原著論文
ヨモギホンヤドカリの性的対立:
オスによる交尾前ガードの試みに対するメスの拒絶行動
Sexual conflict in the hermit crab Pagurus nigrofascia:
Female avoidance of male initiation of precopulatory guarding
木戸結菜
1・伊與田朋未
1・石原
(安田)
千晶
1・和田 哲
1Yuina Kido, Tomomi Iyoda, Chiaki I. Yasuda, and Satoshi Wada
AbstrAct: This paper described female guarding avoidance
behaviors against precopulatory guarding attempts by males and examines the effect of the number of days until female prenuptial molting on rejection behaviors in the hermit crab Pagurus nigrofascia. We used three types of females in an experiment; (1) ovigerous females (OV–female), (2) females which had not been guarded in the field (NG–female), and (3) females which had been guarded by other males in the field (G–female). We placed a male that had been solitary and a female of either type in a small container, observed the interaction, and compared the female behaviors among the three types of female. The frequencies of retreat and escape from approaching male were highest in OV–females, and lowest in G–females. During male assessment behaviors of females, frequencies of struggle and escape in OV– and G– females were higher than NG–females. 33 of 51 G–females were guarded by males although OV– and NG–females were rarely guarded by males. Many G–females showed rejection behavior during male guarding. However, no significant ef-fect was found in the number of days until the female pre-nuptial molt on the frequencies of rejection behavior.
Key Words: sexual conflict, female rejection, hermit crab,
mating behavior はじめに 有性生殖を行う生物において,繁殖は両性の個体 にとって自身の適応度に直結する重要なイベントで ある.しかし,繁殖に要するコストと利益はしばしば 雌雄で異なる.このような繁殖に関する利害の不一 致から雌雄間に生じる対立を性的対立と言う (Parker,
1979; Arnqvist & Rowe, 2005). 性 的 対 立 は 哺 乳 類 (Aloise King et al., 2013),鳥類 (Szentirmai et al., 2007), 爬虫類 (Shine et al., 2000),魚類 (Magurran & Seghers, 1994),昆虫類(Rowe et al., 1994),甲殻類(Jormalain-en, 1998; Takeshita et al., 2011)など多くの分類群で知 られている. オスがメスをガードする状況は,性的対立が生じ る典型的な状況である(Jormalainen, 1998).ガード 行動は,一般にオスが交尾・受精機会を獲得するた めの行動だと考えられており,交尾前ガード行動と 交尾後ガード行動に大別される.このうち交尾前 ガード行動は甲殻類の多くの種で知られている.交 尾前ガード行動は両性にとってコストとなる.オス はガードの間,配偶相手であるメスを運搬するため, エネルギー消費や摂餌速度の低下,捕食リスクの増 加などのコストを受ける (Jormalainen et al., 2001; 吉 井ら,2009).また,ガード中は自身がガードして いるメスを狙うライバルオスとの間に闘争が起こる (Jivoff & Hines, 1998; Takeshita & Henmi, 2010).ガー ド時間が長いほどライバルオスとの遭遇回数は増加 すると考えられるが,オス間闘争では先にメスを保 有 し て い る ほ う が 有 利 で あ る (Wada et al., 1999;
1 北海道大学大学院水産科学研究院海洋生物学講座
〒041–8611 北海道函館市港町3–1–1
Laboratory of Marine Biology, Graduate School of Fisher-ies Sciences, Hokkaido University, Minato-cho, Hakodate, Hokkaido 041–8611, Japan
Takeshita & Henmi, 2010).そのため,性比や密度を 操作した実験ではライバルオスとの遭遇頻度が高い 場合にガード時間が長くなることがわかっている (Jormalainen & Shuster, 1999; Wada et al., 1999).交尾 前ガード時間に関する数理モデルもまた,性比がオ スに偏り競争相手が多い場合や自分と競争相手の闘 争能力が拮抗している場合,そしてメスとの遭遇頻 度が低い場合には,オスのガード時間が長くなるこ とを予想している(Grafen & Ridley, 1983; Yamamura, 1987).
メスにとってもガードは交尾・受精機会を保証す る行動だが,メスはガード中に自身の行動が制限さ れるために,摂餌機会の減少や捕食リスクの増加な どのコストを受ける (Jormalainen et al., 2001; Cothran, 2004; Takeshita et al., 2011).これらのコストはガー ド時間の延長に伴って増加するが,メスがガードに よって受ける利益はガードが長期化しても変化しな いため,長時間のガードで得られるメスの適応度上 の純利益(利益とコストの差)はオスよりも小さい と考えられる.したがって,最適な交尾前ガード時 間は雌雄で異なる(Yamamura & Jormalainen, 1996). このような利害の対立から,オスによるガードの強 制行動とメスによる逃走や抵抗などの拒絶行動がし ばしば観察される(Jormalainen, 1998).ヘラムシ類 の研究ではメスの拒絶行動によりガードの形成が妨 げられ,その結果として,オスが小型である場合や, メスが大型である場合ほどガード時間が短くなるこ とが示されている(Jormalainen & Merilaita, 1995; Mi-ura & Goshima, 2016).
