骨格筋を構成する筋線維には遅筋タイプと速筋タイプ が存在し,タイプ組成により筋組織全体の代謝能力や運 動能力が決まる。筋線維タイプは,これまで後天的には 主として運動(身体活動量)によって変化すると考えら れていたが,我々の研究から,様々な食品栄養学的処置 で筋線維タイプを制御することが可能であることが示さ れた。筋線維タイプの制御は,ヒトの代謝能力や運動能 力の向上に応用ができるだけでなく,家畜骨格筋の特性 を変化させることから,食肉の肉質制御にも応用できる と期待される。 本稿では,筋線維タイプの指標分子であるミオシン重 鎖アイソフォームを明瞭に分離する電気泳動プロトコル の開発,ならびにその解析方法を利用して,筋線維タイ プが変化することを見出した食品栄養学的条件について 概説する。 1.筋線維タイプ 骨格筋は筋線維と呼ばれる直径 10 150 μm,長さ数 30 cm の細長い線維状の細胞が束になった組織である。 名称からは想像しにくいが,この筋線維が骨格筋を構成 するいわゆる筋細胞のことであり,多数の未分化な筋芽 細胞が分化,融合して形成された多核細胞である。筋線 維の数は生後ほとんど増加しないと考えられているが, 多核の筋線維では全ての筋核が完全に並行して分裂しな い限り,細胞分裂ができないはずである。実際にはその ような筋線維の増殖はほぼ起きないと思われる。筋線維 は収縮特性および代謝特性の違いから,大きく遅筋タイ プ(1 型,赤筋)と速筋タイプ(2 型,白筋)に分類され, 筋組織には異なるタイプの筋線維がモザイク状に入り交 じって配置されている。速筋タイプは,ほ乳類ではさら に 2A 型,2X (2D)型,2B 型のサブタイプに分類され, 先に表記したタイプほど遅筋タイプの特性が強まっていく。 筋線維はタイプによって収縮特性,代謝特性,形態特 性が異なっており(表 1),これら異なるタイプの構成 比率(組成比)が,筋組織全体の特性を左右する。筋線 維タイプ組成は,骨格筋組織によって大きく異なる。例 えば,動物実験から得られたデータによると,足関節の 底屈で主動筋となる下腿後部の筋組織の一つヒラメ筋は 姿勢維持に関与し,慢性的に活動をしているが1),多く の研究で典型的な遅筋として利用される。一方下腿前部 の前脛骨筋や長趾伸筋は主として速筋タイプで構成され ており,瞬発的な運動で用いられ,典型的な速筋として 利用されることが多い。筋線維タイプ組成は個人差(個 体差)が大きい。418 人の白人の外側広筋の筋線維タイ プを解析した結果,遅筋タイプ組成の最小は 15%,最 大は 85%とかなり幅があり,変動係数(CV)も約 30%
骨格筋線維タイプの食品栄養学的制御に関する研究
(平成 27 年度日本栄養・食糧学会奨励賞受賞)
水 野 谷 航
*,1 (2015 年 9 月 11 日受付;2015 年 10 月 23 日受理) 要旨:骨格筋を構成する筋線維には遅筋タイプと速筋タイプが存在し,タイプ組成により筋組織全体の代謝 能力や運動能力が決まる。我々は,まず筋線維タイプの指標分子であるミオシン重鎖(MyHC)アイソフォー ムを明瞭に分離する電気泳動プロトコルを開発し,食品栄養学的な処理,具体的には脂肪代謝が促進される 生理条件が筋線維タイプを変化させるか調べた。その結果 48 時間絶食させたラット骨格筋では,MyHC 組 成は変化せず,4 週間寒冷環境下(4℃)で飼育したラット骨格筋では,遅筋主体の筋組織で遅筋タイプの MyHCが増加することを明らかにした。また,魚油を配合した飼料を 4 週間ラットに給餌した結果,大豆 油摂取群に比べ,MyHC 組成が有意に遅筋タイプ側へ変化することを明らかにした。筋線維タイプは,こ れまで運動によってのみ変化すると考えられていたが,我々の研究から,食品栄養学的処理でも筋線維タイ プを制御できることが分かった。 キーワード:骨格筋,筋線維タイプ,ミオシン重鎖,寒冷暴露,魚油 *連絡者・別刷請求先(E-mail: mizunoya.wataru.185@m.kyushu-u.ac.jp) 1 九州大学大学院農学研究院資源生物科学部門動物・海洋生物科学講座(812 8581 福岡県福岡市東区箱崎 6 10 1)に達することが報告されている2)。異なる人種や民族を 含めればさらに大きな差が存在すると予想される。同一 の人間もしくは動物個体でも,先ほど説明したヒラメ筋 や長趾伸筋のように,筋組織ごとに筋線維タイプ組成が 異なる。