he ot her hand, when mi ce r ecei ved l ys os t aphi n( 1. 76 uni t s )at 1 hr af t er t he i nj ect i on wi t h l i ve bact er i a,t he bact er i al gr owt h i n bot h l eukopeni c and nor mal mi ce was near l y equal ,and t her e wer e f ewer bact er i a t han wer e obt ai ned f r om mi ce r ecei vi ng no l ys os t aphi n( Fi g.7B) .
D
ISCUSSION
Al t hough l eukocyt es ar e gener al l y cons i der ed t o cons t i t ut e a par t of t he hos t def ens e mechani s ms , t he dat a pr es ent ed her e i ndi cat e t he pos s i bi l i t i es t hat l eukocyt es pl ayed an i mpor t ant r ol e i n pr ot ect i ng t he S. aur eus or gani s ms i nges t ed by t he l eukocyt es ,and t hat s ever al ki nds of cel l s i n ki dney al s o pl ayed as t he pr ot ect or .
The r enal bact er i al number s i n mi ce gi ven wi t h
Fig.3. Micrographs of phagocytic plaques obtained from mice injected with heat‑killed bacteria. The phagocytic plaque method was carried out each time. A to D cor respond with 0,1,2,and 3 hr after the injection of KB,respectively. The numbers and sizes of plaque became larger with time.
Fig.4. Effect of heat‑killed bacteria on the phagocytic activity of leukocytes. The phagocyt ic plaque method was carried out at 0,1,2,and 3 hr after the injection of KB,and t he area of all plaques within a certain micros copic field was summed up. Details are descri bed in MATERIALS AND
METHODS. Each value i s expressed as a mean from five spots per dish. Er ror bars show the SD.
l ys os t aphi n wer e l ower t han t hos e i n mi ce wi t hout l ys os t aphi n( Fi gs .1 and 2) ,s ugges t i ng t hat ,at mos t , one‑t hi r d of t he bact er i a t hat had adher ed t o t he ki dney wer e ki l l ed by l ys os t aphi n whi l e t he r emai nder wer e not ,one r eas on bei ng t hat s ome of t he i nj ect ed l ys os t aphi n may have been i nact i vat ed by s ubs t ance( s )
i n t he bl oods t r eam. Some r epor t s s ugges t ed t hat l ys os t aphi n gi ven t o t he ani mal was abl e t o r each i nf ect i ous f oci wi t hout al t er at i on and wi t h bact er i - ci dal ef f ect i venes s . Anot her r eas on was t hat S. aur - eus or gani s ms admi ni s t er ed t o t he mi ce may have been i n a l ocat i on wher e t hey wer e pr ot ect ed f r om t he act i on of l ys os t aphi n;f or exampl e,i n l eukocyt es ,i n r enal cel l s ,or i n i nt er s t i t i al s paces bet ween r enal t i s s ues . Anot her exper i ment r eveal ed t hat t he i nt r a- venous i nj ect i on of ei t her l i ve or heat ki l l ed S. aur eus cel l s coul d i nduce neut r ophi l ‑domi nant l eukocyt os i s ( Fi g.6) . The number of t he s t ab cel l s r eached a maxi mum at 2 hr af t er t he i nj ect i on of KB,s ugges t i ng t hat i nf l ammat i on was i nduced by t he i nj ect i on of KB and t hat a l ar ge quant i t y of s t ab cel l s wer e s uppl i ed i n a s hor t t i me f r om bone mar r ow. Accor di ng t o r es ul t s obt ai ned us i ng a phagocyt i c pl aque met hod ( Fi gs .3‑5 and Tabl e 2) ,t he phagocyt i c l evel of each neut r ophi l was hi ghes t at 1 hr af t er t he i nj ect i on of KB,whi l e t he number of neut r ophi l s was maxi mal at 2 hr . The phagocyt i c f unct i on of mous e bl ood was cons i der ed t o be f ul l y act i vat ed 2 hr or mor e af t er t he i nj ect i on of KB.