ホンヤドカリ属では,オスが産卵間近なメスの貝 殻の縁を左鋏脚でつかみ,数日間連れ歩く交尾前 ガード行動が知られている(Hazlett, 1966; Imafuku, 1986).ガード中のオスがライバルオスと遭遇する と,ガードされているメスをめぐってしばしばオス 間闘争が起こる.また,複数種でオスに偏った性比 の下でガード時間が長くなることが報告されている (Wada et al., 1999; Minouchi & Goshima, 2000; 吉井ら,
2009).メスはガードによって何らかのコストを 負っており,ガード時間の延長に伴いコストも増加 すると考えられる.しかし,本属ではオスの交尾前 ガードに対してメスが拒絶を示すという記載はほと んどなく,メスの拒絶行動に着目した研究は著者ら が知る限り皆無である.ホンヤドカリPagurus filholi やテナガホンヤドカリP. middendorffiiでは体サイズ の性差が大きいために,メスはオスに対してほとん ど拒絶できないとみなされている (Wada et al., 1999;
Okamura & Goshima, 2010; Kawaminami & Goshima, 2015).しかし,これら2種に比べ,ヨモギホンヤ ドカリP. nigrofasciaは体サイズの性差が小さく,メ スも大きいため(吉井ら,2009; 小島ら,2016),メ スの拒絶行動が期待できる.また,ヨモギホンヤド カリのメスは年一回産卵で(Goshima et al., 1996), 交尾直前に必ず脱皮するため(Suzuki et al., 2012), メスの次の交尾機会までの時間の比較が容易である. そこで,本研究ではヨモギホンヤドカリを対象種と して,以下の2点を検証した;(1)オスが交尾前 ガードを開始する前後でメスは何らかの拒絶行動を 示すのか,(2)メスの拒絶度合いは,オスの体サイ ズ,メスの体サイズ,メスの交尾直前脱皮までの日 数(メスの成熟度合の指標)によって異なるのか. 材料および方法 ヨモギホンヤドカリP. nigrofasciaは,北海道から 九州までの日本沿岸の岩礁潮間帯に広く分布する種 である.函館湾における本種の交尾・産卵期は4月 下旬から6月上旬までと短く,メスは交尾後すぐに 産 卵 し て 翌 年 の2月まで抱卵する(Goshima et al., 1996).そのため,交尾・産卵期のメスは,オスと の交尾を終えた直後の抱卵メス,ガードに至ってい ない未抱卵メス,そしてオスにガードされているメ ス(以下ガードメス)という3タイプに大別できる. これらの3タイプのメスは,次の交尾機会までの時 間が異なる. 2017年4月13日から5月24日の干潮時に,北海 道函館湾西岸に位置する葛登支岬周辺(41°44′ N, 140°36′ E)の平磯潮間帯でヨモギホンヤドカリの単 独個体と交尾前ガード中のペアを採集した.発見し た全ての個体とペアは海水を入れたポリエチレン製 のチャック付き小袋に個別に入れて実験室に持ち 帰った.実験室では,ペアがまだ交尾・産卵してお らず,ガード中であることを確認した後すぐにオス とメスを引き離した.採集した単独個体は性判別の ため実体顕微鏡下で第一腹肢の有無と抱卵の有無を
観察して,単独オスと抱卵メス・未抱卵メスに分け, 水深2.5 cm程度になるまで海水を入れた小型コンテ ナ(13×9×7 cm)に個別に入れた.全ての個体を 約1時間静置した後に以下の実験を行った.なお, 実験と飼育には採集地から汲んできた天然海水を使 用した.また,付属肢に欠損が認められた個体は実 験には用いなかった. メスの次の交尾・産卵までの時間に基づいて以下 の3タイプのメスを用いた;(1)採集時に単独だっ た抱卵メス (OV–メス),(2) 採集時に単独だった未 抱卵メス (NG–メス),(3) 採集時にオスにガードさ れていた未抱卵メス(G–メス).ランダムに選んだ 単独オスを水深3.5 cm程度の海水を入れたプラス チックコンテナ(16×12×8 cm)に入れて,その後 で3タイプのいずれかのメスを,オスとメスが互い に対角線上に位置するように入れた.デジタルカメ ラ (DMC-LF1, LUMIX) の録画機能を用いて30分間 の録画を行った.本研究では全ての個体を1度だけ 実験に用いた.実験の例数はOV–メス,NG–メス, G–メスについて,それぞれ49例,50例,51例であっ た. 動画再生による行動観察では (1) オスがメスに接 近し始めてから接触する直前まで (接近時),(2) オ スが接触後にメスを貝殻ごと仰向けにひっくり返し て貝殻の開口部を歩脚や鋏脚,触角で触り始めてか ら(あるいは,メスはうつぶせのままだがオスが貝 殻の開口部付近を触り始めてから),ガードや交尾が 始まるまで,あるいは一方の個体が離れるまで(評 価中),(3) オスがメスの貝殻を左鋏脚で掴み始めて から3分間(ガード中)の3場面でメスの行動を記 録した.メスをコンテナに入れてから10分以内に 評価やガードが見られなかった場合には,その場面 のメスの行動は観察されなかったものとして,例数 から除いた.また,接近時において,メスをコンテ ナに入れてからオスがメスに接近・接触するまでの 間,メスが全く貝殻から出てこない場合,もしくは オスの接近に気付いておらずオスに対して反応しな かった場合と,評価中・ガード中にメスの行動が不 明だった場合も例数から除いた.評価中は,評価開 始から終了までの時間がメスの行動に影響を与えて いる可能性があるため,これを評価時間(秒)とし て最長600秒間記録した.