Copyright 近畿作物・育種研究会(The Society of Crop Science and Breeding in Kinki, Japan) 43
Introduction
In most higher plants, nitrate is taken up by the roots. Extensive studies have been carried out on nitrate uptake and reduction (Crawford and Forde 2002, Hasegawa and Araki 2010) but the mechanism by which roots sense external nitrate prior to uptake is not well understood. Under field conditions, nitrate is not evenly distributed near roots. It is believed that if the nitrate concentration is very low in the area surrounding the root, the roots are able to sense nitrate diffused from and elongate toward the nitrate-rich area. Zhang and Forde (1998) devised a segmented agar plate in which the nitrate-containing zone was in between nitrate-free zones and reported that nitrate stimulated the proliferation of lateral roots in Arabidopsis. This proliferation of lateral roots for nitrate uptake is thought to be the first step of nitrate assimilation and is regulated by ANR1 (Zhang and Forde 1998) and AtNRT1.1 (Walch-Liu and Forde 2008, Krouk et al. 2010). In monocotyledonous plants such as rice and wheat, coronal roots are generated after the emergence of primary root(s). If coronal roots can sense nitrate, then the morphology of root architecture should depend on nitrate localization. In rice seedlings, several coronal roots are generated after the emergence of a primary root. In this study, the response of rice roots to nitrate was evaluated as the first step toward screening of genotypes with different nitrate sensing abilities in monocotyledonous crop plants.
Materials and methods
Two japonica rice (Oryza sativa L.) cultivars, Nipponbare and Kinmaze, were used. Nipponbare and Kinmaze are the wild type cultivar of mutants with low nitrate uptake ability (Hasegawa 1996).
Seeds were germinated on a net float in a plastic container with deionized water. When the primary root grew up to approximately 15 mm, each seedling was transferred to a glass tube (35 mm in diameter and 130 mm deep) containing 0.8% agar medium and the root was inserted into the medium. The medium was horizontally divided into half and termed a bi-layered medium (Fig. 1). The upper half (60 mm) was nitrate-free and the lower half (60 mm) contained 0, 2, 20, 200, and 2,000 μM nitrate in the form of potassium nitrate (KNO3). The
gap between the two media was less than 1 mm, which was suffused with distilled water. Seedlings were cultured at 26± 1 ℃ under continuous lighting (31.5 μmol m-2 sec-1) with
70% relative humidity in a growth chamber (Biotron LH200, Nippon Medical and Chemical Instruments, Japan). The depth of the primary root and three coronal roots were measured daily as
Root growth response to nitrate in rice (Oryza sativa L.)
Miwa Kawahara1), Yutori Hori2), Akifumi Shimizu1,2) and Hiroshi Hasegawa1,2) 1)Graduate School of Environmental Science, The University of Shiga Prefecture
2)School of Environmental Science, The University of Shiga Prefecture
(2500, Hassaka-cho, Hikone, Shiga 522 - 8533, Japan)
Summary: Root growth in rice seedlings was measured using a bi-layered agar medium, the upper half of which was nitrate free and the lower half of which contained different concentrations of nitrate. When the primary root grew up to approximately 15 mm in length, each germinated seed was transferred to a glass tube containing the bi-layered medium and cultivated at 25 ℃ under continuous lighting. In the japonica rice cultivars, Nipponbare and Kinmaze, the relative growth rate (RGR) of primary roots was highest between the first and second days after transplanting and gradually decreased. Although nitrate concentration had no effect on RGR in either cultivars, the bi-layered medium presented in this experiment would be useful for detecting nitrate sensing in roots, as change in root growth and proliferation of secondary roots were easily detected.
Key words: root growth, nitrate sensing, rice, bi-layered agar medium
Acccepted : March 8, 2012
Corresponding author:Hiroshi Hasegawa(hasegawa@ses,usp.ac.jp) J. Crop Res. 57:43 - 46(2012)
Report
Fig. 1 A bi-layered agar medium in a glass tube. The upper half is nitrate-free and the lower half contains different concentrations of KNO3.
44
a parameter of root growth until the 7th day after transplanting. Relative growth rate (RGR) was calculated as follows : RGR = (lnW2 - lnW1) / (t2 - t1) where W2 and W1 are the average depth of the roots at times t2 and t1, respectively. Each treatment contained five plants and was conducted in triplicate.