何らかの処理による筋線維タイプ組成への影響 を調べる際は,同じ筋組織を用いることが重要であり, また研究の前提で想定している筋線維タイプの変化が遅 筋から速筋への変化か,速筋から遅筋への変化かを踏ま え,解析に供する筋組織を決定する必要があるだろう。 筋線維タイプ組成は運動パフォーマンス,疲労抵抗性, 代謝能力3),糖尿病発症,肥満4)5)に影響を与えると言わ れている。またいくつかの筋疾患では筋線維タイプの形 態的異常が生じることが知られている6)。正常な動物で は,筋線維タイプ組成は,遺伝的要因と環境的要因両方 の影響を受けて決まるとされる7)。環境的要因の代表が 身体活動である。運動トレーニングにより,筋線維タイ プは遅筋タイプへ移行し8),逆に不活動によって速筋タ イプへ移行することがよく知られている9)。これらの変 化は筋肥大,筋萎縮が生じる条件とも合致するものであ り,筋量と筋線維タイプ制御機構の深い関連がうかがえ る。筋線維を支配する運動神経にも遅筋タイプと速筋タ イプが存在し,Buller et al.10)が用いた交差性神経刺激 支配(遅筋線維を支配している遅筋タイプの運動神経を 速筋線維に移植し,速筋線維を支配している速筋タイプ の運動神経を遅筋線維に移植する)により,筋組織の収 縮特性が新しく移植された運動神経のタイプに変化する ことが報告されており,この変化は筋線維タイプの変化 を示唆している。その後の報告では除神経処理した筋線 維の直接電気刺激によっても筋組織の収縮特性を変化さ せることが可能であることが明らかにされている11)。し たがって運動による筋線維タイプ制御とは,運動神経の 種類に依存する電気刺激パターン(人為的にも誘導が可 能)によって,遅筋あるいは速筋の遺伝的プログラムが 発動する機構であると我々は予測している。いずれにし ろ,現段階では筋線維タイプ制御機構に関するデータは まだ不十分であり,遺伝的要因,環境的要因共に,どの ような遺伝学的,分子的機構でタイプの決定や変換が行 われるのか,解明が待たれる。 2.筋線維タイプがエネルギー代謝に及ぼす影響 筋線維タイプ組成は我々のスポーツパフォーマンスに 強く影響することは古くから知られており,実際エリー ト競技者の筋線維タイプ組成を調べるとスプリント競技 者では速筋線維タイプの比率が高く,逆に持久系競技者 では遅筋線維タイプの比率が高い12)。遅筋タイプはミト コンドリアを多く含み,TCA 回路や β 酸化系を構成す る酵素活性が高く,好気的代謝能力に優れている。一方 速筋線維は解糖系を構成する酵素活性およびクレアチン キナーゼ活性が高く,嫌気的代謝能力に優れている。ま た遅筋タイプの筋線維では酸素を貯蔵する色素タンパク 質であるミオグロビンの量が多く,これが赤色を呈する 原因である。褐色脂肪組織ではミトコンドリアの鉄分に より褐色が見られるが,遅筋タイプの色調にはミトコン ドリアが影響している可能性もある。 エネルギー代謝との関わりをもう少し詳しく述べる と,遅筋タイプは脂肪代謝に優れているため,安静時や 運動時の脂肪酸酸化量が高くなると推測される。実際, Wade et al.13)によると遅筋タイプ比率が高いほど運動時 の呼吸交換比が低くなる(つまり脂肪の利用が高まって いる)ことが報告されている。脂肪酸酸化能力の増加は, 肥満を予防すると期待され,この報告で遅筋タイプ比率 と体脂肪率には有意な負の相関が認められている。近年, 食生活の欧米化や運動不足などにより,日本人の 20% 以上が肥満者とされる。肥満は高脂血症,糖尿病,動脈 硬化,高血圧などの生活習慣病のトリガーとなることか ら,運動トレーニング等により遅筋タイプを増加させる ことは,生活習慣病予防の一つの有効な手段であると考 えられる。 3.筋線維タイプの測定方法とその改良について ミオシンは骨格筋に存在する筋原線維タンパク質で, アクチンと共に筋タンパク質の約 50 60%を占める。ミ オシンは運動性タンパク質 ATP アーゼとして分類され る 酵 素 で あ り,2 つ の ミ オ シ ン 重 鎖(myosin heavy chain, MyHC)および 4 つのミオシン軽鎖サブユニット からなる六量体タンパク質である。ATP アーゼ活性と アクチン結合能をもつ領域は MyHC に存在することか ら14),MyHC がミオシンタンパク質の本体とも言える。 