Tyrosine phosphorylation signaling
in Src-mediated suppression of apoptosis
(Src 型チロシンキナーゼによるチロシンリン酸化
シグナリングを介したアポトーシス抑制機構の解明
)
2017
CONTENTS
Introduction ··· 3
Materials and Methods ··· 5
Results ··· 8
Discussion ··· 13
References ··· 15
Figures ··· 21
Tables ··· 25
Main Publication ··· 28
Examiners ··· 29
Acknowledgements ··· 30
Introduction
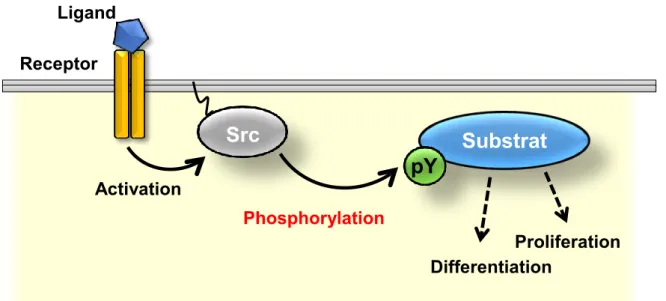
Protein-tyrosine phosphorylation is one of the key posttranslational modifications that regulate various cellular events, such as cell proliferation, cell adhesion, and differentiation (Hubbard and Till, 2000; Hunter, 2009) (Figure 001). Human genome encodes 518 protein kinases, including 428 serine and threonine kinases and 90 tyrosine kinases (Manning et al., 2002). In contrast to receptor-type tyrosine kinases, the functions of non-receptor-type tyrosine kinases depend on their subcellular localizations (Ullrich and Schlessinger, 1990).
Src-family tyrosine kinases, which are non-receptor-type kinases, consist of proto-oncogene products and structurally related proteins, such as c-Src, Lyn, and Fyn (Brown and Cooper, 1996; Thomas and Brugge, 1997) (Figure 002). Multiple members of Src-family kinases are expressed in many cell types and are involved in individual and overlapping signaling pathways (Brown and Cooper, 1996; Thomas and Brugge, 1997). It is generally accepted that Src-family kinases are predominantly located at the cytoplasmic face of the plasma membrane through posttranslational palmitoylation and myristoylation, but in fact, appreciable fractions are found in a variety of intracellular locations, such as lysosomes, late endosomes, the Golgi apparatus, and the nucleus (Kaplan et al., 1992; Ley et al., 1994; Möhn et al., 1995; Bijlmakers et al., 1997; Cans et al., 2000; Yamaguchi et al., 2001; Kasahara et al., 2004; Kasahara et al., 2007a; Kasahara et al., 2007b; Moorhead et al., 2007; Sato et al., 2009; Obata et al., 2010; Honda et al., 2016).
We have shown that Lyn, a member of Src-family kinases, is imported into and rapidly exported from the nucleus (Ikeda et al., 2008), and that nuclear tyrosine phosphorylation plays a role in global changes of chromatin structure and histone modifications (Yamaguchi et al., 2001, Takahashi et al., 2009; Aoyama et al., 2011; Kubota et al., 2015). To explore the roles of protein-tyrosine phosphorylation in cell functions, we recently performed phosphoproteomic analyses (Aoyama et al., 2013; Kubota et al., 2013) and identified novel roles for tyrosine phosphorylation of nuclear proteins, such as the transcription factors JunB (Yamaguchi et al., 2015) and FoxA1 (Yamaguchi et al., 2016), the heterochromatin protein KAP1/TIF1ß/TRIM28 (Kubota et al., 2013), and the chromatin-associated protein AKAP8 (Kubota et al., 2015).
Src-family kinases have been reported to suppress apoptosis by upregulating anti-apoptotic genes (Gillet et al., 1995 Johnson et al., 2000) and downregulating proapoptotic genes (Reginato et al., 2005; Lopez et al., 2012). Although Src-family kinases are well known as an activator of the major downstream pathways involvedin growth and survival signaling, such as the Ras-mitogen-activated-protein-kinase (Ras–MAPK) pathway, the phosphatidylinositol-3-kinase-Akt (PI3K-Akt) pathway, and the STAT3 pathway (Kulik et al., 1997; Bonni et al., 1999; Martin, 2001; Shen et al., 2001), it was reported that caspase-8 and the growth arrest and DNA damage protein 34 (GADD34) are associated with
suppression of apoptosis through tyrosine phosphorylation mediated by Src-family kinases (Grishin et al., 2001; Cursi et al., 2006). Despite the significance of Src-family kinases in suppression of apoptosis, the mechanism of Src-mediated suppression of apoptosis remains to be elucidated.
In this study, I identify Ku70 as a substrate of Src-family kinases involved in apoptosis signaling. Inhibition of the kinase activity of endogenous Src-family kinases enhances the level of apoptosis upon apoptotic stimulation. Phosphorylation of Ku70 at Tyr-530 by Src-family kinases decreases acetylation of Ku70, resulting in suppression of apoptosis. My results demonstrate a novel role of Ku70’s tyrosine phosphorylation in suppression of apoptosis.
Materials and Methods
Plasmids—The sequences of the primers used in this study are shown (Table 1, 2). To construct myc-tagged wild-type Ku70 (myc-Ku70-wt), the BamHI and XhoI sites were created at the both ends of human wild-type Ku70 (Ku70-wt; Open Biosystems) by PCR. Ku70-wt was inserted into the BamHI-XhoI site of the myc-pcDNA3 vector, a myc tag vector (Aoyama et al., 2015). The Tyr to Phe mutant of Ku70 (myc-Ku70-YF) was created by PCR using Ku70-wt as a template. The Tyr to Glu mutant of Ku70 (myc-Ku70-YE) was created by PCR using Ku70-wt as a template. Short hairpin RNA (shRNA)-resistant (shR) Ku70
(Ku70/shR) constructs were made by mutation of the shKu70-CDS target site by PCR using
Ku70 as a template. pcDNA4/TO/puro/myc-Ku70-wt/shR or its mutants were constructed as
follows: the BamHI–XhoI fragment of pcDNA3/myc-Ku70-wt/shR,
pcDNA3/myc-Ku70-YF/shR, or pcDNA3/myc-Ku70-YE/shR was subcloned into the
myc-pcDNA4/TO/puro vector (Kubota et al., 2015). pEBmulti/puro/myc-Ku70-wt or pEBmulti/puro/myc-Ku70-YF was constructed as follows: the XhoI and NotI sites were created at the both ends of pcDNA3/myc-Ku70 by PCR, and the XhoI-NotI fragment of the PCR product was subcloned into the episomal pEBmulti/puro vector (Yuki et al., 2015). Bax was generated by PCR from HeLa cell cDNAs, and the EcoRI-XhoI fragment of the PCR product was subcloned into the EcoRI-XhoI site of the pcDNA3/Flag vector (Kubota et al., 2013), resulting in the construction of Flag-tagged Bax (Flag-Bax). Intact Lyn and Lyn tagged with a nuclear localization signal (NLS-Lyn) was constructed from cDNA encoding human wild-type Lyn (Yamanashi et al., 1987; provided by T. Yamamoto) as described (Sato et al., 2009; Takahashi et al., 2009). Intact c-Src and HA-tagged c-Src (c-Src-HA) were constructed from cDNA encoding human wild-type Src (Bjorge et al., 1995; provided by D. J. Fujita) as described (Sato et al., 2009; Takahashi et al., 2009). HA-tagged, kinase-dead c-Src(K298R) [c-Src(KD)] was constructed as described previously (Fessart et al., 2005; provided by S.A. Laporte). Intact Fyn was constructed from cDNA encoding human wild-type Fyn (Tezuka et al., 1999; provided by T. Yamamoto) as described (Sato et al., 2009; Takahashi et al., 2009). The nucleotides for shRNA (Table 3) were annealed and subcloned into the BglII-XbaI site of the pENTR4/H1 vector (provided by H. Miyoshi) (Nakayama et al., 2009; Obata et al., 2010; Morii et al., 2015).The EBNA1-based episomal pEBmulti/neo/H1 vector, which encodes the H1 promoter and a neomycin-resistant gene, was generated from the pEBmulti/neo vector by replacing the CAG promoter with the H1 promoter as described (Kubota et al., 2015, Hasegawa et al., 2014). pEBmulti/neo/H1-sh(c-Src) was constructed as follows: the SpeI fragment of the PCR product was subcloned into the SpeI site of the pEBmulti/neo vector (Hasegawa et al., 2014).