各場面のメスの行動を Table 1に示した.メスの行動をメスの反応として受 容と拒絶の2カテゴリーに分けた.ただし,接近時に は受容とも拒絶ともとれない行動が見られたため,反 応は中立を含めた3カテゴリーに分けた (Table 1). 動画撮影終了後に,G–メスを野外で採集した際に そのメスをガードしていたオスが入っている小型コ ンテナに戻したところ,すべてのメスがオスにガー ドされた.そこで,それらのガードペアを小型コン テナで飼育して,メスが脱皮したか否かを12時間 間隔で確認し,実験日からメスが脱皮した日までの 日数を0.5日単位で記録した.また,実体顕微鏡下 で対物ミクロメーターを使用し,実験に用いた全個 体の盾長(SL, mm)を体サイズの指標として測定 した.ただし,実験終了から測定までに脱皮し,脱 皮殻が残っていた場合は脱皮殻の盾長を測定した. 実験に用いた全ての単独オスの平均盾長は4.60 mm (±0.90SD),メスの平均盾長は4.16 mm (±0.92) で あった. メスの反応の頻度を3タイプ間で比較するために, Fisherの正確確率検定(Fisher test)を用い,有意差
Table 1. Ethogram of female behavior in the hermit crab Pagurus nigrofascia. Female behavior is categorized into three
responses: Acceptance, Neutral and Rejection.
Phase Response Behavior Description
Approaching Acceptance None No distinguishable behavior when a male approaches Neutral Withdraw Withdraw into her shell when a male approaches Rejection Retreat Retreat by slowly backing away from a male
Escape Rapidly escape from a male by active use of own walking legs Assessing
or Guarding Acceptance Tightly withdrawNo aggressive response Her body or appendage(s) is partially out but no visible movementTightly withdraw into her shell during male assessment or guarding Rejection Struggle Struggle by vigorous use of cheliped(s) and/or legs against a male
が認められた場合にはHolm法による事後検定を行っ た.また,メスの反応に影響を与える要因を検証す るために,多項分布あるいは二項分布を誤差分布に 指定した一般化線形モデル (generalized linear model, GLM) による検定を行った.接近時の解析では,3カ テゴリーのメスの反応 (中立:0,受容:1,拒絶:2) を応答変数として,(1)メスのタイプ(OV–メス, NG–メス,G–メス),(2) オスの体サイズ,(3) メス の体サイズを説明変数とした多項ロジスティック回 帰分析を行った.応答変数のベースラインは中立に 設定した.なお,G–メスの反応に対しては上記の説 明変数に(4) G–メスの交尾直前脱皮までの日数を 加えて,多項ロジスティック回帰分析を行った.評 価中とガード中の解析では2カテゴリーのメスの反 応 (受容:0,拒絶:1) を応答変数とし,説明変数に 上述のモデルと同じ3変数を用いたロジスティック 回帰分析を行った.G–メスの反応に対しても同様 に4つの説明変数を用いて解析した.なお,評価中 のメスの反応に関する解析では各モデルの説明変数 に(5)評価時間を加えた.本研究におけるすべて の統計解析はフリーソフトウェアR(version 3.3.2) (R Core Team, 2017) を用いて行い,多項ロジスティッ ク回帰分析にはパッケージmlogit (Croissant, 2013) を使用した. 結 果 接近時にメスがオスに示した反応は3タイプ (OV– メス,NG–メス,G–メス) 間で有意に異なり (Fisher test; P<0.001; Fig. 1),2タイプ間で個別に検定した 多重比較でも有意に異なった(Holm法,全ての群 間でP=0.012).拒絶行動の頻度はOV–メスで最も 高く,G–メスで最も低かった (Fig. 1).受容行動の 頻度はG–メスで最も高く,OV–メスで最も低い結 果となり,NG–メスでは中立行動の頻度が高かった (Fig. 1).中立行動に対する受容行動の起こりやすさ はNG–メスよりもG–メスで有意に高く,拒絶行動の 起こりやすさはNG–メスよりもOV–メスで有意に高 かった (Table 2).メスの反応は雌雄の体サイズによ り影響されなかった (Table 2).また,G–メスの反応 についても,雌雄の体サイズと交尾直前脱皮までの時 間による有意な影響は認められなかった (Table 3). 評価中にメスがオスに示した反応は3タイプ間で