I nci dent al l y,i n an in vitr o exper i ment( Tabl e 1) t he l i ve bact er i a i n t he ki dney homogenat e of a mous e gi ven bot h ki l l ed and l i ve bact er i a wer e not l ys ed by l ys os t aphi n as much as i n t he ot her t wo condi t i ons . The l i ve bact er i a i nj ect ed l at er mi ght be i nges t ed but pr obabl y not ki l l ed by t he l eukocyt es , whos e phagocyt i c act i vi t y had been s t r ongl y boos t ed by t he i nj ect i on of KB. Such l i ve bact er i a mi ght be di f f i cul t t o be l ys ed by l ys os t aphi n becaus e of bei ng pr ot ect ed wi t hi n t he l eukocyt es . When no pr e‑i nj ect i on of KB was gi ven,t he number of l i ve bact er i a i nges t ed by l eukocyt es was l ow,becaus e t he phagocyt i c act i vi t i es of nor mal mous e l eukocyt es wer e not r ai s ed as s hown i n Fi g.3A. Fur t her mor e,t he r eas on why t he bact er i a mi xed wi t h t he ki dney homogenat e of uni nf ect ed mi ce wer e i mmedi at el y l ys ed by l ys os t aphi n mi ght be t hat t he bact er i a wer e out s i de t he hos t cel l s .
The r es ul t pr evi ous l y obt ai ned i n our l abor at or y was t hat S. aur eus or gani s ms have been mor e f r e- quent l y phagocyt os ed by l eukocyt es t han coagul as e
‑negat i ve s t aphyl ococci of l ow pat hogeni ci t y.
Anot her wor k of our l abor at or y s t at ed t hat about
Fig.5. Phagocytic activity of each leukocyte. The rela- tive area of each plaque after the KB‑injection was estimated about 100 pl aques. Size frequency distributions are shown. Det ails are described in MATERIALS AND METHODS .
20% of S. aur eus or gani s ms wer e phagocyt os ed by l eukocyt es when mi xed i n t he r at i o of 1:1,and t hat over 90% of t he phagocyt os ed bact er i a wer e ki l l ed f or t he f i r s t 30 mi n i n an in vitr o exper i ment . Fur t her -
mor e,t he r es ul t was s hown t hat 30‑f ol d gr owt h was obs er ved af t er t he f ol l owi ng 21 hr ‑i ncubat i on,whi l e Staphylococcus sapr ophyticus of l ow pat hogeni ci t y wer e ki l l ed and coul d not pr ol i f er at e any mor e. Thes e phenomena may be har d t o account f or on t he bas i s of t he common vi ew t hat a man avoi ds s i cknes s as a r es ul t of i nges t i on and des t r uct i on of bact er i a by hi s l eukocyt es . Thus ,i t i s i nt er es t i ng t o cons i der what ki nd of r el at i ons hi p t her e i s bet ween t he S. aur eus char act er i s t i cs of bei ng eas y t o phagocyt os e and havi ng a hi gh pat hogeni ci t y. Thes e t wo r epor t s s ug- ges t ed t hat not ever y bact er i um phagocyt os ed was ki l l ed by t he l eukocyt es ,even i f mos t of t he bact er i a gi ven t o t he mous e wer e i nges t ed. Thi s may be expl ai ned by t he var i ous r es i s t ance f act or s pos s es s ed by a bact er i um agai ns t t he s t er i l i zi ng act i on of t he l eukocyt e,and by t he i nabi l i t y of a l eukocyt e t o s t er i l i ze ef f ect i vel y under a heavy l oad s uch as hyper - phagocyt os i s . S. aur eus wi t hi n a l eukocyt e mi ght concei vabl y be pr ot ect ed f r om at t ack by bact er i ci dal or t oxi c s ubs t ances i n s er um s uch as l ys ozyme,s ome i nt er f er ons ,or ot her s ,and can pr ol i f er at e i n t he human body af t er war ds t o make t he i nf ect i ous f oci .
Fr om t he s t andpoi nt of t he l eukocyt i c r ol e,a l euko- cyt e appear ed t o have t wo oppos i t e r ol es ,t hos e of s t er i l i zat i on and t he pr ot ect i on,agai ns t bact er i a pr es -
ent i n a hos t .
Ther e i s ot her evi dence i ndi cat i ng t hat per i pher al neut r ophi l s i nf l uence t he ci r ucul at i ng bact er i a i nt r avenous l y i nj ect ed t o a mous e. The pr evi ous r epor t ,t hat even an ext r emel y s mal l quant i t y of S.
aur eus or gani s ms coul d es t abl i s h i nf ect i ous f oci af t er di r ect i nocul at i on t o t he ki dney,s ugges t ed t hat al mos t S. aur eus or gani s ms wer e ki l l ed by ci r cul at i ng l eukocyt es bef or e t hey r eached t he ki dney. Mor e-
Table 2. Changes in size of individual plaque after injection of heat‑killedS. aureus
time after KB‑injection(h)
0 1 2 3
relative area 221.8±99.1 290.3±107.2 279.4±156.0 231.5±92.4
The size of each plaque was estimated. Values are mean±SD of 100 plaques. Details are described in MATERI ALS AND METHODS.