Cells and cell culture—Cells (Table5) were cultured in Iscove’s modified Dulbecco’s medium (IMDM) containing 5% bovine serum (BS). To establish a stable Ku70-knockdown
cell line (HeLa S3/shKu70), HeLa S3 cells were cotransfected with pENTR4/H1/shKu70 and a plasmid containing the hygromycin resistant gene, and selected in 250 µg/ml hygromycin. To establish stable cell lines for tetracycline-inducible myc-Ku70-wt, myc-Ku70-YF, or myc-Ku70-YE expression, HeLa S3/TR cells (Aoyama et al., 2011) transfected with pcDNA4/TO/puro/myc-Ku70-wt/shR, pcDNA4/TO/puro/myc-Ku70-YF/shR, or
pcDNA4/TO/puro/myc-Ku70-YE/shR were selected in 350 ng/ml puromycin. Expression of
myc-Ku70-wt, myc-Ku70-Y530F, or myc-Ku70-Y530E was induced by the addition of 1 µg/ml doxycycline, a tetracycline derivative. To stably express myc-Ku70-wt or myc-Ku70-YF in HeLa S3/shKu70 cells, cells transfected with pEBmulti/puro/myc-Ku70-wt and pEBmulti/puro/myc-Ku70-Y5F were selected in 1.5 µg/ml puromycin. To generate c-Src-knockdown cells, HeLa S3 cells transfected with pEBmulti/neo/H1 or pEBmulti/neo/H1-sh(c-Src) were selected in 0.5-1 mg/ml G418. To inhibit Src-mediated tyrosine phosphorylation, cells were treated with 20 µM PP2 or 10 µM SU6656.
Western blotting, Immunoprecipitation and Immunofluorescence— Antibodies used in this study are shown (Table 4). Western blotting was performed with enhanced chemiluminescence (Millipore) as described previously (Kasahara et al., 2004; Aoyama et al., 2013; Kubota et al., 2015). Immunoprecipitation was performed as described previously (Mera et al., 1999; Matsuda et al., 2006; Obata et al, 2010; Ikeda et al., 2008; Aoyama et al., 2015). To detect tyrosine phosphorylation of Ku70, cells were suspended in SDS lysis buffer (0.1% SDS, 50 mM Tris-HCl, pH 7.5, 10 mM Na3VO4, 10 mM unbuffered HEPES, 4 µg/ml
aprotinin, 1.6 µg/ml pepstatin A, 4 µg/ml leupeptin, 1 mM EDTA, and 1 mM PMSF). Then, Triton X-100 was added to the cell lysates at a final concentration of 0.5%. After centrifugation, the cell lysates were subjected to immunoprecipitation. To detect tyrosine phosphorylation of endogenous Ku70, cells were treated with pervanadate. Pervanadate (10 mM) was prepared in Dulbecco’s phosphate-buffered saline (D-PBS) for 5 min at 4°C from a 100-mM sodium orthovanadate (Na3VO4) solution and 30% hydrogen peroxide at final
concentrations of 10 mM and 0.1%. Excess hydrogen peroxide was eliminated by adding catalase (Wako Pure Chemical Industries, Osaka) for 5 min at room temperature (Huyer et al., 1997). The pervanadate solution was immediately added to culture medium, and then cells were treated with 0.1 or 0.5 mM pervanadate for 30 min. To detect acetylation levels of Ku70, cells were lysed in PBS containing 1% Triton X-100, 4 µg/ml aprotinin, 1.6 µg/ml pepstatin A, 4 µg/ml leupeptin, 1 mM EDTA, 1 mM PMSF, and 1 µM TSA. Immunofluorescence staining was performed as described previously (Kasahara et al., 2004; Nakayama et al., 2012). In brief, cells fixed in D-PBS containing 4% paraformaldehyde for 20 min were permeabilized in PBS containing 0.1% saponin and 3% bovine serum albumin at room temperature (Tada et al., 1999). To detect Ku70, cells fixed in D-PBS containing 4% paraformaldehyde for 20 min were extracted in D-PBS containing 0.05% Triton X-100 for 5
min, and then blocked in D-PBS containing 0.1% saponin and 3% bovine serum albumin at room temperature. Confocal images were obtained using a Fluoview Fv500 confocal laser scanning microscope with a 20 x 1.00 NA or 40 x 1.00 NA objective (Olympus, Tokyo), as described (Aoyama et al., 2011; Nakayama et al., 2012). Composite figures were prepared using GIMP 2.6.2 and Illustrator 16.0.
Apoptosis assay—For UV irradiation, cells were treated with UV light using a CL-1000 UV crosslinker (UVP LLC) after removal of medium, and cultured in fresh medium for 10-12h. For Adriamycin treatment, cells were incubated with culture medium containing 1 μg/ml Adriamycin (Wako Pure Chemical Industries, Osaka) for the indicated times. Cells were stained with anti-cleaved caspase-3 antibody for detection of caspase-3 activation or with propidium iodide for detection of nuclear condensation/fragmentation. The percentages of apoptotic cells were measured as the ratio of the number of apoptotic cells to the total number of cells from more than three randomly selected fields.
In vitro kinase assay—Cell lysates were prepared in Triton X-100 lysis buffer (30 mM HEPES, pH7.4, 0.5% Triton X-100, 100 mM NaCl, 4 mM EDTA, 10 mM NaF, 50 µg/ml aprotinin, 100 µM leupeptin, 25 µM pepstatin A, and 2 mM PMSF) as described (Kubota et al., 2013). In vitro kinase assays were performed as described (Iwama et al., 1996; Yamaguchi et al., 2001; Kasahara et al., 2004; Matsuda et al., 2006). In brief, c-Src was immunoprecipitated from Triton X-100 lysates of COS-1 cells transfected with c-Src or c-Src(KD). Ku70-wt was immunoprecipitated from Triton X-100 lysates of COS-1 cells transfected with Ku70-wt. After washing, Ku70-wt was eluted with 0.2 M glycine-HCl buffer (pH 2.5) and immediately neutralized with 1 M Tris-HCl (pH 8.8). Equal amounts of c-Src immunoprecipitates bound to the beads were reacted with eluted Ku70-wt in kinase buffer (40 mM HEPES, pH 7.4, 0.1% Triton X-100, 5 mM MnCl2, 5 mM MgCl2, and 1 mM
Results
1. Tyrosine phosphorylation of Ku70
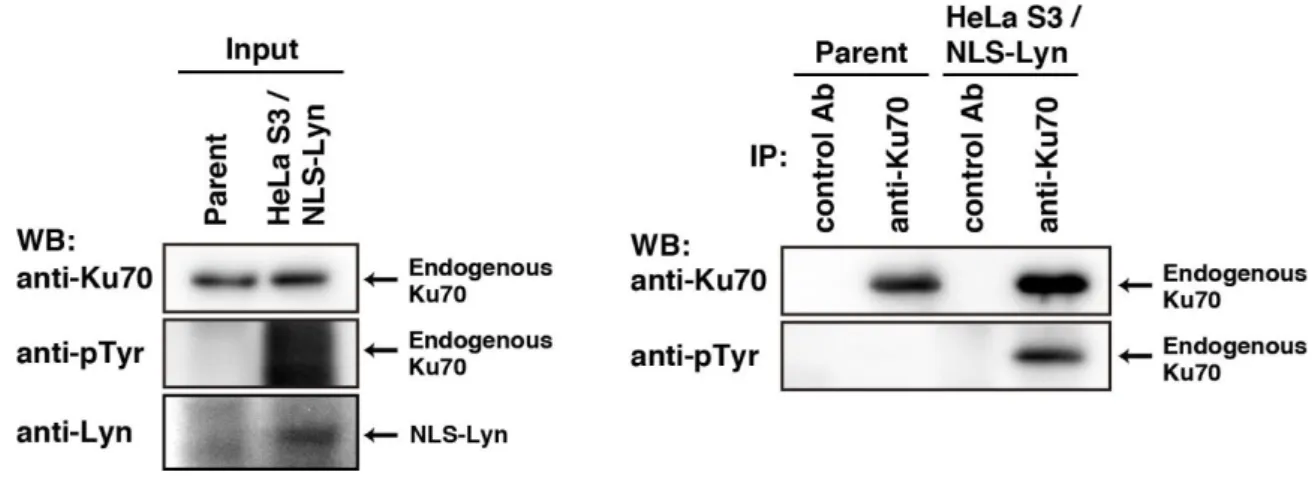
We recently performed phosphoproteomic analyses of tyrosine-phosphorylated proteins in HeLa S3 cells overexpressing the Src-family kinase Lyn tagged with a nuclear localization signal (NLS-Lyn) (Kubota et al., 2013). Purified tyrosine-phosphorylated proteins using anti-phosphotyrosine (pTyr) antibody were digested with trypsin. I found that the tyrosine-phosphorylated peptides detected by LC/MS/MS contained Ku70, a ubiquitously expressed protein, as a candidate substrate of Src-family kinases. To verify tyrosine phosphorylation of endogenous Ku70, I treated HeLa S3 cells stably expressing NLS-Lyn (HeLa S3/NLS-Lyn) were treated with a high dose of the potent tyrosine phosphatase inhibitor Na3VO4, and immunoprecipitated endogenous Ku70 from cell lysates. Endogenous
Ku70 was tyrosine-phosphorylated by NLS-Lyn (Figure 003). Furthermore, cells were cotransfected with wild-type Ku70 (Ku70-wt) plus c-Src or Ku70-wt plus c-Src(KD) and cultured in the presence or absence of the Src inhibitor PP2. I found that expressed Ku70-wt was tyrosine-phosphorylatedby c-Src but not c-Src(KD), and treatment with PP2 inhibited tyrosine phosphorylation of Ku70-wt (Figure 1A in my main publication). To examine whether the other Src members were able to phosphorylate Ku70 at tyrosine residues, I cotransfected cells with Ku70-wt in conjunction with Lyn, c-Src, or Fyn. Among Lyn, c-Src and Fyn, expression of c-Src was found to strongly induce tyrosine phosphorylation of Ku70 (Figure 1B in my main publication). Furthermore, to test whether Ku70 was a direct substrate of c-Src, I transfected COS-1 cells with Ku70-wt, c-Src, or c-Src(KD), immunoprecipitated each protein, and performed in vitro kinase assays. Tyrosine phosphorylation of Ku70-wt by c-Src was detected in a kinase activity-dependent manner (Figure 1C and 1D in my main publication). These results suggest that c-Src directly tyrosine-phosphorylates Ku70.