この MyHC には複数のアイソフォームが存在し,それ ぞれ ATP アーゼ活性(すなわち分子モーターとしての 特性)が異なる。げっ歯類の成体の骨格筋では 4 種類の MyHCアイソフォーム(MyHC1, 2A, 2X, 2B)が発現し ており,右に付記された数字は,先に述べた筋線維タイ 表1 筋線維タイプの特性 遅筋 中間 速筋 収縮特性 最大張力に達するまでの時間 1.0 0.4 0.4 ミオシン ATPase 活性 1.0 3.0 3.0 代謝特性 クエン酸合成酵素活性(TCA 回路) 1.0 0.8 0.6 ホスホフルクトキナーゼ活性 (解糖系) 1.0 1.5 2.1 グリコーゲン量 1.0 1.3 1.5 トリグリセリド量 1.0 0.4 0.2 形態特性 ミトコンドリア量 1.0 0.7 0.4 毛細血管密度 1.0 0.8 0.6
Zierath & Hawley, PLoS Biology, 2004 (2), e3483)を改変。
プと対応している。したがって,現在のところ,筋線維 タイプ組成とは,MyHC アイソフォームの組成と同義 である。ただし,ヒトにおいて,後述するミオシン ATPアーゼ染色法により 2B 型と分類された筋線維に含 まれる MyHC アイソフォームは,アミノ酸配列の相同 性から実際は MyHC2X であり15),MyHC2B はヒトの骨 格筋では発現がほとんど見られない(ゲノムに MyHC- 2B (MYH4)遺伝子は存在する)。興味深いことに,多 くの筋線維(筋細胞)で,1 本の筋線維はたった 1 種類 の MyHC アイソフォームしか発現していない(pure fiber)。一方,1 本の筋線維内に 2 種類以上の MyHC ア イソフォームを発現する筋線維も存在する(hybrid fi-ber)。その割合としてラット後肢筋の解析では単離した 筋線維の 90%以上が pure fiberで16),ヒトの外側広筋で は約 90%が pure fiber という報告もあるが17),約 70% とする報告もある18)。 筋線維タイプを分類するための最も古典的で一般的な 手法は,ミオシン ATP アーゼ染色法である19)。この方 法は,MyHC アイソフォームによって pH に対する安定 性が異なることを利用して筋線維を分類する染色方法で ある。非常によく普及した方法であるが,欠点として, 複数の連続凍結切片を作成しなければならないことと, 染色後に組織切片内の同一箇所を照合することが大変に 手間であること,骨格筋組織全体の一部分(厚さ 10 μm 程度)しか観察できないことが挙げられる。ミオシン ATPアーゼの検出を酵素活性の代わりに抗体で検出す るいわゆる免疫染色も最近ではよく利用されている。特 定の MyHC アイソフォームを特異的に認識するモノク ローナル抗体は既に確立されており,実験操作手順こそ 異なるが,最終的には ATP アーゼも抗 MyHC 抗体の免 疫染色も類似の染色パターンを得ることができる。ただ し,これらの抗体は全てマウスのモノクローナル抗体で あるため同一切片の四重染色は実現できていない。 組織染色に替わる MyHC アイソフォーム評価法とし て,SDS ポリアクリルアミドゲル電気泳動法(SDS-PAGE)が知られている。MyHC はどのアイソフォーム も約 200 kDa で,一般的なポリアクリルアミドゲルでは ほとんど分離されないが,Talmadge & Roy20)は 30%の
グリセロールを含む特殊な組成の分離ゲルを用い,長時 間電気泳動を行うことで,MyHC アイソフォームをミ ニゲルで分離することに成功した。この報告ではラット の筋サンプルを用いているが,マウス21),ウサギ22),お 表2 ミオシン重鎖分類法の特徴 切片作成 染色後の解析作業 全筋の解析 MyHC算出組成比の 形態と局在の情報 スループット ミオシン ATP アーゼ染色 要 要 不可 可 有 × 抗 MyHC 抗体を用いた免疫染色 要 要 不可 可 有 × SDS-PAGE 不要 不要 可 可 無 ○ MyHC mRNAの(リアルタイム)RT-PCR* 不要 不要 可 不可 無 ◎
抗 MyHC 抗体を用いた Western blotting 不要 不要 可 不可 無 ○
