Characterization of Brevilysin H2, an α-Fibrinogenolytic Metalloproteinase from the Venom of Chinese Snake (Gloydius blomhoffi brevicaudus)
Shigeyuki Terada1) and Takahiro HaTTori1)
(Received November 30, 2012)
Abstract
Brevilysin H2 is a non-hemorrhagic metalloproteinase isolated from the venom of Chinese snake (Gloydius blomhoffi brevicaudus). The molecular mass of H2 was 69 kDa on SDS-PAGE, and decreased to 52 kDa after deglycosylation. The enzyme has the optimum pH of 9.5, and is stable in the pH range of 5.5–13.0 and below 50 °C. The enzyme is fully active even in 4 M urea. The activity of H2 did not change after a prolonged incubation at pH 7.0 and 37 °C. Although EDTA and some thiol compounds inhibited the enzyme, alkaline earth metal ions (Mg2+, Ca2+, Sr2+, and Ba2+) significantly enhanced the proteolytic activity and the thermal stability. H2 preferentially cleaved X-Leu bond, and showed an α-fibrinogenase activity. The complete amino acid sequence of H2 was determined by Edman degradation. H2 is a P-III class metalloproteinase composed of 424 amino acids with an N-terminal pyroglutamate residue. Sequence heterogeneity was found exclusively in the cysteine-rich domain of brevilysin H2, showing the presence of isozymes.
Introduction
Snakes belonging to the Viperidae family have hemorrhagic venoms that contain various biologically active substances [1]. Some of them affect blood coagulation, fibrinolytic system, and blood pressure.
Others cause cytotoxicity, hemolysis, tissue necrosis, neurotoxicity and hemorrhage. Metalloproteinases existing in snake venoms are the principal substance of hemorrhagic factors.
Snake venom metalloproteinases (SVMPs) belong to the reprolysin subfamily among several zinc-containing metalloproteinases. They are
characterized by the presence of a conservative zinc chelating sequence HEXXHXXGXXHD as an essential part for the proteolytic activity and a structurally conserved methionine-containing turn [2]. The different molecular sizes of SVMPs reflect the numbers of domains: metalloproteinase (M), disintegrin-like (D), and cysteine-rich (C) domains [3]. Snake venom reprolysins are divided into several classes (P-I to P-III) based on the domain structure of the proteins and genes [4]. The P-I class comprises a low molecular mass enzyme that is composed solely of an M domain. The intermediate type proteins (class P-II) have an additional D domain, which is
1) Department of Chemistry, Faculty of Science, Fukuoka University, 8-19-1 Nanakuma, Jonan-ku, Fukuoka 814-0180, Japan.
Abbreviations: Arg-C, arginylendopeptidase; Asp-N, endoproteinase Asp-N; CAPSO, N-Cyclohexyl-3-aminopropanesulfonic acid; C, cysteine-rich; D, disintegrin-like; FTC, fluorescein thiocarbamoyl; H2, brevilysin H2; HPLC, high-performance liquid chromatography; HVR, hyper-variable region; Lys-C, Achromobacter protease I; M, metalloproteinase; PE, S-pyridylethylated; SVMP, snake venom metalloproteinase; TES, N-tris(hydroxymethyl)methyl-2-aminoethanesulfonic acid.
homologous to those of platelet aggregation inhibitors (disintegrins). High molecular mass proteases (class P-III) consist of the all three domains (the MDC domain), and some have additional domains at the C-terminal.
Chinese viper (G. blomhoffi brevicaudus) venom contains a variety of toxic proteins including some hemorrhagic and non-hemorrhagic metalloproteinases.
Previously, we have isolated eight SVMPs named brevilysins H1–H6, L4, and L6 with different substrate specificity from the G. blomhoffi brevicaudus venom [5]. Although brevilysin H3, H4, and H6 are 60-80 kDa proteins with hemorrhagic activity, H3 and H4 are the major hemorrhagins. On the other hand, brevilysins H1, H2, L4, and L6 show no hemorrhagic activity. The amino acid sequences of two P-I class proteinases brevilysins L4 [6] and L6 [7] and two P-III class enzymes termed brevilysins H1 [8] and H6 [9] have already reported. In order to correlate the substrate specificity and hemorrhagic activity with their primary structures, we intended to characterize these enzymes. Here, we report the property and amino acid sequence of brevilysin H2, a non-hemorrhagic metalloproteinase in the same venom.