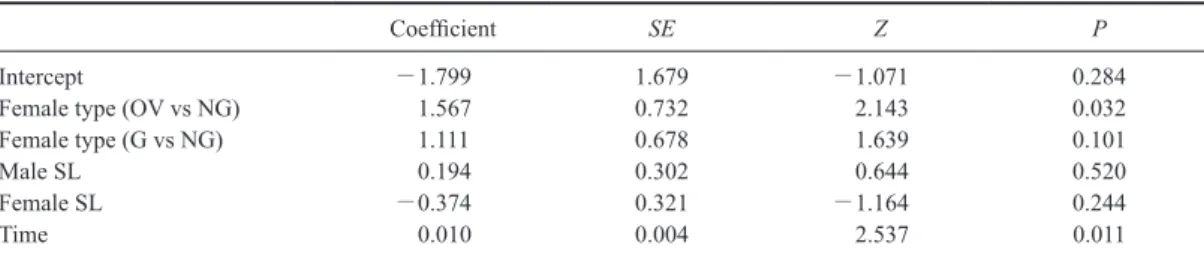
有意に異なった(Fisher test; P=0.016; Fig. 2).メス
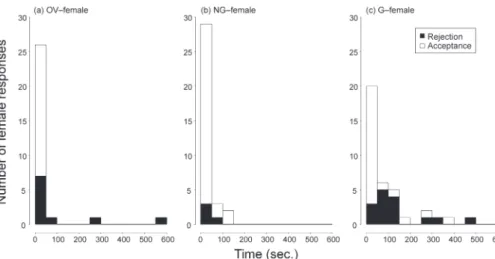
が拒絶行動を示した頻度はNG–メスとG–メス間で 有意に異なり(Holm法;P=0.041),G–メスで最も 高く,NG–メスで最も低かった (Fig. 2).一方,OV– メスと他2タイプの間に有意差は見られなかった (Holm法;P>0.1).評価中のメスの反応には評価時 間が有意な影響を与えており,評価時間が短いほど受 容行動の頻度が高く,長いほど拒絶行動の頻度が高 かった (Table 4).OV–メスは他2タイプと比べ評価 時間が短いときに拒絶行動の頻度が高く (Fig. 3),NG– メスとOV–メスの間に有意差が認められた (Table 4). メスの反応及びG–メスの反応は,接近時と同様に, 雌雄の体サイズ(Table 4, 5)及び交尾直前脱皮まで の日数(Table 5)とは独立であった.また,G–メ スの反応は,評価時間の影響を受けていなかった (Table 5). オスによる交尾前ガードは,OV–メスで1例,NG– メスで3例,G–メスで33例観察された (Fig. 4).G–メ
Fig. 1. Frequency of female response to a male during
the approach phase in the hermit crab Pagurus nigrofascia at Kattoshi, Hakodate Bay. Types of females were ovigerous (OV–female), females which had not been guarded in the field (NG–female), and females which had been guarded by other males in the field (G–female). Lower-case letters (a, b, c) across the top of the graph indicate which sets of conditions resulted in significant differences (different letters) in the frequency of female response.
スでは全体の約6割がガードに至ったが,メスの半数 以上がガード中にオスに対して拒絶を示した (Fig. 4). G–メスの反応に,雌雄の体サイズや交尾直前脱皮 までの時間の影響は認められなかった(Table 6). 考 察 ヨモギホンヤドカリのメスは,オスが交尾前ガー ド行動を開始する前後で逃走や抵抗などの拒絶行動 を示した.これは,本種の雌雄間で交尾前ガード行 動に起因する性的対立が生じていることを示唆する. オスの交尾前ガードに伴ってメスが拒絶行動を示す ことは,ホンヤドカリ属では今まで報告されていな かった.ホンヤドカリP. filholiやテナガホンヤドカリ P. middendorffiiでは体サイズの性差が大きいために, メスは,オスの交尾前ガード行動に対してほとんど 拒絶できないとみなされている(Wada et al., 1999;
Okamura & Goshima, 2010; Kawaminami & Goshima, 2015).これらの2種に比べて,ヨモギホンヤドカリ では体サイズの性差が小さいため(吉井ら,2009; 小島ら,2016),メスの拒絶行動が顕著なのかもし れない.ただし,ホンヤドカリとテナガホンヤドカ リでは,これまでにメスの拒絶行動に焦点を当てた 研究が行われたことはない.これらの2種を含むホ ンヤドカリ属の他種でも,メスの行動を精査すれ ば,拒絶行動が検出されるかもしれない. 交尾前ガードに関するメスの拒絶行動を説明する 仮説には,ガードコスト回避仮説とメスによるオス 選択仮説がある.メスはオスにガードされることで
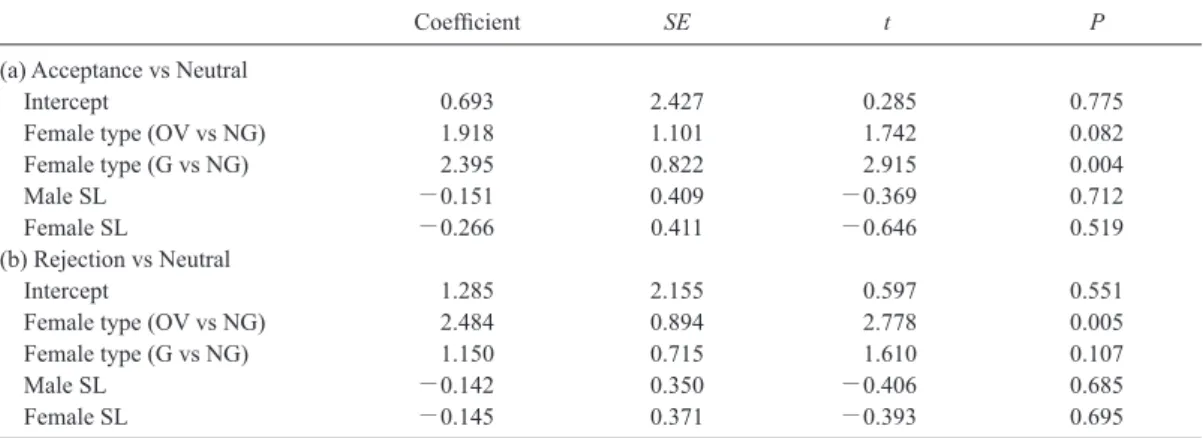
Table 2. Results of multinomial logistic regression analysis on female response during the approach phase. Types of females
were ovigerous females (OV–female), females which had not been guarded in the field (NG–female), and females guarded by other males in the field (G–female). NG females was set as baseline of the regression to compare responses among female types (i.e., OV vs NG, and G vs NG). “Neutral” response (see Table 1) was set as baseline to compare whether and how the other two responses, “Acceptance” and “Rejection”, differed from the baseline. Multinomial logistic regression analysis was applied to examine the effects of female group and shield length (SL, index of body size) of both sexes on female response during the approach phase.