2. Tyrosine phosphorylation of Ku70 at Tyr530
Our recent phosphoproteomic analysis (Kubota et al., 2013) showed a tyrosine-phosphorylated peptide of Ku70-phospho-Tyr530 (Figure 004). I generated a Ku70 mutant that has a phenylalanine substitution for Tyr530 (Ku70-YF) (Figure 2A in my main publication). COS-1 cells were cotransfected with c-Src plus Ku70-wt or Ku70-YF. The level of tyrosine phosphorylation of Ku70-YF was decreased to approximately 56% of that of Ku70-wt (Figure 2B in my main publication). These results suggest that Tyr-530 is one of the major tyrosine phosphorylation sites of Ku70.
3. Involvement of Src-family kinases and Ku70 in suppression of apoptosis
Previous studies reported that Ku70 plays a role in suppression of apoptosis (Cohen et al., 2004a; Cohen et al., 2004b; Subramanian et al., 2005; Mazumder et al., 2007; Subramanian et al., 2007; Dumitru et al., 2012). To understand the regulation of the ability of Ku70 to
suppress apoptosis, I established stable Ku70-knockdown cell lines (Figure 4A and 4B in my main publication). To examine the effect of Ku70 knockdown on UV-induced apoptosis, I treated parental HeLa S3 cells and HeLa S3/shKu70 cells with UV light and measured the percentage of apoptotic cells by staining for cleaved caspase-3 and/or nuclear condensation/fragmentation (Figure 4C in my main publication, Figure 005). In agreement with previous studies (Cohen et al., 2004a), an increase in UV-induced apoptosis was observed in Ku70 knockdown cells (Figure 4D in my main publication).
To examine the involvement of Src-family kinases and Ku70 in UV-induced apoptosis, I treated parental HeLa S3 cells and HeLa S3/shKu70 cells with UV light in the presence or absence of PP2 or SU6656, another Src inhibitor. It was found that Src inhibition increased the level of apoptosis in parental HeLa S3 cells (Figure 4E in my main publication, open bars), but not in Ku70-knockdown cells (Figure 4E in my main publication, solid bars). To substantiate that the kinase activity of Src-family kinases was critical for suppression of apoptosis, parental HeLa S3 cells and HeLa S3/shKu70 cells transfected with the c-Src or Lyn were treated with UV light. I found that c-Src overexpression significantly suppressed UV-induced apoptosis in HeLa S3 cells (Figure 4F in my main publication, open bars) but not in Ku70-knockdown cells (Figure 4F in my main publication, solid bars). Taken together, these results suggest that Src-family kinases suppress UV-induced apoptosis through Ku70.
4. Inability of Src-family kinases to be activated upon UV irradiation
To examine whether Src-family kinases were activated by UV irradiation, autophosphorylation levels of endogenous Src-family kinases were detected by Western blotting with anti-Src[pY416] antibody. Cells were starved for 20 h and then treated with EGF. EGF stimulation for 5 min strongly induced autophosphorylation of endogenous Src-family kinases and increased the levels of tyrosine phosphorylation of cellular proteins, including ERK kinases (Figure 5A in my main publication). In contrast, UV irradiation did not increase those of tyrosine phosphorylation (Figure 5B in my main publication). Furthermore, those of tyrosine phosphorylation was decreased by PP2 treatment (Figure 5C in my main publication). These results suggest that the basal level of endogenous Src activity is maintained under normal culture conditions and is not increased by UV irradiation.
5. Contribution of Src members to suppression of apoptosis
To verify tyrosine phosphorylation of endogenous Ku70, I treated cells with or without UV light in the presence of pervanadate, an extremely potent protein-tyrosine phosphatase inhibitor. I immunoprecipitated endogenous Ku70 with anti-Ku70 antibody and succeeded in detecting tyrosine phosphorylation of endogenous Ku70 irrespective of UV irradiation (Figure 6A in my main publication). Importantly, tyrosine phosphorylation of endogenous Ku70 was inhibited by treatment with PP2 (Figure 6A in my main publication). Furthermore,
I examined the effect of knockdown of Src members on the level of tyrosine phosphorylation of Ku70 and apoptosis in HeLa S3 cells. c-Src knockdown decreased the level of tyrosine phosphorylation of Ku70 (Figure 6B in my main publication), and significantly increased the levels of UV- and Adriamycin -induced apoptosis (Figure 6C in my main publication). We recently established a HeLa S3 cell line stably expressing shRNA against Lyn and/or Fyn (Morinaga et al., 2014). I immunoprecipitated endogenous Ku70 from Fyn-knockdown cells and Lyn-knockdown cells and found that the levels of tyrosine phosphorylation of endogenous Ku70 were slightly decreased by Fyn knockdown but rather increased by Lyn knockdown (Figure 6D in my main publication). In contrast to single knockdown of Fyn or Lyn, Fyn/Lyn double-knockdown markedly decreased the levels of tyrosine phosphorylation of endogenous Ku70 (Figure 6D in my main publication). Furthermore, the levels of UV- and Adriamycin-induced apoptosis were significantly increased by Fyn/Lyn double-knockdown (Figure 6E in my main publication). Taken together, these results suggest that c-Src, Fyn and Lyn, the major Src members in HeLa S3 cells, play a redundant role in tyrosine phosphorylation of Ku70 and suppression of apoptosis.
6. Role for tyrosine phosphorylation of Ku70 at Tyr-530 in suppression of apoptosis
Replacement of a tyrosine residue with a glutamic acid residue would exert a phosphomimetic effect owing to its negative charge. To examine the role of tyrosine phosphorylation of Ku70 at Tyr-530 in suppression of apoptosis, I established HeLa S3 cell lines stably expressing tetracycline-inducible Ku70-wt, its nonphosphorylatable YF mutant, or its phosphomimetic YE mutant. These cells were treated with doxycycline for induction of Ku70-wt, Ku70-YF, or Ku70-YE, and then irradiated with UV (Figure 7A in my main publication) (Figure 006). Expression of Ku70-wt suppressed UV-induced apoptosis (Figure 7B in my main publication), in agreement with previous studies (Cohen et al., 2004a). Notably, expression of Ku70-YE suppressed UV-induced apoptosis to a similar extent to that of Ku70-wt, but expression of Ku70-YF could not suppress UV-induced apoptosis so much (Figure 7B in my main publication). In the presence of PP2, there was no difference in the levels of apoptosis between Ku70-wt-expressing cells and Ku70-YF-expressing cells (Figure 7C in my main publication). To confirm these results, I transiently rescued HeLa S3/shKu70 cells with Ku70-wt and its nonphosphorylatable YF mutant, or its phosphomimetic YE mutant (Figure 7D in my main publication), and found that similar results were observed in HeLa S3/shKu70 cells upon treatment with UV or Adriamycin (Figure 7E, 7F and 7G in my main publication). These results suggest that tyrosine phosphorylation of Ku70 at Tyr-530 plays a role in suppression of apoptosis.