Experimental
Materials
Crude G. blomhoffi brevicaudus venom preserved by lyophilization was used. Brevilysin H2 was prepared as described [5]. Fluorescein thiocarbamoyl (FTC)-casein was prepared according to Twining [10]. Bovine insulin, Achromobacter protease I (Lys-C), Glycopeptidase F, arginylendopeptidase (Arg-C), pyroglutamate aminopeptidase from porcine liver, and endopeptidase Asp-N (Asp-N) were purchased from Takara (Kyoto). N-Cyclohexyl- 3-aminopropanesulfonic acid (CAPSO) and N-tris(hydroxymethyl)methyl-2- aminoethanesulfonic acid (TES) are products of Dojindo (Kumamoto).
Performic acid-oxidized insulin B-chain was prepared according to Sanger [11]. Human fibrinogen (plasminogen-free) was from Nacalai tesque (Kyoto).
All other chemicals were purchased from Wako Pure Chem. Inc. (Osaka).
Measurement of protease activity
Protease activity was measured using FTC-
casein as the substrate in 50 mM Gly-NaOH buffer (pH 9.5) at 37 °C [9]. The increase in fluorescence was measured with a FP-750A spectrofluorometer (Jasco) at 520 nm with the excitation at 490 nm.
Deglycosylation by glycopeptidase F
Purified H2 (25 µg) was dissolved in 5 µL of 0.75
% 2-mercaptoethanol, 0.5 %SDS, 0.5 M Tris-HCl buffer (pH 8.6) and the mixture was heated at 100 °C for 3 min. Then, 5 µL of 5 % Nonidet P- 40 was added. After the addition of 15 µL of glycopeptidase F (1 mU), the reaction mixture was kept at 37 °C for 20 h.
Stability of enzyme to pH, heating and urea
The pH stability of H2 was examined in various buffers: 20 mM Gly-HCl (pH 2.0-4.0), 20 mM TES- NaOH (pH 4.5-8.0), or 20 mM Gly-NaOH (pH 8.5-13.0).
After 60 µL of the enzyme solutions (127 µg/mL) had been kept at 37 °C for 30 min, 190 µL of 0.26 % FTC-casein in 195 mM CAPSO–NaOH buffer (pH 9.5) were added. The residual activity was measured as described above. Thermal stability was examined in 20 mM TES-NaOH buffer (pH 7.0). The enzyme solution (150 µg/mL) was heated at 40-80 °C for 30 min in a water bath and then cooled in ice. Thermal stability was evaluated in terms of a T50, a temperature at which the enzymes activity reduced by half. After enzyme solutions (150 µg/mL) in 50 mM Gly-NaOH buffer (pH 9.5) containing 0-4 M urea were pre- incubated at 25 °C for 5 min, the residual activity was measured. To evaluate the effects of Ca2+ on the stability of enzyme, 2.5 mM CaCl2 was added in the pre-incubation mixture.
Effect of the prolonged incubation on the proteolytic activity
H2 (0.15 mg/mL) was dissolved in 50 mM Gly- NaOH buffer (pH 9.5) and incubated at 37 °C for up to 4 h. Aliquots of the reaction mixture were withdrawn and the residual activity was measured as described above.
Hydrolysis of protein and peptide
Fibrinogenase activity was analyzed by SDS- PAGE under reducing conditions. Human fibrinogen (50 µg/mL) was treated with brevilysin H2 (10 µg/
mL) in 10 µL of 2 mM CaC12, 150 mM NaCl, 50 mM Tris-HC1 buffer (pH 7.4) at 37 °C. The reaction was stopped by the addition of 7.5 M urea, 4 % SDS, 4 % 2-mercaptoethanol (50 µL), and 10 µL of the mixture was subjected to SDS-PAGE analysis in 12 % polyacrylamide gel according to Laemmli [12].
Proteins were detected by staining gels with 0.1 % Coomassie brilliant blue R-250 and destained by 10 % acetic acid.
Performic acid-oxidized insulin (320 µg) was incubated with H2 (10 µg) at 37 °C for 20 h in 50 µL of 50 mM Tris-HCl buffer (pH 8.5). The resultant peptide fragments were separated by a reverse- phase high-performance liquid chromatography (HPLC) on a TSK-gel ODS-120T column (0.46 × 25 cm, Tosoh) with a linear gradient of acetonitrile in 0.1 % trifluoroacetic acid. The positions of cleavage sites were determined by the sequence of each fragment.