*厳密には MyHC の mRNA レベルとタンパク質レベルが一致しないとの報告もある25)。
MyHC mRNAの in situ hybridization による評価も可能だが,一般的ではないためリストには挙げていない。
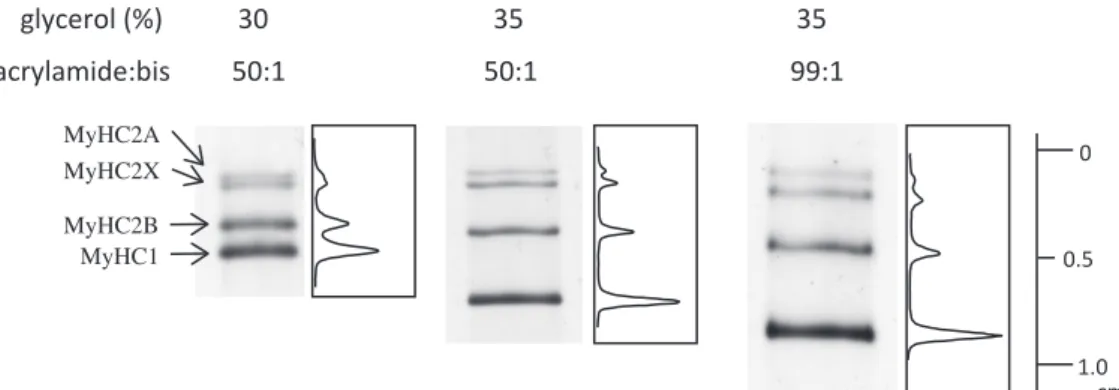
図1 分離ゲルのグリセロール濃度とビスアクリルアミド比を変化させることでミオシン重
鎖(MyHC)アイソフォーム分離能が改善した
Talmadge & Roy20)が開発した SDS-PAGE による MyHC 分離プロトコル(左レーン)は,
ミニゲルで MyHC 組成比を簡便に測定できる画期的な方法であったが,MyHC2A と MyHC- 2X の分離は不十分であった。我々は分離ゲルの成分濃度を少し変化させるだけで MyHC 分離能が劇的に改善することを見出した(Analytical Biochemistry, 2008 (377), 111 11324)よ
よびヒト23)でも応用可能なことが報告されている。ミ
オシン ATP アーゼ染色法と比較して,多数のサンプル の処理が短時間で実施できること,筋組織全てをホモジ ナイズすることで部位ごとの偏りのない解析が可能とな ること,Western blotting アッセイにも応用ができると いった利点がある。しかし Talmadge & Roy の報告にあ る泳動像でも示されているが20),MyHC2A と MyHC2X アイソフォームの分離は十分ではなかった。我々は,分 離ゲルのグリセロールおよび架橋剤濃度および試料調製 試薬を変更することにより,非常に良好な MyHC アイ ソフォームの分離を達成できた(図 1)24)。 他に筋線維タイプを評価する方法としては,(リアル タイム)RT-PCR による MyHC mRNA の定量25)や特異 抗体を用いた Western blotting も用いられている。これ らの筋線維タイプを評価する手法の特徴を表 2 にまとめた。 4.食品栄養学的な処理が 筋線維タイプ組成に及ぼす影響 我々は,先に述べた改良 SDS-PAGE 法を用いて,脂 肪代謝が促進される生理条件がラットの筋線維タイプを 遅筋タイプへ移行させるか調べた。全身の脂肪代謝の促 進は,すなわち身体最大のエネルギー消費器官である骨 格筋の脂肪代謝を促進する条件であり,骨格筋は環境へ の適応として遅筋タイプを増加させるのではないかと着 想した。現在まで 48 時間絶食,寒冷暴露,魚油,リン ゴポリフェノールについての結果を発表しているので以 下に紹介する。 4.1 48 時間絶食26) 絶食は運動と類似した代謝変化を引き起こす。例えば 絶食によって骨格筋のリポプロテインリパーゼや fatty acid translocase (FAT/CD36)といった脂肪酸酸化に関 わる遺伝子発現が増加する27)。一方,筋線維タイプ組成 への影響はどうかというと,まず絶食によってタンパク 質分解が促進されるのは速筋タイプであると言われてい る28)。これらを考慮して,絶食によって遅筋タイプ組成 の増加が生じると,当初は予想していた。しかしラット に 48 時間の絶食をかけた結果,ヒラメ筋,長趾伸筋共 に MyHC 組成は変化しなかった。むしろ長趾伸筋の MyHC2B の mRNA 発現は対照群より有意に増加してい た。絶食によって遅筋タイプ移行を誘導する peroxi-some proliferator-activated receptor gamma coactivator 1α (PGC-1α)の発現が低下し29),速筋タイプ移行を誘導
する forkhead box protein O1(FOXO1)の活性化(脱 リン酸化)が生じていたので27),確かに遅筋タイプへの 移行を抑制する応答も見られている。脂肪代謝に関わる 因子は絶食でかなり増加していたのとは対照的に, MyHC組成については短期間の環境変化で変化させな い恒常性が生体には備わっているのかもしれない。さら にこれらの結果から骨格筋の代謝特性と収縮特性が,必 ずしも連動していないことが示唆された。 4.2 寒冷暴露30) 寒冷暴露も運動と類似した代謝変化を引き起こす。例 えば 11℃の環境でラットを 3 週間飼育すると,骨格筋 のミトコンドリアが有意に増加することが報告されてい る31)。