Next, apoptosis can be initiated by activation of the proapoptotic factor Bax (Scorrano and Korsmeyer, 2003; Tait and Green, 2003), and Ku70 suppresses Bax-mediated apoptosis by sequestering Bax away from mitochondria through the Ku70–Bax interaction (Cohen et al.,
2004a; Cohen et al., 2004b; Subramanian et al., 2005; Mazumder et al., 2007; Subramanian et al., 2007; Dumitru et al., 2012). Cotransfection of Bax with control vector, c-Src, or v-Src, a highly activated variant of c-Src, showed that the levels of Bax-mediated apoptosis were decreased by expression of c-Src and v-Src (Figure 7H in my main publication). To examine the effect of tyrosine phosphorylation of Ku70 at Tyr-530 on Bax-induced apoptosis, HeLa S3 cells inducibly expressing v-Src were transfected with Bax plus Ku70-wt or Bax plus Ku70-YF (Figure 7I in my main publication). Expression of Ku70-wt suppressed Bax-induced apoptosis (Figure 7I in my main publication), in agreement with previous studies (Cohen et al., 2004a). In contrast, expression of Ku70-YF showed a significant reduction in suppression of Bax-induced apoptosis compared with that of Ku70-wt (Figure 7I in my main publication). Taken together, these results suggest that Src-mediated phosphorylation of Ku70 at Tyr-530 is important for suppression of Bax-mediated apoptosis.
7. Role of Ku70 phosphorylation at Tyr-530 in acetylation of Ku70
Previous studies reported that the multiple lysine residues in Ku70 are acetylated, and that acetylation of Ku70 in the C-terminal flexible linker region adjacent to the Bax-interaction domain chiefly contributes to promote apoptosis by possible inactivation of the Ku70’s Bax-interaction domain (Cohen et al., 2004a; Cohen et al., 2004b; Subramanian et al., 2005; Subramanian et al., 2007). My findings demonstrate that phosphorylation of Ku70 at Tyr-530 is important for suppression of apoptosis (Figure 7 in my main publication) and that the phosphorylation site of Ku70 is located close to the possible acetylation sites in the C-terminal flexible linker region (Figure 2 in my main publication), leading to the intriguing hypothesis that tyrosine phosphorylation of Ku70 interferes with acetylation of Ku70. To examine the levels of Ku70 acetylation upon UV irradiation, cells expressing Ku70-wt were treated with UV light in the presence or absence of PP2 and performed immunoprecipitation assays using anti-myc antibody. I found that Src inhibition enhanced the level of acetylation of Ku70-wt upon UV irradiation (Figure 8A and 8B in my main publication). Conversely, I immunoprecipitated Ku70-wt with anti-acetylated lysine antibody from cells expressing Ku70-wt (Figure 5C in my main publication) and confirmed that acetylation of Ku70-wt is increased by Src inhibition upon UV irradiation (Figure 5D and 5E in my main publication). These results suggest that Src-mediated tyrosine phosphorylation of Ku70 inhibits acetylation of Ku70. Next, I assessed the levels of acetylation of Ku70-wt and Ku70-YE upon UV irradiation. I immunoprecipitated Ku70-wt or Ku70-YE with anti-acetylated lysine antibody from HeLa S3 cells inducibly expressing Ku70-wt or Ku70-YE, and found that expression of Ku70-YE significantly decreased the level of its acetylation compared with that of Ku70-wt (Figure 007). Furthermore, by means of knockdown-rescue, I examined the levels of acetylation of Ku70-wt and Ku70-YF upon UV irradiation (Figure 8F in my main publication). Notably, expression of Ku70-YF significantly enhanced the level of its
acetylation compared with that of Ku70-wt (Figure 8G in my main publication). These results suggest that the level of phosphorylation at Tyr-530 on Ku70 is inversely correlated with the level of Ku70 acetylation.
Discussion
Ku70 is known to suppress apoptosis by negatively regulating Bax activation (Cohen et al., 2004a; Cohen et al., 2004b; Subramanian et al., 2005; Mazumder et al., 2007; Subramanian et al., 2007; Dumitru et al., 2012). Upon apoptotic stimulation, acetylation of Ku70 in the C-terminal flexible linker region results in activating Bax, thereby promoting apoptosis (Cohen et al., 2004a; Cohen et al., 2004b; Subramanian et al., 2005; Subramanian et al., 2007).
In the present study, I provide evidence that Src-family kinases act as an effector for Ku70-dependent suppression of apoptosis. First, Src-family kinases phosphorylate Ku70 at Tyr-530. Second, Ku70 knockdown impairs c-Src-mediated suppression of apoptosis, Third, phosphorylation of Ku70 at Tyr-530 suppresses UV-, Adriamycin- or Bax-induced apoptosis. Fourth, inhibition of the activity of endogenous Src-family kinases increases the level of Ku70 acetylation. Fifth, Ku70 mutation at Tyr-530 to phenylalanine enhances acetylation of Ku70.
Thus, I present a model for reduced apoptotic susceptibility by Src-family kinases (Figure 8 in my main publication). Src-family kinases phosphorylate Ku70 at Tyr-530, which is located close to the possible acetylation sites in the C-terminal flexible linker region (Cohen et al., 2004a). Phosphorylation of Ku70 at Tyr-530 decreases acetylation of Ku70, resulting in a reduction in susceptibility to apoptotic stimuli. Inhibition of the activity of Src-family kinases decreases phosphorylation of Ku70 at Tyr-530 and concomitantly enhances the level of acetylation of Ku70, thereby promoting Bax-mediated apoptosis. My results suggest the intriguing possibility that endogenous Src activity raises the cellular threshold to apoptotic stimuli through tyrosine phosphorylation of Ku70 in every cell. Taken together with ubiquitous expression of Src-family kinases and Ku70, I hypothesize that Src-family kinase is a safety device for Ku70-mediated apoptosis to protect our bodies from hyperactive apoptotic cell death.
I show that the levels of Ku70 acetylation is enhanced by Src inhibition (Figure 8A, 8B, 8D and 8E in my main publication) and by the mutation of Ku70 at Tyr-530 to phenylalanine (Figure 8F and 8G in my main publication). Previous studies reported that Ku70 is acetylated at multiple sites, including the sites located in the C-terminal flexible linker region and the DNA binding domain (Cohen et al., 2004a, Chen et al., 2007), and that acetylation of Ku70 in the C-terminal flexible linker region negatively regulates Ku70-mediated suppression of apoptosis (Cohen et al., 2004a; Cohen et al., 2004b; Subramanian et al., 2005; Subramanian et al., 2007). Ku70 is acetylated by the acetyltransferases CBP and PCAF (Cohen et al., 2004a, Subramanian et al., 2007) and deacetylated by the deacetylases SIRT1, SIRT3, and HDAC6 (Cohen et al., 2004b; Sundaresan et al., 2008; Subramanian et al., 2011). It should be emphasized that Src-mediated phosphorylation of Ku70 at Tyr-530 decreases the levels of
Ku70 acetylation, leading to suppression of apoptosis (Figures 7 and 8 in my main publication). There is no report about the interaction between phosphorylation and acetylation of Ku70 in apoptosis thus far. The crosstalk between phosphorylation and acetylation has been documented among histone and non-histone proteins (Latham and Dent, 2007; Yang and Seto, 2008; Choudhary et al., 2014). Considering that the phosphorylation site of Ku70 at Tyr-530 is located close to the C-terminal flexible linker region (Figures 2 in my main publication), I hypothesize that phosphorylation at Tyr-530 may promote the association of Ku70 with deacetylases or may weaken the association of Ku70 with acetyltransferases. Further experiments are needed to fully understand the relationship between tyrosine phosphorylation and acetylation on Ku70.