Sequence analysis
To remove a small quantity of impurities, brevilysin H2 was further purified by a reverse-phase HPLC on a Cosmosil C8-AR-300 column (0.46 × 15 cm, Nacalai tesque) with a convex gradient of acetonitrile in 0.1 % trifluoroacetic acid. Pure brevilysin H2 was then reduced and subjected to S-pyridylethylation according to Friedman et al. [13].
In brief, H2 (11.0 mg) was dissolved in 1.7 mL of 30 mM dithiothreitol, 6 M guanidine-HC1, 10 mM EDTA, 0.5 M Tris-HC1 buffer (pH 8.6), and the mixture was left at room temperature for 2.5 h under N2
substitution. 4-Vinylpyridine (28 µL) was added and the mixture was further incubated at room temperature in a dark for 2.5 h under N2 substitution.
After the reaction was terminated by the addition of small amount of acetic acid, the excess reagents were removed by a reverse-phase HPLC on Cosmosil 5C8- AR-300 column (0.46 × 15 cm), yielding 6.8 mg of S-pyridylethylated (PE)-brevilysin H2.
PE-H2 (0.9 mg) was digested with Lys-C (2.5 mU) in 100 mM Tris-HCl buffer (pH 9.0) at 37 °C for 4 h. PE-H2 (1.0 mg each) was also treated with Arg-C (20 µg) or Asp-N (20 µg) in 50 mM phosphate buffer (pH 8.0) at 37 °C for 20 h. The N-terminal pyroglutamic residue was removed by pyroglutamate aminopeptidase as described previously [8]. For chemical cleavage, PE-H2 (1.0 mg) was dissolved in 70 µL of 70 % formic acid and BrCN (2.35 mg) was added. After 24 h at room temperature, the reaction mixture was diluted with distilled water and lyophilized. Peptides derived from enzymatic or chemical cleavage were separated by a reserve- phage HPLC on an ODS-120T column (0.46 × 25 cm, Tosoh) with a linear gradient of acetonitrile in 0.1 %
trifluoroacetic acid. The amino acid sequences of peptides were determined using an automatic protein sequencing system PPSQ 21 (Shimadzu).
Results
Property of brevilysin H2
Brevilysin H2 was prepared as described [5].
The purified protein showed a single band on SDS- PAGE and the molecular mass was estimated to be 69 kDa under non-reducing conditions (data not shown). This agrees with the value in a previous report [5]. The molecular mass value decreased to 52 kDa after digestion with glycopeptidase F (data not shown), indicating that brevilysin H2 is a glycoprotein.
Proteolytic activity of the enzyme was measured in buffers of various pHs using FTC-casein as the substrate. The optimum pH of H2 was about 9.5 (Fig.
1A). Stability of the enzyme to various pHs was examined by measuring FTC-caseinolytic activity after treatment in buffers of various pH at 37 °C for 30 min. Brevilysin H2 was stable in the pH range of 5.5 to 13.0 (Fig. 1B). Thermal stability of H2 was tested in 20 mM TES-NaOH buffer (pH 7.0). The enzyme solution was heated at 40-80 °C for 30 min and the residual activity was measured. The activity decreased slowly at 55 °C and disappeared above 70
°C. The temperature (T50) at which the enzyme loses 50 % of activity was estimated to be approximately 56
°C under the above conditions (Fig. 2A). In the meanwhile, brevilysin H1 and H6 have T50 values of 64 and 47 °C, respectively [8, 9]. Addition of Ca2+
slightly enhanced the stability of H2 and raised the T50 value to 61 °C (Fig. 2A). Similar effect on the thermal stability was observed by Ba2+ ion (data not shown). Brevilysin H2 was rather stable for a prolonged incubation at 37 °C and pH 9.5 and its FTC- caseinolytic activity did not change so much even after 4 h (Fig. 2B).
Effect of urea, metal ions, thiol compounds, and several proteinase inhibitors
Protease activity of brevilysin H2 was fully maintained in the presence of 4 M urea (Fig. 3A).