Coefficient SE t P
(a) Acceptance vs Neutral
Intercept 0.693 2.427 0.285 0.775
Female type (OV vs NG) 1.918 1.101 1.742 0.082
Female type (G vs NG) 2.395 0.822 2.915 0.004
Male SL -0.151 0.409 -0.369 0.712
Female SL -0.266 0.411 -0.646 0.519
(b) Rejection vs Neutral
Intercept 1.285 2.155 0.597 0.551
Female type (OV vs NG) 2.484 0.894 2.778 0.005
Female type (G vs NG) 1.150 0.715 1.610 0.107
Male SL -0.142 0.350 -0.406 0.685
Female SL -0.145 0.371 -0.393 0.695
Table 3. Results of multinomial logistic regression analysis
on female response in G–females during the approach phase. “Neutral” response (see Table 1) was set as baseline to compare whether and how the other two responses, “Acceptance” and “Rejection” differed from the baseline. Multinomial logistic regression analysis was applied to examine the effects of shield length (SL, index of body size) of both sexes and the number of days until female prenuptial molting (Day) on female response in G–females during the approach phase.
Coefficient SE t P
(a) Acceptance vs Neutral
Intercept 2.924 4.013 0.729 0.466 Male SL -0.879 0.820 -1.072 0.284 Female SL 0.767 0.684 1.121 0.262 Day -0.184 0.533 -0.345 0.730 (b) Rejection vs Neutral Intercept 5.329 4.197 1.270 0.204 Male SL -1.473 0.896 -1.645 0.100 Female SL 0.841 0.720 1.168 0.243 Day -0.287 0.567 -0.506 0.613
摂餌機会の減少,捕食リスクの増加,オス間闘争に 巻き込まれることによる身体的損傷など様々なコス ト を 受 け る 可 能 性 が あ る(Jormalainen et al., 2001; Cothran, 2004; Takeshita et al., 2011).よって,メス はガードそのものを回避するため,もしくはガード 時間を短くするために拒絶を示すという説がガード コスト回避仮説である (Jormalainen et al., 2001).こ の仮説が正しい場合,交尾間近のメスほどガードに より受けるコストは小さくなり,いつまでもオスを 拒絶していると交尾機会自体を失う恐れがあるため, メスは交尾・産卵が近づくにつれて抵抗しなくなる と考えられている.実際に,ヘラムシ類のIdotea balticaのメスでは,交尾の日が近づくほどオスに対す る抵抗を弱めたことが報告されている(Jormalainen et al., 2000). 一方,本種では交尾直前脱皮までの日数がG–メス の反応に有意な影響を与えていなかった.しかし, 接近時に拒絶行動を示したメスの頻度はOV–メスで 最も高く,G–メスで最も低かった.本種のメスは 1年に一度しか産卵しないため (Goshima et al., 1996), 次の交尾機会まで約1年あるOV–メスは,長時間の ガードで受けるコストを避けるために拒絶行動を示 したと考えられる.一方,評価中及びガード中に拒 絶行動を示したメスの頻度は次の交尾機会までの時 間に依存して変化することはなく,交尾間近なG–メ スでも評価中は4割程度,ガード中は7割近くが拒 絶行動を示した.しかし,ごく一部の種を除けば (e.g. マルエラワレカラCaprella penantis; Takeshita et