Results and Discussion
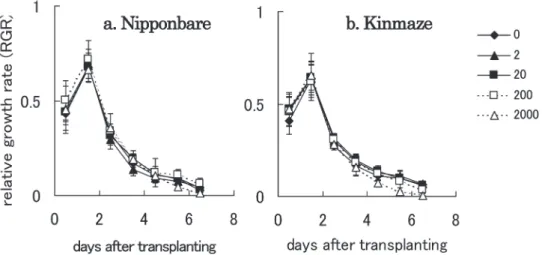
In this experiment, root growth of two japonica cultivars, Nipponbare and Kinmaze, was evaluated using a bi-layered agar medium for detecting nitrate sensing in rice roots. Similar bi-layered medium was used for detecting nitrogen response of maize roots (Tanaka et al. 2000). In both cultivars, the primary root grew linearly until the 7th day after transplanting and reached the gap between the upper and lower halves within 3 days of transplanting (Fig. 2). RGR of the primary roots was highest between the first and second days after transplanting and then gradually decreased with prolonged incubation (Fig. 3). It was assumed that when roots penetrated the gap RGR would increase, because roots elongate in order to detect and take up nitrate. However, in both cultivars used in this experiment, when the primary root reached the gap, RGR was lower than that observed the day before. Furthermore, there were no differences in root growth in regard to nitrate concentration in the lower medium except for the root growth observed in Kinmaze grown with 2,000μM nitrate. This result is in accordance with the report by Zhang and Forde (1998), which demonstrates that primary root growth was not influenced by nitrate concentration in Arabidopsis. Because coronal roots develop following emergence of the primary root(s) in monocotyledonous plants, we assumed that coronal roots can sense nitrate and influence root architecture. In this experiment, coronal roots of both
varieties did not reach the gap until the 7th day after except for the first coronal root in both cultivars. Activation of secondary root development was not observed during the experiment. However, our preliminary experiment using a bi-layered medium showed that in several local Japanese cultivars, RGR for the longest roots was highest when the roots penetrated the gap, suggesting that roots of these genotypes elongate in response to nitrate (Hasegawa et al. 2010). Therefore, the bi-layered agar medium presented in this paper may be useful to screen for nitrate sensing. However, further investigation is necessary. Several technical difficulties were observed in the screening system proposed here, which need improvement. First, in this experiment, diffusion of nitrate from the lower medium to the upper one was ignored. However, this difficulty is thought to be trivial for screening for nitrate sensing, because roots respond to low concentrations of nitrate before reaching the gap. If nitrate diffuses from the lower medium, the roots can detect nitrate before reaching the gap. Second, the validity of our hypothesis that coronal roots, in monocotyledonous plants, can respond to nitrate must be verified. Third, the present screening was carried out under continuous lighting similar to the experiment reported by Zhang and Forde (1998). However, as most roots grow in the dark, the lighting conditions may have affected root growth. Therefore, further improvements are essetial.
Walch-Liu et al. (2006) suggested that nitrate sensing can be controlled by metabolites of both photosynthesis and nitrate assimilation from the observation that accumulation of glutamate represses lateral root proliferation in Arabidopsis. In rice, mutants in which both nitrate uptake ability and nitrate reductase activity were reduced have been isolated (Hasegawa et al. 1992, Hasegawa 1996), and future studies could investigate nitrate response of roots in mutants defective for nitrate assimilation.
J. Crop Res. 57(2012)
Fig. 2 Primary root growth of Nipponbare and Kinmaze after transplanting onto a bi-layered medium. Root growth was evaluated as depth reached by the primary root in the medium. Each symbol indicates nitrate concentration in the lower half of the medium. a : Nipponbare, b : Kinmaze
45
References
Crawford, N. M. and B. G. Forde(2002)Molecular and devel-opmental biology of inorganic nitrogen nutrition. In ; The Arabidopsis Book. American Society of Plant Biologists. 1 - 25.
Hasegawa, H.(1996)Selection for mutants with low nitrate uptake ability in rice(Oryza sativa). Physiol. Plant. 199 - 204.
Hasegawa, H. and R. Araki(2010)Nitrate uptake in rice. In “Nitrogen Assimilation in Plants”Ohyama, T. and K. Sueyoshi (eds.), Research Signpost, Kelala, India. 83 - 94.
Hasegawa, H., T. Katagiri, S. Ida, O. Yatou and M. Ichii(1992) Characterization of a rice(Oryza sativa L.)mutant deficient in the heme domain of nitrate reductase. Theor. Appl. Genet. 84:6 - 9.
Hasegawa, H., Y. Hori, Y. Asano, M. Kawahara, A. Shimizu and R. Araki(2010)Screening for nitrate sensing related genotypes in rice. Abstract, Nitrogen 2010, 1st Int’l. Sympo.
on the Nitrogen Nutrition of Plants. Inuyama, Japan. 35. Krouk, G., B. Lacombe, A. Bielach, F. Perrine-Walker, K.
Malinska, E. Mounier, K. Hoyerova, P. Tillard, S. Leon, K. Ljung, E. Zazimalova, E. Benkova, P. Nancy and A. Gojon (2010)Nitrate-regulated auxin transport by NRT1.1 defines a
mechanism for nutrient sensing in plants. Developmental Cell 18:927 - 937.
Tanaka, S., A. Yamauchi, K. Yano, M. Iijima and J. Tatsumi (2000)Plastic development of maize root system in relation
to nitrogen acquisition. Root Res. 9:167-171.(in Japanese) Walch-Liu, P. and B. G. Forde(2008)Nitrate signaling mediated by the NRT1.1 nitrate transporter antagonizes L-glutamate-induced changes in root architecture. Plant J. 820 - 828. Walch-Liu, P., L.-H. Liu, T. Remans, M. Tester and B. G. Forde
(2006)Evidence that L-glutamate can act as an exogenous signal to modulate root growth and branching in Arabidopsis thaliana. Plant Cell Physiol. 47:1045 - 1057.
Zhang, H. and B. G. Forde(1998)An Arabidopsis MADS box gene that controls nutrient-induced changes in root archi-tecture. Science 279:407 - 409.
Root growth response to nitrate in rice (Oryza sativa L.)
Fig. 3 RGR of Nipponbare and Kinmaze after transplanting onto a bi-layered medium. Root growth was evaluated as depth reached by the primary root in the medium. Each symbol indicates nitrate concentration in the lower half of the medium. a : Nipponbare, b : Kinmaze
46 J. Crop Res. 57(2012)