Metalloproteinases are influenced by some metal ions and inactivated by metal chelating agents that can remove a zinc ion from the active site of enzymes. We examined the effects of several metal ions and metal chelating agents on the caseinolytic activity of H2.
The activity was decreased profoundly by some heavy metal ions such as Cu2+, Fe2+, and Hg2+, and weakly by Zn2+ (Table 1). On the other hand, all the alkaline earth metal ions enhanced the activity almost twice. When the enzyme was incubated in the presence of various concentration of MgC12, CaC12, SrC12, or BaC12, a dose-dependent increase of activity was observed (Fig. 3B). At 5 mM, these ions enhanced the activity up to 250 %. Ba2+ seemed to be the most effective among these ions. Effect of thiol compounds was also determined similarly. Dithiothreitol and L-cysteine were inhibitory to H2, but thioglycolic acid, cysteamine, and 2-mercaptoethanol gave no effect (Table 1).
Several inhibitors to serine and cysteine proteases were ineffective (Table 2). Low molecular mass inhibitors (chymostatin, leupeptin, and antipain) from bacterial origin were not effective either. On the contrary, metal chelating agents such as EDTA, TPEN, and 1,10-phenanthrolin strongly inhibited the enzyme, indicating that H2 is a metalloproteinase.
However, a synthetic metalloproteinase inhibitor Cl- Ac-(OH)Leu-Ala-Gly-NH2 [14] gave no effect on the activity of H2 even at 1 mM. We have already Fig. 1. (A) Optimum pH of brevilysin H2. Activity
was measured using 0.2 % FTC-casein in 40 mM Tricine-NaOH (pH 7.5-10.5). Enzyme concentration was 30 µg/mL. (B)Stability of H2 at several pHs. After the enzyme was incubated at 37 °C for 30 min in 20 mM Gly- HCl (pH 2-4, ○), 20 mM TES-NaOH (pH 4.5- 8.5, ▲), or 20 mM Gly-NaOH (pH 9-13.5, ■), the residual FTC-caseinolytic activity was measured in 148 mM CAPSO-NaOH (pH 9.5).
Fig. 2. (A) Thermal stability of brevilysin H2. The protease was incubated in 20 mM TES-NaOH (pH 7.0) at the indicated temperature for 30 min. The residual activity was measured in 50 mM Gly-NaOH (pH 9.5). Open and closed circles indicate the activity in the absense and presence of 2.5 mM CaCl2. Dotted lines indicate T50 values. (B) Stability of H2 on the prolonged incubation. H2 was dissolved in 100 mM Gly- NaOH (pH 9.5). After the mixture was incubated at 37 °C for several hours, the residual activity was measured using 0.2 % FTC-casein as the substrate. Enzyme concentration was 30 µg/mL.
reported that brevilysin H2 is not inhibited by MSF, a natural proteinous inhibitor of SVMP isolated from the serum of G. blomhoffi brevicaudus [15].
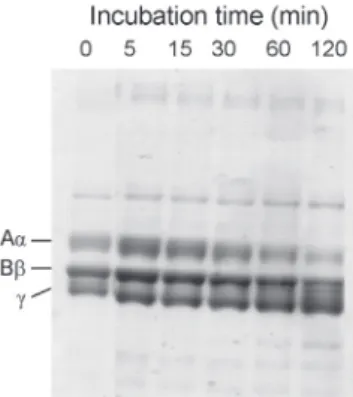
Action of H2 on protein and peptide substrates Fibrinogenolytic activity was determined by SDS-PAGE analysis. Human fibrinogen gave 3 bands corresponding to Aα (70 kDa), Bβ (60 kDa), and γ (52 kDa) chains on SDS-PAGE under reducing conditions. When human fibrinogen was incubated
with the enzyme at 37 °C for 5-120 min, the Aα chain was rapidly cleaved (Fig. 4). The Bβ chain was cleaved very slowly, while the γ chain was not affected at all. This feature is common to many SVMPs [16].
Oxidized insulin was digested with the enzyme at pH 8.5 and 37 °C for 20 h and the resultant peptides were separated by a reserve-phase HPLC. The positions of peptide bonds cleaved were identified by means of the amino acid sequencing. Brevilysin H2 degraded the B-chain of insulin at four peptide bonds, His5-Leu6, Ser9-His10, Ala14-Leu15, and Tyr16-Leu17 (Fig.