Significant differences were confirmed between 0 and 1 hr,0 and 2 hr,1 and 3 hr,and 2 and 3 hr,respectively(p<0. 005).
Fig.6. Appearance of stab cells after the injection of KB.
Segmented and stab cells in peripheral blood were counted after KB wer e injected. Details are described in MATERIALS AND METHODS. Open bar indicates segmented cel ls,and shaded bar indi- cates stab cells.
Fig.7. Effect of lysostaphin on bacterial proliferation in leukopenic mice. Both nor mal and leukopenic mice received S. aureus organisms. One hour after the bacterial injecti on,mice were given PBS (A)or lysostaphin(B),respectively. Details are described in MATERIALS AND METHODS. Values are mean±SD (n=5). The di fference between normal and leukopenic mi ce without lysostaphin‑
injection was statistically significant(,p<0.05).
over ,as ment i oned i n “RESULTS”,mor e bact er i a wer e s een i n l eukopeni c mi ce t han nor mal mi ce( Fi g.
7A) ,al t hough,r egar di ng l ys os t aphi n‑i nj ect ed mi ce, t he number s of bact er i a i n l eukopeni c mi ce wer e equal t o t hos e i n t hei r nor mal count er par t s( Fi g.7B) . I n l eukopeni c mous e whi ch has onl y a f ew l eukocyt es ,i t mi ght be cons i der ed t hat l ys os t aphi n s t er i l i zat i on was ef f ect i ve becaus e t he bact er i a wer e abl e t o es cape phagocyt os i s by l eukocyt es and t o mul t i pl y ef f i ci ent l y out s i de t he l eukocyt es . Fur t her mor e,t her e was a pos s i bi l i t y t hat t he bact er i a wer e al s o i nges t ed by t he cel l s ot her t han l eukocyt es i n t he ki dney t i s s ue.
Some evi dence exi s t s t hat S. aur eus may be i nt er nal - i zed i nt o,and be abl e t o mul t i pl y wi t hi n,cel l s i ncl ud- i ng r enal cel l s ,epi t hel i al cel l s ,os t eobl as t s , endot hel i al cel l s ,and f i br obl as t s ,al t hough S. au- r eus has been cl as s i cal l y des cr i bed as an excl us i vel y ext r acel l ul ar pat hogen. The bact er i a i ns i de t hes e cel l s woul d pr obabl y be har dl y af f ect ed at al l by l ys os t aphi n or by neut r ophi l s . Becaus e t he number of bact er i a di d not decr eas e ver y much under s uch condi -
t i ons ,and becaus e t hey wer e i nges t ed by neut r ophi l s , i t was concl uded t hat l ys os t aphi n excl us i vel y ki l l ed ext r acel l ul ar bact er i a,whi l e i nt r acel l ul ar bact er i a coul d es cape i t s act i on. Ther ef or e,f r om t he vi ew-
poi nt of bact er i a,t her e ar e per haps obvi ous advan- t ages t o t he mai nt enance of an i nt r acel l ul ar l ocat i on, becaus e t he bact er i a woul d be abl e t o s hel t er t hem- s el ves f r om many hos t def ens e mechani s ms s uch as ant i bodi es and s er um,or f r om ant i bi ot i cs .
Fur t her exper i ment s ar e i n pr ogr es s t o def i ne t he f at e of bot h l eukocyt es and,f or exampl e,r enal cel l s t hat have i nges t ed many S. aur eus or gani s ms . We ar e al s o at t empt i ng t o exami ne how t he bact er i a i ns i de a cel l ar e abl e t o expand i nf ect i ous f oci .
Acknowledgements :The aut hor s t hank Ms .Yoko Kobayas hi and Ms .At s uko Uchi da f or t hei r t echni cal as s i s t ances .
R
EFERENCES
1. Curran JP, Al‑Sulihi FL. Neonatal staphylococcal scaled skin syndrome:mas sive outbreak due to an un- usual phage type. Pediatrics 1980;66:285‑90.
2. Breuer K,Haussler S,Kapp A,Werfel T.Staphylococcus
aureus:colonizing features and influence of an antibacterial treatment in adul ts with atopic dermatitis. Br J Dermatol 2002;147:55‑61.
3. Sheagren JN.Staphylococcus aureus:the persistent pathogen(first of two parts) . N Engl J Med 1984;310:
1368‑73.
4. Sheagren JN.Staphylococcus aureus:the persistent pathogen(second of two par ts). N Engl J Med 1984;
310:1437‑42.
5. Watankunakorn C,Burkert T. Infective endocarditis at a large community teachi ng hospital, 1980‑1990:a review of 210 episodes. Medi cine(Baltimore)1993;72:
90‑102.