寒冷暴露によってラット腓腹筋外側で若干遅筋タ イプが増加することがミオシン ATP アーゼ染色により 調べられていたが32),筋組織全体の MyHC 組成へ及ぼ す影響は調べられていなかったので,改良 SDS-PAGE 法を用いて MyHC 組成の変化を中心に検討を行った。 ラットを 4 週間,4℃環境下で飼育した結果,遅筋タイ プが主体のヒラメ筋でさらなる遅筋タイプへの移行が見 ら れ,MyHC1 組 成 は 対 照 群 の 約 90 % に 対 し, ほ ぼ 100%まで増加していた。しかし,長趾伸筋と腓腹筋と いった速筋主体の筋組織では MyHC 組成に変化が見ら れなかった。速筋主体の筋組織の多くで有意な変化が見 られなかった点は Matoba & Murakami の結果と一致し ていた32)。一方,代謝に関わる因子はどの筋組織でも上 昇していたので,こちらの結果からも,骨格筋の代謝特 性と収縮特性が,独立して制御されていることが示唆さ れる。 4.3 魚 油33) 魚油摂取は動物の脂肪代謝を亢進し,内臓脂肪の蓄積 を抑制する作用があることが知られている34)。魚油の脂 肪代謝亢進は,骨格筋の筋線維タイプを遅筋タイプへ変 化させる可能性を考え,我々は魚油の 4 週間摂取がラッ ト骨格筋の筋線維タイプに及ぼす影響を調べた。その結 果,魚油摂取群では,速筋主体の長趾伸筋で速筋タイプ の MyHC2B が減少し,代わりに中間タイプの MyHC2X の比率が,大豆油摂取群と比べ有意に増加することが明 らかとなった(図 2)。魚油摂取では速筋サブタイプ間 の遅筋方向への移行に合わせ,脂肪代謝に関わる遺伝子 発現が有意に増加していた。一方遅筋主体のヒラメ筋で は,MyHC 組成および代謝に関わる因子共に変化はな かった。ヒラメ筋でも運動トレーニングによって酸化的 代謝能力が高まると言われているが,速筋主体の筋組織 より応答性は低いことが示唆されているため35),この魚 油の結果は,非常に運動トレーニングに近い変化と言える。 4.4 リンゴポリフェノール36) リンゴ幼果から抽出したリンゴポリフェノール(AP) は,カテキンが多数重合したプロシアニジン類を主成分 とする。これまでの研究で,抗酸化作用37)や抗肥満作 用38)などの生理活性が報告されている。我々はこの AP の摂取がラットの筋線維タイプに及ぼす影響を調べた。 5%の AP を含む飼料をラットに 8 週間与えた結果,速 筋主体の足底筋において,速筋タイプの MyHC2B が減 少し,代わりに中間タイプの MyHC2X の比率が対照群 と比べ有意に増加していた。この変化は魚油摂取と類似 していた。この研究ではさらに筋持久力についても検討 を行った。その結果 AP 群では対照群と比較して筋持久 力が約 1.5 倍まで有意に増加することを確認した。この
変化は遅筋タイプの増加と対応していると考えられ,運 動トレーニングをしなくても,AP 摂取により速筋サブ タイプ間の遅筋方向への移行が生じ,その変化は筋機能 にも反映されることを意味している。 お わ り に 以上の研究成果から,食品成分の中には,運動トレー ニングを行わなくても,運動トレーニングと類似した筋 線維タイプ変化を引き起こす物質が存在することが明ら かとなった。また興味深いことに,筋線維タイプ組成が 異なる筋組織では(例えばヒラメ筋と長趾伸筋),刺激 に対する応答性がかなり異なる点がどの実験でも共通し ていた。確認はまだされていないが,例えばヒトで遅筋 タイプの比率が高い者と,速筋タイプの比率が高い者で は,ここで紹介した食品栄養学的な刺激も含め,多くの 環境要因に対し応答性が異なるのかも知れない。 骨格筋を対象とした研究において,筋肥大や筋萎縮を 食品栄養学的に制御することを試みた研究に比べると, 筋線維タイプを制御しようと試みた研究はそれほど多く はないため,今後多くの発見が期待できる分野である。 筋線維タイプの人為的制御が実現できれば,ヒトの代謝 能力や運動能力の向上への応用にとどまらない。筋線維 タイプは食肉の肉質(肉の栄養価や物性ならびに美味し さ)にも影響があると言われているためである。現在, 我々は筋線維タイプ制御を食肉生産に応用することに取 り組んでおり,具体的には,ある飼料や飼育方法を施せ ば,狙った肉質(筋線維タイプ)を得られる技術を,エ ビデンスベースで確立することを目標にしている。 