5). The cleavage sites by other brevilysins are also Table 1. Effect of metal ions and thiol compounds on
the FTC-caseinolytic activity of brevilysin H2.
Agents Concentration Relative
(mM) activity (%)
MgC12 1 164.7
CaC12 1 188.3
SrC12 1 177.2
BaC12 1 198.0
CuSO4 1 13.7
FeSO4 1 6.0
HgC12 1 0.0
ZnC12 1 22.0
L-Cysteine 1 26.7
Cysteamine 1 99.6
2-Mercaptoethanol 1 94.0
Dithiothreitol 1 0.0
Thioglycolic acid 1 81.3
Fig. 3. (A) Effect of urea on the proteolytic activity of protease H2. Activity was measured using 0-4 M urea and 0.2 % FTC-casein in 50 mM Gly-NaOH (pH 9.5). Enzyme concentration was 30 µg/mL. (B) Dose-dependent activation of H2 by alkaline earth metal ions. FTC- caseinolytic activity was measured in 50 mM CAPSO-NaOH (pH 9.5) in the presence of CaCl2 (○), MgCl2 (●), BaCl2 (▲), and SrCl2
(△).
Table 2. Effect of several proteinase inhibitors on the FTC-caseinolytic activity of brevilysin H2.
Agents Concentration Relative
(mM) activity(%) Serine protease inhibitors
Diisopropylfluorophosphate 0.01 60.5 Phenylmethanesulfonylfluoride 1 95.3
Chymostatin 1 96.1
Leupeptin 1 100.0
Cysteine protease inhibitors
N-Ethylmaleimide 1 104.9
p-Chloromercurybenzoic acid 1 72.9
Antipain 0.01 90.4
Metalloproteinase inhibitors
EDTA 1 15.9
1,10-Phenanthrolin 1 58.3
TPENa) 1 27.5
Cl-Ac-(OH)Leu-Ala-Gly-NH2b) 1 90.1
a) N,N,N',N'-Tetrakis(2-pyridylmethyl)ethylenediamine.
b) A synthetic inhibitor for bacterial metalloproteinase.
illustrated in Fig. 5. Preference of H2 to X-Leu bonds is noticeable.
Sequence Analysis of brevilysin H2
Because direct sequencing of the intact H2 and PE-H2 was also unsuccessful, the N-terminus of the protein might be blocked probably by a pyroglutamic residue. This cyclized residue does not have a free amino group, and therefore, the peptide or protein is not amenable to sequence determination, unless the pyroglutamyl derivative is removed. When the intact PE-H1 was sequenced after digestion with pyroglutamate aminopeptidase, we could not
determine the N-terminal sequence. PE-H2 was subjected to chemical cleavage at Met-X bonds with BrCN to give fragments M1-M8. Although Edman degradation of peptides M5 liberated no amino acid, the treatment of M5 with this enzyme gave a sequence of QRYLNAKR..., suggesting that the N-terminal of H2 is QQRYLNAKR... (Fig. 6)
After PE-brevilysin H2 was digested by several hydrolytic enzymes, the resulting peptides were separated by reverse-phase HPLC (data not shown).
Digestions by Lys-C, Arg-C, and Asp-N yielded peptides K1-K30, R1-R17, and D1-D28, respectively.
The arrangement of these fragments along with the complete amino acid sequence of H2 is summarized in Fig. 6. In the case of peptide K16 (E65DLIXVTSAAK75), no amino acid was detected at 5 cycle. The presence of a following sequence Val-Thr indicates that an N-linked sugar chain is attached to residue 69 (Asn- 69) of this peptide. Similarly, Asn-185 residue in peptides M7 (172-), K9 (182-188), and D10 (177-189) was glycosylated.
Brevilysin H2 was composed of 424 amino acids (Fig. 6). During sequencing experiment of H2, however, we noticed the sequence heterogeneity of the C-terminal region of this protein. The results of sequence analysis of the C-terminal parts are cited in detail (Fig. 7). Total 18 amino acid substitutions are present at residues 303-406. On the other hand, no heterogeneity was found in the preceding sequence (residues 1-302). This implies that our H2 preparation Fig. 4. SDS-PAGE analysis of human fibrinogen
digestion by brevilysin H2. Human fibrinogen (10 mg/mL) was digested with H2 (10 µg/
mL) in 2 mM CaCl2-150 mM NaCl-50 mM Tris- HC1 buffer (pH 7.4) at 37 °C. The reaction was terminated by adding 5 volumes of 7.5 M urea-4 % SDS-4 % 2-mercaptoethanol.