6. Thometz JG,Lamdan R,Kehl KS,Chusid MJ. Mi- crobiological tolerance in orthopaedic infections:
delayed response of septic arthritis and osteomyelitis of the hip due to infection wi th tolerant Staphylococcus aureus. J Pediatr Orthop 1996;16:518‑21.
7. Baek GH,Chung MS. Methicillin‑resistant Staphylococ- cus aureus osteomyelitis of the scaphoid from a catheter in the radial artery. J Bone Joint Surg Br 2002;84:
273‑4.
8. Kondo I,Masuda S,Kimura K,Kurosaka K,Hasegawa N. Effects of intrarenal i noculation of Staphylococcus aureus on mice. Infect Immun 1971;4:103‑9.
9. Masuda S. Increased resistance to staphylococcal infec- tion observed in splenectomized mice. Jikeikai Med J 1972;19:29‑50.
10. Hasegawa N,Kondo I,Hoshina S,Kurosaka K. Com- parison of virulence and immunity of Staphylococcus aureus in mouse kidney. Ji keikai Med J 1987;34:195‑
203.
11. Seki K,Nishihara S,Ikigai H,Masuda S. Effect of intravenous administration of heat‑killed bacterial cells on blood clearance and ki dney lodgment property of Staphylococcus aureus organi sms subsequently injected to mice. Jikeikai Med J 1988;35:275‑84.
12. Seki K,Ogasawara M,Sakurada J,Murai M,Masuda S.
Altered virulence of a pleiotropic Staphylococcus aureus mutant with a low producibi lity of coagulase and other factors in mice. Microbiol I mmunol 1989;33:981‑90.
13. Spagnolo N,Greco F,Rossi A,Ciolli L,Teti A,Posteraro P. Chronic staphylococcal os teomyelitis:a new experi- mental rat model. Infect Immun 1993;61:5225‑30.
14. Hienz SA,Sakamoto H,Flock JI,Morner AC,Reinholt FP,Heimdahl A,et al. Devel opment and characteriza- tion of a new model of hematogenous osteomyelitis in the rat. J Infect Dis 1995;171:1230‑6.
15. Yoshii T,Magara T,Miyai D,Nishimura H,Kuroki E, Furudo S,et al. Local levels of interleukin‑1 beta,‑4,‑6 and tumor necrosis factor al pha in an experimental model of murine osteomyel itis due to Staphylococcus aureus. Cytokine 2002;19:59‑65.
16. Lopes JD,dos Reis M,Brentani RR. Presence of laminin receptors in Staphylococcus aur eus. Science 1985;229:275‑6.
17. Kondo I,Miyaji J,Yoshizawa Y,Yamaguchi M. A possible role of staphylococcal cell wall proteins capable of binding with laminin and col lagen in staphylococ- cal infection. In:Mollby R,Flock,J‑I,Nord,CE.Chris- tenssen B,editors. Staphylococci and Staphylococcal Infections. Stuttgart:Gust av Fischer Verlag;1994.
p.361‑5.
18. Liang OD,Maccarana M,Flock J‑I,Paulsson M,Preis- sner KT,Wadstrom T. Multiple interactions between human vitronectin and Staphylococcus aur eus. Biochim Biophys Acta 1993;1225:57‑63.
19. Kanzaki H,Morishita Y,Akiyama H,Arata J. Adhe- sion of Staphylococcus aureus to horny layer:role of fibrinogen. J Dermatol Sci 1996;12:132‑9.
20. Patti JM,Bremell T,Krajewska‑Pietrasik D,Abdelnour A,Tarkowski A,Ryden C. The Staphylococcus aur eus collagen adhesin is a virul ence determinant in experi- mental septic arthritis. Infect Immun 1994;62:152‑61.
21. Patti JM,House‑Pompeo K,Boles JO,Garza N,Gurusid- dappa S,Hook M. Critical residues in the ligand‑bind- ing site of the Staphylococcus aureus collagen‑binding adhesion(MSCRAMM). J Bi ol Chem 1995;270:12005‑
11.
22. Elasri MO,Thomas JR,Skinner RA,Blevius JS,Beeken KE,Nelson CL,et al.Staphylococcus aur eus collagen adhesion contributes t o the pathogenesis of osteomyelitis. Bone 2002;30:275‑80.
23. Masuda S,Kondo I. The effects of intravenous adminis- tration of staphylococcal protein A in Staphylococcus
‑infected mice. Microbiol Immunol 1979;23:1223‑36.
24. Nguyen T,Ghebrehiwet B,Peerschke EI.Staphylococ- cus aureus protein A recognizes platelet gC1qR/p33:a novel mechanism for staphyl ococcal interaction with platelets. Infect Immun 2000;68:2061‑8.