al., 2011),甲殻類のメスが交尾前ガードによって負 うコストを実証した研究は非常に少なく,本種のメ スでもガードが大きなコストになることが実証され ているわけではない.よって,評価とガードの2場 面では,メスはガードコストの回避ではなく別の理 由から拒絶行動を示しているのかもしれない. ガードコスト回避仮説に対して,質の良いオス, もしくはメスの抵抗を打ち負かすオスを選択すると いう仮説がメスによるオス選択仮説である (Ridley &
Thompson, 1979; Sparkes et al., 2002).甲殻類では,
Fig. 2. Frequency of female response to a male during
the assessment phase. Types of females were ovigerous (OV–female), females which had not been guarded in the field (NG–female), and females which had been guarded by other males in the field (G–female). Lower-case letters (a, b) across the top of the graph indicate which sets of conditions resulted in significant differences (different letters) and no significant differences (same letters) in the frequency of female response.
Table 4. Results of logistic regression analysis on female response during the assessment phase. Logistic regression analysis
was applied to examine the effect of female group, shield length (SL, index of body size) of both sexes and assessment duration by a male (Time) on female response during the assessment phase (Acceptance or Rejection). Types of females were ovigerous females (OV–female), females which had not been guarded in the field (NG–female), and females guarded by other males in the field (G–female).
Coefficient SE Z P
Intercept -1.799 1.679 -1.071 0.284
Female type (OV vs NG) 1.567 0.732 2.143 0.032
Female type (G vs NG) 1.111 0.678 1.639 0.101
Male SL 0.194 0.302 0.644 0.520
Female SL -0.374 0.321 -1.164 0.244
オスの体サイズや精子保有量がメスの繁殖成功度に
影響を与えることが知られている(Sato & Goshima,
2007; Miura & Goshima, 2016).よって,この仮説が正 しければ,メスは次の交尾機会までの期間に関係な く,オスの質によって反応を変えると考えられる. Miura & Goshima (2016) は,イソヘラムシCleantiella isopusにおいて,メスが小型のオス,もしくは交尾 経験回数の多いオスほど拒絶することを明らかに し,本種においてメスによるオス選択仮説が支持さ れることを報告している.しかし,ヨモギホンヤド カリではメスの拒絶行動に対して雌雄の体サイズは 有意な影響を与えていなかったため,メスの拒絶行 動は少なくとも大型オスを選択するためではないと 考えられる. ホンヤドカリのメスは交尾前ガード中にも性フェ
Fig. 3. Relationships between the assessment duration by a male (Time, seconds) and the response of each female during
the assessment phase (Acceptance or Rejection). Types of females were (a) ovigerous (OV–female), (b) females which had not been guarded in the field (NG–female), and (c) females which had been guarded by other males in the field (G–female).
Fig. 4. Number of female response to a male during
the guarding phase. Types of females were ovigerous (OV–female), females which had not been guarded in the field (NG–female), and females which had been guarded by other males in the field (G–female).