Fig. 5. Comparison of the substrate specificities of brevilysins. The cleavage sites of the oxidized bovine insulin B chain are indicated by arrows.
was the mixture of two similar proteins. The main and minor components are then designated to be H2a and H2b, respectively (Figs. 6 and 7).
Discussion
A 69-kDa metalloproteinase named brevilysin H2 was purified from G. blomhoffi brevicaudus venom, and the enzymatic property was first determined.
Brevilysin H2 contains Zn2+ as an essential metal ion for their catalytic activity. Thus, the treatment with metal cheating agents destroys the enzyme activity (Table 1). Enhancement of proteolytic activity by Ca2+ ion was often reported for several SVMPs [17, 18]. Proteolytic activity of H2 was influenced by all the alkaline metal ions including Ca2+ ion.
Approximately 250 % increase of the activity was observed at 5 mM. Thermal stability of H2 was slightly augmented by these ions. As reported previously [9], both proteolytic activity and thermal stability of H6 were also markedly enhanced by the addition of Ca2+ ion. On the contrary, Ca2+ ion had no effect on the activity and thermal stability of H1 and L6 [7, 8]. The effect of Ca2+ ion on brevilysin L4 was different. Although this ion enhanced the proteolytic activity, it was ineffective at raising its thermal stability.
Substrate specificity of SVMPs has been examined using B-chain of oxidized insulin as a common substrate. Most of SVMPs attack preferentially the bonds with bulky and hydrophobic residues such as Leu and Phe at the C-terminal part [19, 20]. When compared with other brevilysins, H2 showed a similar specificity to L4 (Fig. 5). H2 appears to be specific for X-Leu bonds.
Fibrinogen is an oligomeric protein composed of two sets of three subunits (Aα, Bβ, and γ chains). A number of snake venom proteases can digest fibrinogen as a fibrinogenase. This is often related to the toxic action of hemorrhagic venoms [21]. Venom proteinases with fibrinogenase activity are divided into two groups: α-fibrinogenase, a metalloproteinase specific to Aα chain of fibrinogen and β-fibrinogenase, a serine protease specific to Bβ- and Aα-chains of fibrinogen [22]. Brevilysin H2 is an α-fibrinogenase because it specifically attacked the Aα chain.
Brevilysin H1 is also an α-fibrinogenase. Both enzyme are non-hemorrhagic.
Fig. 6. Summary of sequence determination of brevilysin H2. Peptides derived from digestions of PE-H2 with Lys-C (K), Asp-N (D), Arg-C (R), and BrCN (M) are shown below the sequence. Residues not identified are indicated by X.
Fig. 7. Sequence heterogeneity in the C-terminal region of brevilysin H2. Major and minor isozymes are cited by H2a and H2b, respectively.
Hyphens in the sequence of H2b show the identical residues to those of H2a. Names of peptide fragments from H2b are underlined.
The primary structures of many SVMPs have been determined or deduced from the nucleotide sequence [23, 24]. SVMPs are classified into three groups according to their domain structures [25].
The P-I class is composed of a M domain, while the P-II class consists of a M domain and a D domain (a MD domain). The P-III class has an additional C domain. The previous P-IV class containing C-terminal disulfide-linked C-type lectin domains was reclassified as part of class III [4]. The P-III SVMPs usually possess much diverged activities such as hemorrhage, fibrinogenolysis, apoptosis induction, edema formation, extracellular matrix degradation, and modulation of coagulation and complement systems [1]. Many structure-function studies showed that P-III SVMP was more strongly hemorrhagic than P-I and P-II SVMPs [26]. In Fig. 8, the amino acid sequences of 5
brevilysins are compared. Brevilysin L6 is a P-I class SVMP composed solely of a M domain. Brevilysin L4 is a P-II class enzyme having a MD domain. H1, H2, and H6 are typical P-III class SVMPs composed of M, D, and C domains (a MDC domain).
