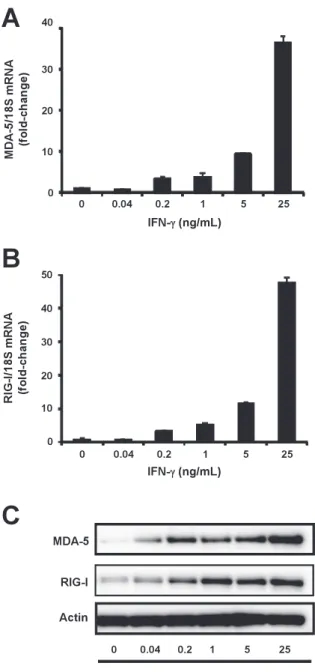
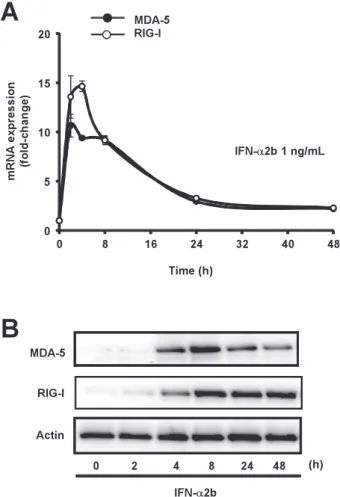
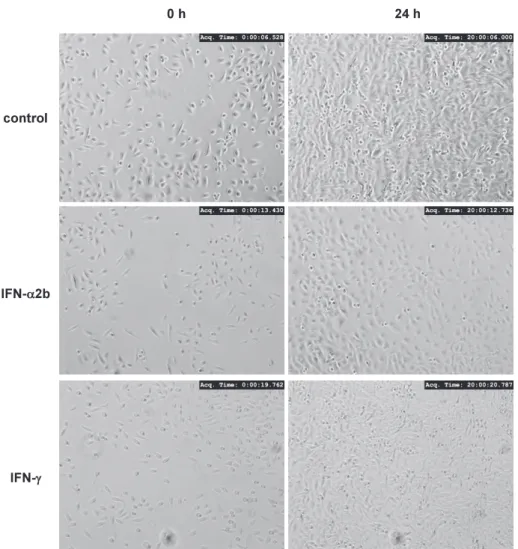
Role of type I- and type II-interferon in expression of melanoma differentia- tion-associated gene-5 in HSC-3 oral squamous carcinoma cells
Takao K ON
1, Tomoh M ATSUMIYA
2, Ryo H AYAKARI
2, Norihiko N ARITA
1, Ryohei I TO
1, Kosei K UBOTA
1, Hirotaka S AKAKI
1, Hidemi Y OSHIDA
2, Tadaatsu I MAIZUMI
2, Wataru K OBAYASHI
1, and Hiroto K IMURA
11