本稿を終えるにあたり,栄誉ある日本栄養・食糧学会 奨励賞を授与されたことに対し,選考委員の先生方をは じめ,関係の諸先生,ご推薦頂きました京都大学大学院 農学研究科(現龍谷大学農学部食品栄養学科)伏木亨教 授に謹んで御礼申し上げます。特に,伏木亨教授には学 部 4 回生から大学院博士課程までの 6 年間研究者として の基礎を指導して頂き,感謝の念に堪えません。京都大 学大学院農学研究科の河田照雄教授,井上和生准教授, 都築巧助教にも多くのご指導を賜ったことを心より御礼 申し上げます。本研究は,九州大学大学院農学研究院畜 産化学研究室にて行われたものです。畜産化学研究室で 本研究を支えて頂いた九州大学大学院農学研究院池内義 秀教授,辰巳隆一准教授に厚く御礼申し上げます。共同 研究者としてご協力頂いた九州大学大学院農学研究院中 村真子准教授,佐藤匡央准教授,城内文吾助教,福岡女 子短期大学澤野祥子講師,宇都宮大学農学部佐藤祐介助 教,京都大学大学院農学研究科後藤剛准教授,東京農業 大学応用生物科学部高橋信之准教授に心より感謝申し上 げます。共に実験を進めた畜産化学研究室の大学院生, 学部学生には本当に感謝しています。さらに PI として の在り方を教授頂きましたカナダ国マニトバ大学 Judy Anderson教授に心より感謝申し上げます。最後に,研 究室で研究を始めてから現在まで公私ともに常に尊敬す 図2 大豆油,魚油,ラード 4 週間摂取後のラット骨格筋 MyHC アイソフォーム組成 雄性 Wistar ラットに 15%の大豆油,魚油,ラードを含む飼料を与え 4 週間飼育した後,長趾伸筋(EDL)とヒラメ 筋(Soleus)の MyHC アイソフォーム組成を SDS-PAGE で解析した。魚油群は大豆油群より MyHC2B が有意に低く, MyHC2X が有意に高くなっていたことから,我々は運動に依存せず,食品成分でも筋線維タイプが変化することを 見出した。平均値±標準誤差,n=6。各 MyHC アイソフォームにおいて,3 群間の比較を,一元配置分散分析の後, Tukey-Kramer multiple-comparison testにより行った。異なる文字は,群間の有意差を示している(p < 0.05)(PLoS ONE, 2013 (8), e8015233)より転載)。
べき先輩としてご指導を頂いた龍谷大学農学部食品栄養 学科石原健吾准教授に心からお礼申し上げます。 本研究は MEXT/JSPS 科研費(18780096,22580136, 26712023),伊藤記念財団,上原記念生命科学財団の研 究助成を受けたものです。 文 献
1) Hnik P, Vejsada R, Goldspink DF, Kasicki S, Krekule I (1985) Quantitative evaluation of electromyogram
ac-tivity in rat extensor and flexor muscles immobilized at different lengths. Exp Neurol 88: 515 28.
2) Simoneau JA, Bouchard C (1989) Human variation in skeletal muscle fiber-type proportion and enzyme ac-tivities. Am J Physiol 257: E567 72.
3) Zierath JR, Hawley JA (2004) Skeletal muscle fiber type: influence on contractile and metabolic proper-ties. PLoS Biol 2: e348.
4) Schuler M, Ali F, Chambon C, Duteil D, Bornert JM, Tardivel A, Desvergne B, Wahli W, Chambon P, Metzger D (2006) PGC1alpha expression is con-trolled in skeletal muscles by PPARbeta, whose abla-tion results in fiber-type switching, obesity, and type 2 diabetes. Cell Metab 4: 407 14.