Src-family kinases are overexpressed and activated in various human cancers and play important roles in cancer development, progression, and resistance to anticancer therapy (Blume-Jensen and Hunter, 2001; Yeatman, 2004; Kim et al., 2009; Zhang and Yu, 2012). My results suggest that, upon UV treatment, high expression of c-Src markedly reduces susceptibility to apoptosis (Figure 4F in my main publication). I further show that inhibition of Tyr-530 phosphorylation enhances susceptibility to apoptosis (Figure 7 in my main publication), leading to the promising possibility that development of inhibitors for tyrosine phosphorylation of Ku70 might potentiate the sensitivity of cancer cells to anticancer therapy. In conclusion, I identify Ku70 as a substrate of Src-family kinases. This is a first report of the significance of Src-mediated phosphorylation of Ku70 at Tyr-530 in suppression of apoptosis. Further studies will be needed to understand the relationship between tyrosine phosphorylation of Ku70 and cancer cell survival or therapeutic resistance.
References
1. Aoyama, K., Fukumoto, Y., Ishibashi, K., Kubota, S., Morinaga, T., Horiike, Y., Yuki, R., Takahashi, A., Nakayama, Y., and Yamaguchi, N. (2011) Nuclear c-Abl-mediated tyrosine phosphorylation induces chromatin structural changes through histone modifications that include H4K16 hypoacetylation. Exp. Cell Res. 317, 2874-2903
2. Aoyama, K., Yuki, R., Horiike, Y., Kubota, S., Yamaguchi, N., Morii, M., Ishibashi, K., Nakayama, Y., Kuga, T., Hashimoto, Y., Tomonaga, T., and Yamaguchi, N. (2013) Formation of long and winding nuclear F-actin bundles by nuclear c-Abl tyrosine kinase. Exp. Cell Res. 319, 3251–3268
3. Aoyama, K., Yamaguchi, N., Yuki, R., Morii, M., Kubota, S., Hirata, K., Abe, K., Honda, T., Kuga, T., Hashimoto, Y., Tomonaga, T., and Yamaguchi, N. (2015) c-Abl induces stabilization of histone deacetylase 1 (HDAC1) in a kinase activity-dependent manner. Cell Biol. Int. 39, 446–456
4. Bijlmakers, M. J. E., Isobe-Nakamura, M., Ruddock, L. J., and Marsh, M. (1997) Intrinsic signals in the unique domain target p56lck to the plasma membrane independently of CD4. J. Cell Biol. 137, 1029–1040
5. Bjorge, J. D., Bellagamba, C., Cheng, H. C., Tanaka, A., Wang, J. H., and Fujita, D. J. (1995) Characterization of two activated mutants of human pp60c-src that escape c-Src kinase regulation by distinct mechanisms. J. Biol. Chem. 270, 24222–24228
6. Blume-Jensen, P., and Hunter, T. (2001) Oncogenic kinase signalling. Nature 411, 355– 365
7. Bonni, A., Brunet, A., West, A. E., Datta, S. R., Takasu, M. A., and Greenberg, M. E. (1999) Cell survival promoted by the Ras-MAPK signaling pathway by transcription-dependent and -independent mechanisms. Science 286, 1358–1362
8. Brown, M. T., and Cooper, J. A. (1996) Regulation, substrates and functions of src. Biochim. Biophys. Acta 1287, 121–149
9. Cans, C., Mangano, R., Barilá, D., Neubauer, G., and Superti-Furga, G. (2000) Nuclear tyrosine phosphorylation: the beginning of a map. Biochem. Pharmacol. 60, 1203–1215 10. Choudhary, C., Weinert, B. T., Nishida, Y., Verdin, E., and Mann, M. (2014) The growing
landscape of lysine acetylation links metabolism and cell signalling. Nat. Rev. Mol. Cell Biol. 15, 536–550
11. Cohen, H. Y., Lavu, S., Bitterman, K. J., Hekking, B., Imahiyerobo, T. A., Miller, C., Frye, R., Ploegh, H., Kessler, B. M., and Sinclair, D. A. (2004a) Acetylation of the C terminus of Ku70 by CBP and PCAF controls Bax-mediated apoptosis. Mol. Cell 13, 627–638 12. Cohen, H. Y., Miller, C., Bitterman, K. J., Wall, N. R., Hekking, B., Kessler, B., Howitz,
K. T., Gorospe, M., de Cabo, R., and Sinclair, D. A. (2004b) Calorie restriction promotes mammalian cell survival by inducing the SIRT1 deacetylase. Science 305, 390–392
13. Cursi, S., Rufini, A., Stagni, V., Condò, I., Matafora, V., Bachi, A., Bonifazi, A. P., Coppola, L., Superti-Furga, G., Testi, R., and Barilà, D. (2006) Src kinase phosphorylates Caspase-8 on Tyr380: a novel mechanism of apoptosis suppression. EMBO J. 25, 1895– 1905
14. Dumitru, R., Gama, V., Fagan, B. M., Bower, J. J., Swahari, V., Pevny, L. H., and Deshmukh, M. (2012) Human embryonic stem cells have constitutively active Bax at the Golgi and are primed to undergo rapid apoptosis. Mol. Cell 46, 573–583
15. Fessart, D., Simaan, M., and Laporte, S. A. (2005) c-Src regulates clathrin adapter protein 2 interaction with beta-arrestin and the angiotensin II type I receptor during clathrin-mediated internalization. Mol. Endocrinol. 19, 491–503
16. Gillet, G., Guerin, M., Trembleau, A., and Brun, G. (1995) A Bcl-2-related gene is activated in avian cells transformed by the Rous sarcoma virus. EMBO J. 14, 1372–1381 17. Grishin, A. V., Azhipa, O., Semenov, I., and Corey, S. J. (2001) Interaction between
growth arrest-DNA damage protein 34 and Src kinase Lyn negatively regulates genotoxic apoptosis. Proc. Natl. Acad. Sci. U.S.A. 98, 10172–10177
18. Hasegawa, H., Ishibashi, K., Kubota, S., Yamaguchi, C., Yuki, R., Nakajo, H., Eckner, R., Yamaguchi, N., Yokoyama, K. K., and Yamaguchi, N. (2014) Cdk1-mediated phosphorylation of human ATF7 at Thr-51 and Thr-53 promotes cell-cycle progression into M phase. PLoS One 9, e116048
19. Honda, T., Soeda, S., Tsuda, K., Yamaguchi, C., Aoyama, K., Morinaga, T., Yuki, R., Nakayama, Y., Yamaguchi, N., and Yamaguchi, N. Protective role for lipid modifications of Src-family kinases against chromosome missegregation. Sci. Rep. 9, 38751
20. Hubbard, S. R., and Till, J. H. (2000) Protein tyrosine kinase structure and function. Annu. Rev. Biochem. 69, 373–398
21. Hunter, T. (2009) Tyrosine phosphorylation: thirty years and counting. Curr. Opin. Cell Biol. 21, 140–146
22. Huyer, G., Liu, S., Kelly, J., Moffat, J., Payette, P., Kennedy, B., Tsaprailis, G., Gresser, M. J., and Ramachandran, C. (1997) Mechanism of inhibition of protein-tyrosine phosphatases by vanadate and pervanadate. J Biol. Chem. 272, 843–851
23. Ikeda, K., Nakayama, Y., Togashi, Y., Obata, Y., Kuga, T., Kasahara, K., Fukumoto, Y., and Yamaguchi, N. (2008) Nuclear localization of Lyn tyrosine kinase mediated by inhibition of its kinase activity. Exp. Cell Res. 314, 3392–3404
24. Iwama, A., Yamaguchi, N., and Suda, T. (1996) STK/RON receptor tyrosine kinase mediates both apoptotic and growth signals via the multifunctional docking sites conserved among the HGF receptor family. EMBO J. 15, 5866–5875