25. Palmqvist N,Foster T,Tarkowski A,Josefsson E.
Protein A is a virulence factor in Staphylococcus aureus arthritis and septic death. Mi crob Pathog 2002;33:
239‑49.
26. Hawiger J,Kloczewiak M,Timmons S,Strong D,Doolit- tle RF. Interaction of fibrinogen with staphylococcal clumping factor and with pl atelets. Ann NY Acad Sci 1983;408:521‑35.
27. OʼBrien LM,Walsh EJ,Massery RC,Peacock SJ,Foster TJ.Staphylococcus aureus cl umping factor B (ClfB) promotes adherence to human type I cytokeratin 10:
implications for nasal colonization. Cell Microbiol 2002;4:759‑70.
28. Bibel DJ,Aly R,Shinefield HR,Maibach HI. The Staphylococcus aureus recept or for fibronectin. J Invest Dermatol 1983;80:494‑6.
29. Shinji H,Sakurada J,Seki K,Murai M,Masuda S.
Different effects of fibronectin on the phagocytosis of Staphylococcus aureus and coagul ase‑negative staph- ylococci by murine peritoneal macrophages. Microbiol Immunol 1998;42:851‑61.
30. Peacock SJ,Foster TJ,Cameron BJ,Berendt AR.
Bacterial fibronectin‑binding proteins and endothelial cell surface fibronectin medi ate adherence of Staphylococ- cus aureus to resting human endothelial cells. Microbi- ology 1999;145:3477‑86.
31. Shinji H,Seki K,Tajima A,Uchida A,Masuda S.
Fibronectin bound to the surface of Staphylococccus au- reus induces association of very late antigen 5 and intracellular signaling fact ors with macrophage cyto- skeleton. Infect Immun 2003;71:140‑6.
32. Seki K,Murai M,Sakurada J,Shirahige A,Kobayashi K, Hwang SM,et al. Simple method for observation of phagocytosis on bacterial t hinlayer. Microbiol Im- munol 1989;33:81‑5.
33. Sears PM,Smith BS,Pollak J,Gusik SN,Blackburn P.
Lysostaphin efficacy for treatment of Staphylococcus aureus intramammary infect ion. J Dairy Sci 1988;
71(Suppl 1):244.
34. Bramley AJ, Foster R. Effects of lysostaphin on Staphylococcus aureus infect ions of the mouse mammary glands. Res Vet Sci 1990;49:120‑1.
35. Oldham ER,Daley MJ. Lysostaphin:use of a recom- binant bactericidal enzyme as a mastitis therapeutic. J Dairy Sci 1991;74:4175‑82.
36. Murai M,Kobayashi Y,Shinji H,Sakurada J,Seki K.
Role of leukocytes in intratumoral growth of a small inoculum of Staphylococcus aur eus:Special reference to staphylocidal activity of neut rophils in an Ehrlich ascites tumor cell preparation. Jikei kai Med J 1999;46:95‑
108.
37. Murai M,Usui A,Seki K,Sakurada J,Masuda S.
Intracellular localization of Staphylococcus aureus within primary cultured mouse ki dney cells. Microbiol Im- munol 1992;36:431‑43.
38. Murai M,Seki K,Sakurada J,Usui A,Masuda S.
Effects of cytochalasins B and D on Staphylococcus aureus adherence to and ingestion by mous e renal cells from primary culture. Microbiol Immunol 1993;37:69‑73.
39. Bayles KW,Wesson CA,Liou LE,Fox LK,Bohach GA, Trumble WR. Intracellular Staphylococcus aureus escapes the endosome and induces apoptosis in epithelial cells. Infect Immun 1998;66:336‑42.
40. Kahl BC,Goulian M,van Wamel W,Herrnmann M, Simons SM,Kaplan G,et al.Staphylococcus aureus RN 6390 replicates and induces apopt osis in a pulmonary epithelial cell line. Infect I mmun 2000;68:5385‑92.
41. Hudson MC,Ramp WK,Nicholson NC,Williams AS, Nousiainen MT. Internalization of Staphylococcus au- reus by cultured osteoblasts. Microb Pathog 1995;19:
409‑19.
42. Menzies BE,Kourteva I. Internalization of Staphylococ- cus aureus by endothelial cells induces apoptosis. Infect Immun 1998;66:5994‑8.
43. Usui A,Murai M,Seki K,Sakurada J,Masuda S.
Conspicuous ingestion of Staphylococcus aureus organ- isms by murine fibroblasts in vitro. Microbiol Immunol 1992;36:545‑50.