5) Wang YX, Zhang CL, Yu RT, Cho HK, Nelson MC, Bayuga-Ocampo CR, Ham J, Kang H, Evans RM (2004) Regulation of muscle fiber type and running
endurance by PPARdelta. PLoS Biol 2: e294.
6) D Amico A, Bertini E (2008) Congenital myopathies.
Curr Neurol Neurosci Rep 8: 73 9.
7) Simoneau JA, Bouchard C (1995) Genetic determin-ism of fiber type proportion in human skeletal mus-cle. FASEB J 9: 1091 5.
8) Fitzsimons DP, Diffee GM, Herrick RE, Baldwin KM (1990) Effects of endurance exercise on isomyosin patterns in fast- and slow-twitch skeletal muscles. J
Appl Physiol 68: 1950 5.
9) Caiozzo VJ, Baker MJ, McCue SA, Baldwin KM (1997) Single-fiber and whole muscle analyses of MHC isoform plasticity: interaction between T3 and unloading. Am J Physiol 273: C944 52.
10) Buller AJ, Eccles JC, Eccles RM (1960) Interactions between motoneurones and muscles in respect of the characteristic speeds of their responses. J Physiol 150: 417 39.
11) Lomo T, Slater CR (1980) Acetylcholine sensitivity of developing ectopic nerve-muscle junctions in adult rat soleus muscles. J Physiol 303: 173 89.
12) Saltin B, Henriksson J, Nygaard E, Andersen P, Jans-son E (1977) Fiber types and metabolic potentials of skeletal muscles in sedentary man and endurance runners. Ann N Y Acad Sci 301: 3 29.
13) Wade AJ, Marbut MM, Round JM (1990) Muscle fi-bre type and aetiology of obesity. Lancet 335: 805 8. 14) Rayment I (1996) The structural basis of the myosin
ATPase activity. J Biol Chem 271: 15850 3.
15) Smerdu V, Karsch-Mizrachi I, Campione M, Lein-wand L, Schiaffino S (1994) Type IIx myosin heavy
chain transcripts are expressed in type IIb fibers of human skeletal muscle. Am J Physiol 267: C1723 8. 16) Staron RS, Pette D (1993) The continuum of pure
and hybrid myosin heavy chain-based fibre types in rat skeletal muscle. Histochemistry 100: 149 53. 17) Galpin AJ, Raue U, Jemiolo B, Trappe TA, Harber
MP, Minchev K, Trappe S (2012) Human skeletal muscle fiber type specific protein content. Anal
Bio-chem 425: 175 82.
18) Paoli A, Pacelli QF, Cancellara P, Toniolo L, Moro T, Canato M, Miotti D, Reggiani C (2013) Myosin iso-forms and contractile properties of single fibers of human Latissimus Dorsi muscle. Biomed Res Int 2013: 249398.
19) Brooke MH, Kaiser KK (1970) Muscle fiber types: how many and what kind? Arch Neurol 23: 369 79. 20) Talmadge RJ, Roy RR (1993) Electrophoretic
separa-tion of rat skeletal muscle myosin heavy-chain iso-forms. J Appl Physiol 75: 2337 40.
21) Blough ER, Rennie ER, Zhang F, Reiser PJ (1996) Enhanced electrophoretic separation and resolution of myosin heavy chains in mammalian and avian skel-etal muscles. Anal Biochem 233: 31 5.
22) Bacou F, Rouanet P, Barjot C, Janmot C, Vigneron P, d Albis A (1996) Expression of myosin isoforms in denervated, cross-reinnervated, and electrically stim-ulated rabbit muscles. Eur J Biochem 236: 539 47. 23) Kohn TA, Myburgh KH (2006) Electrophoretic
sepa-ration of human skeletal muscle myosin heavy chain isoforms: the importance of reducing agents. J
Physi-ol Sci 56: 355 60.
24) Mizunoya W, Wakamatsu J, Tatsumi R, Ikeuchi Y (2008) Protocol for high-resolution separation of
ro-dent myosin heavy chain isoforms in a mini-gel elec-trophoresis system. Anal Biochem 377: 111 3. 25) Vadaszova A, Hudecova S, Krizanova O, Soukup T
(2006) Levels of myosin heavy chain mRNA tran-scripts and content of protein isoforms in the slow so-leus muscle of 7 month-old rats with altered thyroid status. Physiol Res 55: 221 5.
26) Mizunoya W, Sawano S, Iwamoto Y, Sato Y, Tatsumi R, Ikeuchi Y (2013) Effect of 48 -h food deprivation on the expression of myosin heavy-chain isoforms and fiber type-related factors in rats. J Nutr Sci Vitaminol 59: 289 98.