25. Johnson, D., Agochiya, M., Samejima, K., Earnshaw, W., Frame, M., and Wyke, J. (2000) Regulation of both apoptosis and cell survival by the v-Src oncoprotein. Cell Death Differ. 7, 685–696
26. Kaplan, K. B., Swedlow, J. R., Varmus, H. E., and Morgan, D. O. (1992) Association of p60c-src with endosomal membranes in mammalian fibroblasts. J. Cell Biol. 118, 321-333
27. Kasahara, K., Nakayama, Y., Ikeda, K., Fukushima, Y., Matsuda, D., Horimoto, S., and Yamaguchi, N. (2004) Trafficking of Lyn through the Golgi caveolin involves the charged residues on αE and αI helices in the kinase domain. J. Cell Biol. 165, 641–652
28. Kasahara, K., Nakayama, Y., Kihara, A., Matsuda, D., Ikeda, K., Kuga, T., Fukumoto, Y., Igarashi, Y., and Yamaguchi, N. (2007a) Rapid trafficking of c-Src, a non-palmitoylated Src-family kinase, between the plasma membrane and late endosomes/lysosomes. Exp. Cell Res. 313, 2651–2666
29. Kasahara, K., Nakayama, Y., Sato, I., Ikeda, K., Hoshino, M., Endo, T., and Yamaguchi, N. (2007b) Role of Src-family kinases in formation and trafficking of macropinosomes. J. Cell. Physiol. 211, 220–232
30. Kim, L. C., Song, L., and Haura, E. B. (2009) Src kinases as therapeutic targets for cancer. Nat. Rev. Clin. Oncol. 6, 587–595
31. Kubota, S., Fukumoto, Y., Aoyama, K., Ishibashi, K., Yuki, R., Morinaga, T., Honda, T., Yamaguchi, N., Kuga, T., Tomonaga, T., and Yamaguchi, N. (2013) Phosphorylation of KRAB-associated protein 1 (KAP1) at Tyr-449, Tyr-458, and Tyr-517 by nuclear tyrosine kinases inhibits the association of KAP1 and heterochromatin protein 1α (HP1α) with heterochromatin. J. Biol. Chem. 288, 17871–17883
32. Kubota, S., Morii, M., Yuki, R., Yamaguchi, N., Yamaguchi, H., Aoyama, K., Kuga, T., Tomonaga, T., and Yamaguchi, N. (2015) Role for tyrosine phosphorylation of A-kinase anchoring protein 8 (AKAP8) in its dissociation from chromatin and the nuclear matrix. J. Biol. Chem. 290, 10891–10904
33. Kulik, G., Klippel, A., and Weber, M. J. (1997) Antiapoptotic signalling by the insulin-like growth factor I receptor, phosphatidylinositol 3-kinase, and Akt. Mol. Cell. Biol. 17, 1595–1606
34. Latham, J. A., and Dent, S. Y. (2007) Cross-regulation of histone modifications. Nat. Struct. Mol. Biol. 14, 1017–1024
35. Ley, S. C., Marsh, M., Bebbington, C. R., Proudfoot, K., and Jordan, P. (1994) Distinct intracellular localization of Lck and Fyn protein tyrosine kinases in human T lymphocytes. J. Cell Biol. 125, 639–649
36. Lopez, J., Hesling, C., Prudent, J., Popgeorgiev, N., Gadet, R., Mikaelian, I., Rimokh, R., Gillet, G., and Gonzalo, P. (2012) Src tyrosine kinase inhibits apoptosis through the Erk1/2-dependent degradation of the death accelerator Bik. Cell Death Differ. 19,1459– 1469
37. Manning, G., Whyte, D. B., Martinez, R., Hunter, T., and Sudarsanam, S. (2002) The protein kinase complement of the human genome. Science 298, 1912–1934
38. Martin, G. S. (2001) The hunting of the Src. Nat. Rev. Mol. Cell Biol. 2, 467–475
39. Matsuda, D., Nakayama, Y., Horimoto, S., Kuga, T., Ikeda, K., Kasahara, K., and Yamaguchi, N. (2006) Involvement of Golgi-associated Lyn tyrosine kinase in the translocation of annexin II to the endoplasmic reticulum under oxidative stress. Exp. Cell Res. 312, 1205–1217
40. Mazumder, S., Plesca, D., Kinter, M., and Almasan, A. (2007) Interaction of a cyclin E fragment with Ku70 regulates Bax-mediated apoptosis. Mol. Cell. Biol. 27, 3511–3520 41. Mera, A., Suga, M., Ando, M., Suda, T., and Yamaguchi, N. (1999) Induction of cell
shape changes through activation of the interleukin-3 common beta chain receptor by the RON receptor-type tyrosine kinase. J. Biol. Chem. 274, 15766–15774
42. Möhn, H., Le Cabec, V., Fischer, S., and Maridonneau-Parini, I. (1995) The src-family protein-tyrosine kinase p59hck is located on the secretory granules in human neutrophils and translocates towards the phagosome during cell activation. Biochem. J. 309, 657–665 43. Moorhead, G. B., Trinkle-Mulcahy, L., and Ulke-Lemée, A. (2007) Emerging roles of
nuclear protein phosphatases. Nat. Rev. Mol. Cell Biol. 8, 234–244
44. Morii, M., Fukumoto, Y., Kubota, S., Yamaguchi, N., Nakayama, Y., and Yamaguchi, N. (2015) Imatinib inhibits inactivation of the ATM/ATR signaling pathway and recovery from adriamycin/doxorubicin-induced DNA damage checkpoint arrest. Cell Biol. Int. 39, 923–932
45. Morinaga, T., Abe, K., Nakayama, Y., Yamaguchi, N., and Yamaguchi, N. (2014) Activation of Lyn tyrosine kinase through decreased membrane cholesterol levels during a change in its membrane distribution upon cell detachment. J. Biol. Chem. 289, 26327– 26343
46. Nakayama, Y., Igarashi, A., Kikuchi, I., Obata, Y., Fukumoto, Y., and Yamaguchi, N. (2009) Bleomycin-induced over-replication involves sustained inhibition of mitotic entry through the ATM/ATR pathway. Exp. Cell Res. 315, 2515–2528
47. Nakayama, Y., Matsui, Y., Takeda, Y., Okamoto, M., Abe, K., Fukumoto, Y., and Yamaguchi, N. (2012) c-Src but not Fyn promotes proper spindle orientation in early prometaphase. J. Biol. Chem. 287, 24905–24915
48. Obata, Y., Fukumoto, Y., Nakayama, Y., Kuga, T., Dohmae, N., and Yamaguchi, N. (2010) The Lyn kinase C-lobe mediates Golgi export of Lyn through conformation-dependent ACSL3 association. J. Cell Sci. 123, 2649–2662 20.
49. Reginato, M. J., Mills, K. R., Becker, E. B. E., Lynch, D. K., Bonni, A., Muthuswamy, S. K., and Brugge, J. S. (2005) Bim regulation of lumen formation in cultured mammary epithelial acini is targeted by oncogenes. Mol. Cell. Biol. 25, 4591–4601
50. Sasaki, H., Toyomura, K., Matsuzaki, W., Okamoto, A., Yamaguchi, N., Nakamura, H., and Murayama, T. (2014) Regulation of alkaline ceramidase activity by the c-Src-mediated pathway. Arch. Biochem. Biophys. 550–551, 12–19
51. Sato, I., Obata, Y., Kasahara, K., Nakayama, Y., Fukumoto, Y., Yamasaki, T., Yokoyama, K. K., Saito, T., and Yamaguchi, N. (2009) Differential trafficking of Src, Lyn, Yes and Fyn is specified by the state of palmitoylation in the SH4 domain. J. Cell Sci. 122, 965– 975
52. Scorrano, L., and Korsmeyer, S. J. (2003) Mechanisms of cytochrome c release by proapoptotic BCL-2 family members. Biochem. Biophys. Res. Commun. 304, 437–444 53. Shen, Y., Devgan, G., Darnell, J. E., Jr., and Bromberg, J. F. (2001) Constitutively
activated Stat3 protects fibroblasts from serum withdrawal and UV-induced apoptosis and antagonizes the proapoptotic effects of activated Stat1. Proc. Natl. Acad. Sci. U.S.A. 98, 1543–1548