By BLAST search using the sequence data of brevilysin H2a, the primary structures of SVMPs type III isozymes (63 % identity) from Crotalus adamanteus and Agkistrodon piscivorus leucostoma, HR1b from Trimeresurus flavoviridis (65 % identity), and atrolysin-A from Crotalus atrox (64 % identity) showed high similarity to H2a.
Recent crystallographic studies of P-III SVMPs have shed new light on their structure-function properties [27]. They have a common C-shaped MDC domain. The cysteine-rich domain at the distal end points toward the catalytic site of the M domain. The
Fig. 8. Comparison of the amino acid sequence of brevilysin H2 with those of other brevilysins from G. blomhoffi brevicaudus venom. H1, H6, L4, and L6 denotes brevilysins H1 [8], H6 [9], L4 [6], and L6 [7], respectively.
The residues are numbered according to the sequence of H2. Gaps were inserted to maximize similarity.
Asterisks under sequences indicate conserved residues. The zinc-binding region is boxed with a solid line.
Hyper-variable-region (HVR) is marked with a broken line. Cysteine residues are highlighted. Glycosylated asparagine residues are cited in bold.
structures of the peripheral loops in the C domain differ markedly among these proteins, providing extended surface areas, and this region has been designated as the hyper-variable region (HVR) [28].
Each P-III SVMP has a distinct HVR sequence, which results in distinct surface features. Therefore, they might function in specific protein-protein interactions, explaining the diversity of biological activities characteristic of P-III SVMPs. The HVR may constitute an exosite that captures target molecules and contribute to the substrate specificity of the enzyme [28]. The sequence heterogeneity observed in H2 was limited in its C domain ranging from residues 301 to 424. The HVR of H2 spans residues 375-397, and the C-terminal half of HVR differs markedly between brevilysin H2a and H2b. This may influence on a subtle difference in the substrate specificity and thus on their physiological roles of these isozymes.
The HVRs in P-III SVMPs can be involved in the binding to several SVMP inhibitors. HSF and MSF are such proteins isolated as antihemorrhagic factors from the blood of venomous snakes, T. flavoviridis and G. blomhoffi brevicaudus, respectively [15, 29].
They are fetuin-family proteins and inhibit many hemorrhagic SVMPs, but show only weak or no inhibition against P-I SVMPs [15, 30]. Despite of P-III class proteins, some high-molecular mass SVMPs are also insensitive to HSF/MSF. They include brevilysin H1 [8], brevilysin H2 [5, 15], and HV1, an apoptosis- inducing SVMP isolated from T. flavoviridis venom [31, 32]. Interestingly, the inhibition by HSF is non- competitive, indicating that it does not bind to the active site of SVMP [32].
Brevilysin L4 was found as a P-I SVMP at first [33]. However, cDNA cloning of L4 revealed that this protein is a P-II class enzyme and a precursor of brevicaudin 1 [34], a disintegrin previously isolated from the same venom. The P-II SVMPs have been suspected to undergo autolysis to release disintegrins.
Some P-III SVMPs can also autolyse under several conditions, giving rise to a stable fragment composed of a disintegrin-like domain and a cysteine-rich domain [9, 35, 36]. However, H2 was stable to such an autoproteolysis (Fig. 2B). Structural features that associate with the mechanism of autolysis are to be elucidated.
References
[1] J. B. Bjarnason and J. W. Fox, Pharmacol.
Ther. 62, 325-372 (1994).
[2] N. D. Rawlings and A. J. Barrett, in Methods in Enzymology (A. J. Barrett, ed.) Vol. 248, pp.
183-228, Academic Press, New York (1995).
[3] H. Takeya, K. Oda, T. Miyata, T. Omori-Satoh, and S. Iwanaga, J. Biol. Chem. 265, 16068-16073 (1990).
[4] J. W. Fox and S. M. Serrano, FEBS J. 275, 3016-3030 (2008).
[5] M. Deshimaru and S. Terada, Fukuoka Univ.
Sci. Rep. 33, 37-43 (2003).
[6] M. Deshimaru, M. Ichihara, T. Hattori, K.
Koba, and S. Terada, Toxicon 45, 571-580 (2005).
[7] S. Terada, J. Hori, S. Fujimura, and E. Kimoto, J. Biochem. 125, 64-69 (1999).
[8] S. Terada, Y. Tsumura, and A. Kawabata, Fukuoka Univ. Sci. Rep. 41, 195-202 (2011).