Table 5. Results of logistic regression analysis on female
response in G females during the assessment phase. Logistic regression analysis was applied to examine the effect of shield length (SL, index of body size) of both sexes, assessment duration by a male (Time) and the number of days until female prenuptial molting (Day) on female response in G–females during the assessment phase (Acceptance or Rejection). Coefficient SE Z P Intercept 1.269 2.472 0.513 0.608 Male SL -0.207 0.549 -0.376 0.707 Female SL -0.298 0.462 -0.645 0.519 Time 0.007 0.004 1.598 0.110 Day -0.133 0.290 -0.457 0.648
ロモンを放出して他のオスを惹き付け,オス間闘争 を誘発する.その結果,メスは闘争に強いオスと交 尾をする (Okamura & Goshima, 2010).このような例 は間接的な配偶者選択と呼ばれている.様々な分類 群で,メスが性フェロモンを用いてオスを誘引する ことが知られている.ブルークラブCallinectes sapidus やクリガニTelmessus cheiragonusでは,メスの尿中に 含まれる脱皮バイオマーカーNAGLの濃度が交尾可 能になるまでの期間が短くなるにつれて高くなるこ とから,本物質が性フェロモンとして機能すると考 えられている(Kamio et al., 2014; Yano et al., 2016). ヨモギホンヤドカリのメスは,ブルークラブ,クリ ガニと同様に交尾前に必ず脱皮するため(交尾直前 脱皮;Suzuki et al., 2012),同じような化学キュー を放出している可能性がある.本研究では,評価中 やガード中に観察された本種のメスの行動のほとん どが,歩脚や鋏脚を振りまわして暴れるような行動 であったため,これらの行動をメスによる拒絶の反 応とみなした.しかし,この行動はオスを拒絶する ためだけではなく,性フェロモンを拡散するという 機能も持ち合わせる行動なのかもしれない.また, Yamanoi et al. (2006)はホンヤドカリのメスが交尾を 遅らせることによりオス間闘争を多く引き起こすこ とを示した.本研究では,ガード中のオスが交尾を 試みようとメスを貝から引きずり出すものの,メス が歩脚や鋏脚を激しく動かしたため交尾に至らない 場面がしばしば観察された(木戸,個人的観察). このように,ガード中のメスの拒絶行動は交尾のタ イミングを遅らせる機能があるのかもしれない. 本研究から本種の雌雄間に性的対立が存在するこ とが示唆された.メスが拒絶行動によって得る利益 について検証を行うと同時に,本種のメスがガード によって受けるコストを検証することで,メスの拒 絶行動についてより詳細な考察ができるだろう.ま た,メスの拒絶行動に対するオスの行動を含めた雌 雄の相互作用に着目した研究を行うことも,本種に 生じる性的対立を解明するうえで重要な課題だと考 えられる. 謝 辞 本研究を進めるにあたり,多くの助言と協力をい ただきました北海道大学大学院水産科学研究院海洋 生物学講座動物生態学研究室の皆様に心より感謝申 し上げます.また,本稿に対して多くの有益なご助言 をいただいた査読者にも厚くお礼申し上げます.な お,本研究の一部は,科学研究費助成金 (17K1518807) で実施された. 文 献
Aloise King, E. D., Banks, P. B., & Brooks, R. C., 2013. Sexual conflict in mammals: consequences for mating systems and life history. Mammal Review, 43: 47–58. Cothran, R. D., 2004. Precopulatory mate guarding affects
predation risk in two freshwater amphipod species. Ani-mal Behaviour, 68: 1133–1138.
Croissant, Y., 2013. mlogit: multinomial logit model. R package version 0. 2–4. http://CRAN.R-project.org/package= mlogit.
Goshima, S., Wada, S., & Ohmori, H., 1996. Reproductive biology of the hermit crab Pagurus nigrofascia (Ano-mura: Paguridae). Crustacean Research, 25: 86–92. Grafen, A., & Ridley, M., 1983. A model of mate guarding.
Journal of Theoretical Biology, 102: 549–567.
Hazlett, B. A., 1966. Social behavior of the Paguridae and Diogenidae of Curaçao. Studies on the Fauna of Cura-çao and Other Caribbean Islands, 88: 1–143.
Imafuku, M., 1986. Sexual discrimination in the hermit crab Pagurus geminus. Journal of Ethology, 4: 39–47. Jivoff, P., & Hines, A. H., 1998. Female behavior, sexual
competition and mate guarding in the blue crab, Calli-nectes sapidus. Animal Behaviour, 55: 589–603. Jormalainen, V., 1998. Precopulatory mate guarding in
crus-taceans: Male competitive strategy and intersexual con-flict. The Quarterly Review of Biology, 73: 275–304. Jormalainen, V., & Merilaita, S., 1995. Female resistance
Table 6. Results of logistic regression analysis on response
in G females during the guarding phase. Logistic regression analysis was applied to examine the effect of shield length (SL, index of body size) of both sexes and the number of days until female prenuptial molting (Day) on response in G– females during the guarding phase (Acceptance or Rejection). Coefficient SE Z P Intercept 0.764 2.249 0.340 0.734 Male SL -0.370 0.472 -0.783 0.434 Female SL 0.470 0.441 1.065 0.287 Day -0.116 0.303 -0.384 0.701
and duration of mate-guarding in three aquatic peraca-rids (Crustacea). Behavioral Ecology and Sociobiology, 36: 43–48.
Jormalainen, V., Merilaita, S., & Härdling, R., 2000. Dynam-ics of intersexual conflict over precopulatory mate guarding in two populations of the isopod Idotea balti-ca. Animal Behaviour, 60: 85–93.
Jormalainen, V., Merilaita, S., & Riihimäki, J., 2001. Costs of intersexual conflict in the isopod Idotea baltica. Journal of Evolutionary Biology, 14: 763–772. Jormalainen, V., & Shuster, S. M., 1999. Female
reproduc-tive cycle and sexual conflict over precopulatory mate-guarding in Thermosphaeroma (Crustacea, Isopoda). Ethology, 105: 233–246.
Kamio, M., Schmidt, M., Germann, M. W., Kubanek, J., & Derby, C. D., 2014. The smell of moulting: N-acetyl-glucosamino-1,5-lactone is a premoult biomarker and candidate component of the courtship pheromone in the urine of the blue crab, Callinectes sapidus. The Journal of Experimental Biology, 217: 1286–1296.