54. Soeda, S., Nakayama, Y., Honda, T., Aoki, A., Tamura, N., Abe, K., Fukumoto, Y., and Yamaguchi, N. (2013) v-Src causes delocalization of Mklp1, Aurora B, and INCENP from the spindle midzone during cytokinesis failure. Exp. Cell Res. 319, 1382–1397 55. Subramanian, C., Opipari, A. W., Jr., Bian, X., Castle, V. P., and Kwok, R. P. (2005) Ku70
acetylation mediates neuroblastoma cell death induced by histone deacetylase inhibitors. Proc. Natl. Acad. Sci. U.S.A. 102, 4842–4847
56. Subramanian, C., Jarzembowski, J. A., Opipari, A. W., Jr., Castle, V. P., and Kwok, R. P. (2007) CREB-binding protein is a mediator of neuroblastoma cell death induced by the histone deacetylase inhibitor trichostatin A. Neoplasia 9, 495–503
57. Subramanian, C., Jarzembowski, J. A., Opipari, A. W., Jr., Castle, V. P., and Kwok, R. P. (2011) HDAC6 deacetylates Ku70 and regulates Ku70-Bax binding in neuroblastoma. Neoplasia 13, 726–734
58. Sundaresan, N. R., Samant, S. A., Pillai, V. B., Rajamohan, S. B., and Gupta, M. P. (2008) SIRT3 is a stress-responsive deacetylase in cardiomyocytes that protects cells from stress-mediated cell death by deacetylation of Ku70. Mol. Cell. Biol. 28, 6384–6401 59. Tada, J., Omine, M., Suda, T., and Yamaguchi, N. (1999) A common signaling pathway
via Syk and Lyn tyrosine kinases generated from capping of the sialomucins CD34 and CD43 in immature hematopoietic cells. Blood 93, 3723–3735
60. Tait, S. W., and Green, D. R. (2010) Mitochondria and cell death: outer membrane permeabilization and beyond. Nat. Rev. Mol. Cell Biol. 11, 621–632
61. Takahashi, A., Obata, Y., Fukumoto, Y., Nakayama, Y., Kasahara, K., Kuga, T., Higashiyama, Y., Saito, T., Yokoyama, K. K., and Yamaguchi, N. (2009) Nuclear localization of Src-family tyrosine kinases is required for growth factor-induced euchromatinization. Exp. Cell Res. 315, 1117–1141
62. Tamura, T., Kunimatsu, T., Yee, S. T., Igarashi, O., Utsuyama, M., Tanaka, S., Miyazaki, S., Hirokawa, K., and Nariuchi, H. (2000) Molecular mechanism of the impairment in activation signal transduction in CD4(+) T cells from old mice. Int. Immunol. 12, 1205– 1215
63. Tezuka, T., Umemori, H., Akiyama, T., Nakanishi, S., and Yamamoto, T. (1999) PSD-95 promotes Fyn-mediated tyrosine phosphorylation of the N-methyl-D-aspartate receptor subunit NR2A. Proc. Natl. Acad. Sci. U.S.A. 96, 435–440
64. Thomas, S. M., and Brugge, J. S. (1997) Cellular functions regulated by Src family kinases. Annu. Rev. Cell Dev. Biol. 13, 513–609
65. Trachootham, D., Alexandre, J., and Huang, P. (2009) Targeting cancer cells by ROS-mediated mechanisms: a radical therapeutic approach? Nat. Rev. Drug Discov. 8, 579–591
66. Ullrich, A., and Schlessinger, J. (1990) Signal transduction by receptors with tyrosine kinase activity. Cell 61, 203–212
67. Yamaguchi, N., Nakayama, Y., Urakami, T., Suzuki, S., Nakamura, T., Suda, T., and Oku, N. (2001) Overexpression of the Csk homologous kinase (Chk tyrosine kinase) induces multinucleation: a possible role for chromosome-associated Chk in chromosome dynamics. J. Cell Sci. 114, 1631–1641
68. Yamaguchi, N., Yuki, R., Kubota, S., Aoyama, K., Kuga, T., Hashimoto, Y., Tomonaga, T., and Yamaguchi, N. (2015) c-Abl-mediated tyrosine phosphorylation of JunB is required for Adriamycin-induced expression of p21. Biochem. J. 471, 67–77
69. Yamaguchi, N., Shibazaki, M., Yamada, C., Anzai, E., Morii, M., Nakayama, Y., Kuga, T., Hashimoto, Y., Tomonaga, T., and Yamaguchi, N. (Nov. 24, 2016) Tyrosine phosphorylation of the pioneer transcription factor FoxA1 promotes activation of estrogen signaling. J. Cell. Biochem. 10.1002/jcb.25804.
70. Yamanashi, Y., Fukushige, S., Semba, K., Sukegawa, J., Miyajima, N., Matsubara, K., Yamamoto, T., and Toyoshima, K. (1987) The yes-related cellular gene lyn encodes a possible tyrosine kinase similar to p56lck. Mol. Cell. Biol. 7, 237–243
71. Yang, X. J., and Seto, E. (2008) Lysine acetylation: codified crosstalk with other posttranslational modifications. Mol. Cell 31, 449–461
72. Yeatman, T. J. (2004) A renaissance for SRC. Nat. Rev. Cancer 4, 470–480
73. Yuki, R., Aoyama, K., Kubota, S., Yamaguchi, N., Kubota, S., Hasegawa, H., Morii, M., Huang, X., Liu, K., Williams, R., Fukuda, M. N., and Yamaguchi, N. (2015) Overexpression of zinc-finger protein 777 (ZNF777) inhibits proliferation at low cell density through down-regulation of FAM129A. J. Cell. Biochem. 116, 954–968
74. Zhang, S., and Yu, D. (2012) Targeting Src family kinases in anti-cancer therapies: turning promise into triumph. Trends Pharmacol. Sci. 33, 122–128
Figures
Figure 001. Src-mediated tyrosine phosphorylation.
pY, phosphorylated tyrosine residues.
Figure 002. A schematic representation of Src.
SH, Src homology domain; Kinase, the tyrosine kinase domain.
Ligand Activation Receptor Differentiation Proliferation
Src
Substrat
e
P
PhosphorylationpY
SH4
SH3
SH2
Src
Kinase
Figure 003. Tyrosine phosphorulation of endogenous Ku70 in NLS-Lyn expressing cells.
Parental HeLa S3 cells or HeLa S3/NLS-Lyn were treated with sodium orthovanadate (Na3VO4). Endogenous Ku70 was immunoprecipitated (IP) with anti-Ku70 antibody. Western
blotting was performed with the indicated antibodies.
Figure 004. A sequence of Ku70 phosphopeptide detected by phosphoproteomic analysis.
Figure 005. Quantification for UV-induced apoptosis.
HeLa S3 cells were treated with UV light and cultured for 12 h. After fixation, cells were stained with propidium iodide for DNA. Arrowheads indicate cells showing apoptotic DNA fragmentation/condensation.
Figure 006. Role of Ku70 phosphorylation at Tyr-530 in suppression of apoptosis.
HeLa S3/TR cells inducibly expressing myc-Ku70-wt, myc-Ku70-YF, or myc-Ku70-YE were incubated with doxycycline for 24 h, and then irradiated with 100 J/m2 UV. At 12 h after UV
irradiation, cells were fixed and doubly stained with anti-myc antibody and propidium iodide. Arrowheads indicate cells showing apoptotic DNA fragmentation/condensation.
Cell line: None myc-Ku70-wt myc-Ku70-YF myc-Ku70-YE
m
yc
-K
u
70
D
N
A
Figure 007. Role of Ku70 phosphorylation at Tyr-530 in acetylation.
HeLa S3/TR cells inducibly expressing myc-Ku70-wt or myc-Ku70-YE were incubated with doxycycline for 24 h, and then irradiated with 100 J/m2 UV. At 12 h after UV irradiation,
acetylated Ku70-wt or Ku70-YE was immunoprecipitated with anti-acetylated lysine antibody. Western blotting was performed with anti-myc and anti-α-tubulin antibodies.