[9] S. Fujimura, K. Oshikawa, S. Terada, and E.
Kimoto, J. Biochem. 128, 167-173 (2000).
[10] S. S. Twining, Anal. Biochem. 143, 30-34 (1984).
[11] F. Sanger, Biochem. J. 44, 126-128 (1949).
[12] U. K. Laemmli, Nature 227, 680-685 (1970).
[13] M. Friedman, L. H. Krull, and J. F. Cavins, J.
Biol. Chem. 245, 3868-3871 (1970).
[14] N. Nishino and J. C. Powers, J. Biol. Chem.
255, 3482-3486 (1980).
[15] N. Aoki, K. Tsutsumi, M. Deshimaru, and S.
Terada, Toxicon 51, 251-261 (2008).
[16] S. Swenson and F. S. Markland, Jr., Toxicon 45, 1021-1039 (2005).
[17] M. Satake, Y. Murata, and T. Suzuki, J.
Biochem. 53, 438-447 (1963).
[18] M. T. Assakura, A. P. Reichl, M. C. A. Asperti, and F. R. Mandelbaum, Toxicon 23, 691-706 (1985).
[19] D. J. Civello, J. B. Moran, and C. R. Geren, Biochemistry 22, 755-762 (1983).
[20] T. Nikai, N. Mori, M. Kishida, H. Sugihara, and A. T. Tu, Arch. Biochem. Biophys. 231, 309- 319 (1984).
[21] C. Y. Lee and S. Y. Lee, in Snake Venoms (Lee, C.Y., ed.), Springer-Verlag, New York, pp. 547- 590 (1979).
[22] R.A. Hutton and D.A. Warell, Blood Reviews 7, 176-189 (1993).
[23] J. B. Bjarnason and J. W. Fox, in: Methods in Enzymology (A. J. Barrett, ed.), Vol. 248, pp.
345-368 (1995).
[24] J. W. Fox and S. M. Serrano, J. Proteomics 72, 200-209 (2009).
[25] L. A. Hite, L. G. Jia, J. B. Bjarnason, and J. W.
Fox, Arch. Biochem. Biophys. 308, 182-191 (1994).
[26] A. M. Moura-da-Silva, O. H. Ramos, C. Baldo, S.
Niland, U. Hansen, J. S. Ventura, S. Furlan, D.
Butera, M. S. Della-Casa, I. Tanjoni, P. B. Clissa, I. Fernandes, A. M. Chudzinski-Tavassi, and J.
A. Eble, Biochimie 90, 484-492 (2008).
[27] S. Takeda, H. Takeya, and S. Iwanaga, Biochim.
Biophys. Acta 1824, 164-176 (2011).
[28] S. Takeda, T. Igarashi, H. Mori, and S. Araki, EMBO J. 25, 2388-2396 (2006).
[29] Y. Yamakawa and T. Omori-Satoh, J. Biochem.
112, 583-589 (1992).
[30] M. Deshimaru, C. Tanaka, K. Fujino, N. Aoki, S.
Terada, S. Hattori, and M. Ohno, Toxicon 46, 937-945 (2005).
[31] S. Masuda, H. Hayashi, H. Atoda, T. Morita, and S. Araki, Eur. J. Biochem. 268, 3339-3345 (2001).
[32] N. Shioi, Y. Mizukami, S. Abe, R. Hayashi, and S. Terada, J. Biochem. in press (2012).
[33] S. Fujimura, T. Rikimaru, S. Baba, J. Hori, X.-Q.
Hao, S. Terada, and E. Kimoto, Biochim.
Biophys. Acta 1243, 94-100 (1995).
[34] K. Oshikawa, Y. Yasukouchi, and S. Terada, Fukuoka Univ. Sci. Rep. 30, 201-208 (2000).
[35] Y. Usami, Y. Fujimura, S. Miura, H. Shima, E.
Yoshida, A. Yoshioka, K. Hirano, M. Suzuki, and K. Titani, Biochem. Biophys. Res. Commun.
201, 331-339 (1994).
[36] A. M. Moura-da-Silva, M. S. Della-Casa, A. S.
David, M. T. Assakura, D. Butera, I. Lebrun, J.
D. Shannon, S. M. Serrano, and J. W. Fox, Arch. Biochem. Biophys. 409, 395-401 (2003).