27) Kamei Y, Mizukami J, Miura S, Suzuki M, Takahashi N, Kawada T, Taniguchi T, Ezaki O (2003) A fork-head transcription factor FKHR up-regulates lipopro-tein lipase expression in skeletal muscle. FEBS Lett 536: 232 6.
28) Goodman CA, Kotecki JA, Jacobs BL, Hornberger TA (2012) Muscle fiber type-dependent differences in the regulation of protein synthesis. PLoS One 7: e37890.
29) Lin J, Wu H, Tarr PT, Zhang CY, Wu Z, Boss O, Mi-chael LF, Puigserver P, Isotani E, Olson EN, Lowell BB, Bassel-Duby R, Spiegelman BM (2002) Tran-scriptional co-activator PGC-1 alpha drives the for-
mation of slow-twitch muscle fibres. Nature 418: 797 801.
30) Mizunoya W, Iwamoto Y, Sato Y, Tatsumi R, Ikeuchi Y (2014) Cold exposure increases slow-type myosin heavy chain 1 (MyHC1) composition of soleus mus-cle in rats. Anim Sci J 85: 293 304.
31) Buser KS, Kopp B, Gehr P, Weibel ER, Hoppeler H (1982) Effect of cold environment on skeletal muscle mitochondria in growing rats. Cell Tissue Res 225: 427 36.
32) Matoba H, Murakami N (1981) Histochemical changes of rat skeletal muscles induced by cold accli-mation. Jpn J Physiol 31: 273 8.
33) Mizunoya W, Iwamoto Y, Shirouchi B, Sato M, Komi-ya Y, Razin FR, Tatsumi R, Sato Y, Nakamura M, Ikeuchi Y (2013) Dietary fat influences the expres-sion of contractile and metabolic genes in rat skeletal muscle. PLoS One 8: e80152.
34) Belzung F, Raclot T, Groscolas R (1993) Fish oil n-3 fatty acids selectively limit the hypertrophy of
ab-dominal fat depots in growing rats fed high-fat diets.
Am J Physiol 264: R1111 8.
35) Spangenburg EE, Brown DA, Johnson MS, Moore RL (2009) Alterations in peroxisome proliferator-acti-vated receptor mRNA expression in skeletal muscle after acute and repeated bouts of exercise. Mol Cell
Biochem 332: 225 31.
36) Mizunoya W, Miyahara H, Okamoto S, Akahoshi M, Suzuki T, Do MK, Ohtsubo H, Komiya Y, Lan M, Waga T, Iwata A, Nakazato K, Ikeuchi Y, Anderson JE, Tatsumi R (2015) Improvement of endurance based on muscle fiber-type composition by treatment with dietary apple polyphenols in rats. PLoS One 10: e0134303.
37) Akazome Y (2004) Characteristics and physiological functions of polyphenols from apples. Biofactors 22: 311 4.
38) Nakazato K, Song H, Waga T (2006) Effects of die-tary apple polyphenol on adipose tissues weights in Wistar rats. Exp Anim 55: 383 9.
J Jpn Soc Nutr Food Sci 69: 3 9(2016)
Review
Study on Regulation of Skeletal Muscle Fiber Type by Nutritional Components
(Young Investigator Award of JSFNS (2015))
Wataru Mizunoya
*,1(Received September 11, 2015; Accepted October 23, 2015)
Summary: There are two major muscle fiber types: type 1 fibers (slow-twitch oxidative) and type 2 fibers
(fast-twitch glycolytic). Muscle fiber type is a critical physiological property that affects sports performance as well as metabolic ability. First, we developed an electrophoresis protocol that allows clear separation of the myo-sin heavy chain (MyHC) isoforms, which are common molecular markers of muscle fiber type. Then we exam-ined the effects of nutritional treatment on muscle fiber type in rats. Food deprivation for 48 h did not alter the MyHC composition of skeletal muscles. Exposure to cold (4℃) for 4 weeks increased slow-type MyHC1 in the soleus muscle. Intake of fish oil for 4 weeks induced significantly lower levels of fast-type MyHC2B and higher levels of the intermediate-type MyHC2X composition in the extensor digitorum longus (EDL) muscle in com-parison with soybean oil intake. These results clearly indicate that muscle fiber type can be modified by nutri-tional components.
Key words: skeletal muscle, muscle fiber type, myosin heavy chain, cold exposure, fish oil
*Corresponding author (E-mail: mizunoya.wataru.185@m.kyushu-u.ac.jp)
1 Department of Bioresource Sciences, Faculty of Agriculture, Kyushu University, 6 10 1 Hakozaki,