Kawaminami, T., & Goshima, S., 2015. Guarding males of the hermit crab climb up algae to avoid male-male competition. Journal of Ethology, 33: 25–33.
小島早智・石原(安田)千晶・和田哲,2016.ヨモギホ
ンヤドカリの交尾前ガードペアにみられる同類交
配と闘争経験に基づいたオスの配偶者選択.
Can-cer, 25: 17–24.
Magurran, A. E., & Seghers, B. H., 1994. Sexual conflict as a consequence of ecology: evidence from guppy, Poe-cilia reticulata, populations in Trinidad. Proceedings of the Royal Society of London, Series B, 255: 31–36. Minouchi, S., & Goshima, S., 2000. The effect of male size
and sex ratio on the duration of precopulatory mate guarding in the hermit crab Pagurus filholi. Benthos Research, 55: 37–41.
Miura, Y., & Goshima, S., 2016. Temporal dynamics of in-tersexual conflict and the effect of male quality on fe-male fecundity in the marine isopod Cleantiella isopus. Behaviour, 153: 569–589.
Okamura, S., & Goshima, S., 2010. Indirect female choice mediated by sex pheromones in the hermit crab Pagu-rus filholi. Journal of Ethology, 28: 323–329.
Parker, G. A., 1979. Sexual Selection and Reproductive Competition in Insects. In: M.S. Blum, & N.A. Blum (eds.), Sexual selection and sexual conflict. Academic Press, New York, pp. 123–166.
R Core Team, 2017. R: A language and environment for sta-tistical computing. R Foundation for Stasta-tistical Com-puting, Vienna, Austria. URL http://www.R-project. org/.
Ridley, M., & Thompson, D. J., 1979. Size and mating in Asellus aquaticus (Crustacea: Isopoda). Zeitschrift für Tierpsycholodie, 51: 380–397.
Rowe, L., Arnqvist, G., Sih, A., & Krupa, J. J., 1994. Sexual
conflict and the evolutionary ecology of mating pat-terns: water striders as a model system. Trends in Ecol-ogy & Evolution, 9: 289–293.
Sato, T., & Goshima, S., 2007. Female choice in response to risk of sperm limitation by the stone crab, Hapalogaster dentata. Animal Behaviour, 73: 331–338.
Shine, R., O’Connor, D., & Mason, R. T., 2000. Sexual con-flict in the snake den. Behavioral Ecology and Sociobi-ology, 48: 392–401.
Sparkes, T. C., Keogh, D. P., & Orsburn, T. H., 2002. Female resistance and mating outcomes in a stream-dwelling isopod: effects of male energy reserves and mating his-tory. Behaviour, 139: 875–895.
Suzuki, Y., Yasuda, C., Takeshita, F., & Wada, S., 2012. Male mate choice and male-male competition in the hermit crab Pagurus nigrofascia: importance of female quality. Marine Biology, 159: 1991–1996.
Szentirmai, I., Székely, T., & Komdeur, J., 2007. Sexual con-flict over care: antagonistic effects of clutch desertion on reproductive success of male and female penduline tits. Journal of Evolutionary Biology, 20: 1739–1744. Takeshita, F., & Henmi, Y., 2010. The effects of body size,
ownership and sex-ratio on the precopulatory mate guarding of Caprella penantis (Crustacea: Amphipoda). Journal of the Marine Biological Association of the United Kingdom, 90: 275–279.
Takeshita, F., Lombardo, R. C., Wada, S., & Henmi, Y., 2011. Increased guarding duration reduces growth and offspring number in females of the skeleton shrimp Ca-prella penantis. Animal Behaviour, 81: 661–666. Wada, S., Tanaka, K., & Goshima, S., 1999. Precopulatory
mate guarding in the hermit crab Pagurus middendorffii (Brandt) (Decapoda: Paguridae): effects of population parameters on male guarding duration. Journal of Ex-perimental Marine Biology and Ecology, 239: 289–298. Yamamura, N., 1987. A model on correlation between pre-copulatory guarding and short receptivity to copulation. Journal of Theoretical Biology, 127: 171–180.
Yamamura, N., & Jormalainen, V., 1996. Compromised strategy resolves intersexual conflict over pre-copulato-ry guarding duration. Evolutionapre-copulato-ry Ecology, 10: 661– 680.
Yamanoi, T., Yoshino, K., Kon, K., & Goshima, S., 2006. Delayed copulation as a means of female choice by the hermit crab Pagurus filholi. Journal of Ethology, 24: 213–218.
Yano, H., Kamio, M., & Nagai, H., 2016. The molting bio-marker metabolite N-acethlglucosamino-1,5-lactone in female urine of the helmet crab Telmessus cheiragonus. The Biological Bulletin, 230: 143–151.
吉井健二・竹下文雄・和田哲,2009.ヨモギホンヤド
カリのオスにおける繁殖と成長:交尾前ガード行