Tables
Table 1. Primer list for proteins
Table 2. Primer list for Ku70 mutants
Table 3. Nucleotides list for shRNA
shRNA target sequence (5'-3') Ku70 shRNA-3’UTR GAAGAGTCTACCCGACATAAG Ku70 shRNA-CDS GATGAGTCATAAGAGGATCAT
Lyn shRNA GGACAGAGGTTTCAAACTA (Morinaga et al., 2014) Fyn shRNA GGAAGAGCTCTGAAATTAC (Morinaga et al., 2014) c-Src shRNA GCTCCAGATTGTCAACAAC (Sasaki et al., 2014)
Forward primer (5'-3') Reverse primer (5'-3')
Ku70-YF TGGATGAGTTTAAGGAACTAGTTTTCCCAC CAGATTACAATCCTGAAGG TGTAATCTGGTGGGAAAACTAGTTCCTTAA ACTCATCCACCAAGGAGCC Ku70-YE TGGATGAGTTTAAGGAGCTTGTTGAGCCTCC GGATTACAATCCTGAAGG TGTAATCTGGTGGCTCAACAAGCTCTTTAA ACTCATCCACCAAGGAGCC Ku70-(shRNA)-resistant TGATGTCCAATTTAAAATGTCTCACAAACG AATCATGCTGTTCACCAATG ACAGCATGATTCGTTTGTGAGACATTTTAA ATTGGACATCACTAAAGAGG
Forward primer (5'-3') Reverse primer (5'-3')
Ku70 GAGTCGGATCCTTAAGCAGTGGAATGTCAG GGTGGGAGTC GAATAGGGCCCATCTCGAGTCAGTCCTGGA AGTGCTTGG Bax ACTGAATTCAGTGGAGGTATGGACGGGTCC GGGGAGCAGCCCAG CAGTCTCGAGTCAGCCCATCTTCTTCCAGAT GGTGAGTGAGGC
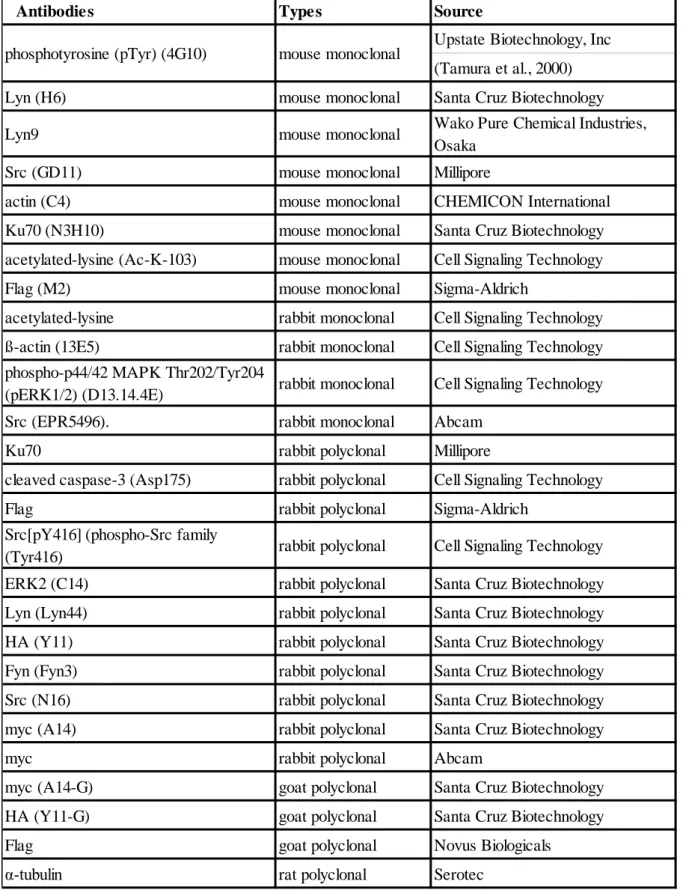
Table 4. Antibodies
Antibodies Types Source
Upstate Biotechnology, Inc (Tamura et al., 2000) Lyn (H6) mouse monoclonal Santa Cruz Biotechnology Lyn9 mouse monoclonal Wako Pure Chemical Industries,
Osaka Src (GD11) mouse monoclonal Millipore
actin (C4) mouse monoclonal CHEMICON International Ku70 (N3H10) mouse monoclonal Santa Cruz Biotechnology acetylated-lysine (Ac-K-103) mouse monoclonal Cell Signaling Technology Flag (M2) mouse monoclonal Sigma-Aldrich
acetylated-lysine rabbit monoclonal Cell Signaling Technology ß-actin (13E5) rabbit monoclonal Cell Signaling Technology phospho-p44/42 MAPK Thr202/Tyr204
(pERK1/2) (D13.14.4E) rabbit monoclonal Cell Signaling Technology Src (EPR5496). rabbit monoclonal Abcam
Ku70 rabbit polyclonal Millipore
cleaved caspase-3 (Asp175) rabbit polyclonal Cell Signaling Technology Flag rabbit polyclonal Sigma-Aldrich
Src[pY416](phospho-Src family
(Tyr416) rabbit polyclonal Cell Signaling Technology ERK2 (C14) rabbit polyclonal Santa Cruz Biotechnology Lyn (Lyn44) rabbit polyclonal Santa Cruz Biotechnology HA (Y11) rabbit polyclonal Santa Cruz Biotechnology Fyn (Fyn3) rabbit polyclonal Santa Cruz Biotechnology Src (N16) rabbit polyclonal Santa Cruz Biotechnology myc (A14) rabbit polyclonal Santa Cruz Biotechnology myc rabbit polyclonal Abcam
myc (A14-G) goat polyclonal Santa Cruz Biotechnology HA (Y11-G) goat polyclonal Santa Cruz Biotechnology Flag goat polyclonal Novus Biologicals
α-tubulin rat polyclonal Serotec phosphotyrosine (pTyr) (4G10) mouse monoclonal
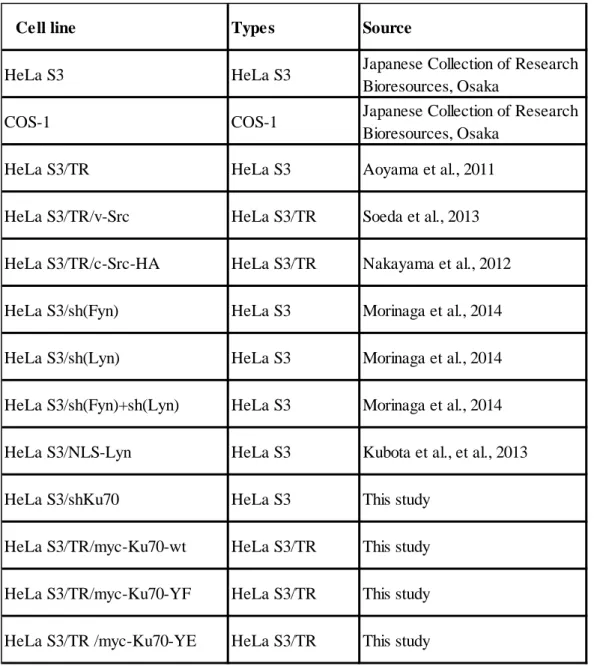
Table 5. Cell line used in this study
Cell line Types Source
HeLa S3 HeLa S3 Japanese Collection of Research Bioresources, Osaka
COS-1 COS-1 Japanese Collection of Research Bioresources, Osaka
HeLa S3/TR HeLa S3 Aoyama et al., 2011 HeLa S3/TR/v-Src HeLa S3/TR Soeda et al., 2013 HeLa S3/TR/c-Src-HA HeLa S3/TR Nakayama et al., 2012 HeLa S3/sh(Fyn) HeLa S3 Morinaga et al., 2014 HeLa S3/sh(Lyn) HeLa S3 Morinaga et al., 2014 HeLa S3/sh(Fyn)+sh(Lyn) HeLa S3 Morinaga et al., 2014 HeLa S3/NLS-Lyn HeLa S3 Kubota et al., et al., 2013 HeLa S3/shKu70 HeLa S3 This study
HeLa S3/TR/myc-Ku70-wt HeLa S3/TR This study HeLa S3/TR/myc-Ku70-YF HeLa S3/TR This study HeLa S3/TR /myc-Ku70-YE HeLa S3/TR This study
Main Publication
1. Morii, M., Kubota, S., Honda, T., Yuki, R., Morinaga, T., Kuga, T., Tomonaga, T., Yamaguchi, N., and Yamaguchi, N. Src acts as an effector for Ku70-dependent suppression of apoptosis through phosphorylation of Ku70 at Tyr-530. J. Biol. Chem., 292: 1648-1665, 2017.
Examiners
This thesis was reviewed by the following thesis committee members authorized by Graduate School of Pharmaceutical Sciences, Chiba University
Professor Toshihiko Murayama, Ph.D.
(Graduate School of Pharmaceutical Sciences, Chiba University) Chief examiner
Professor Motoyuki Itoh, Ph.D.
(Graduate School of Pharmaceutical Sciences, Chiba University) Professor Atsushi Iwama, M.D., Ph.D.
(Graduate School of Medicie, Chiba University) Professor Ashfaq Mahmood, Ph.D.
Acknowledgements
I would like to express my appreciation to my thesis adviser Professor Naoto Yamaguchi for all his invaluable guidance, suggestions, supervision, kindness and continuous encouragement during these years.
I also would like to thank my committee members: Professor Toshihiko Murayama, Professor Motoyuki Itho, Professor Atsushi Iwama, and Professor Ashfaq Mahmood for their constructive advice and thoughtful discussions.
I am grateful to Dr. Noritaka Yamaguchi, Dr. Yasunori Fukumoto, and Dr. Yuji Nakayama for their technical expertise, helpful suggestions, and discussions.
I also want to thank all my labmates for their support, encouragement, friendship, wisdom, and patience.
We are indebted to Dr. Donald J. Fujita, Dr. Stéphane A. Laporte, Dr. Tadashi Yamamoto, Dr. Toshiki Tamura, Dr. Takayuki Yoshimoto, and Dr. Hiroyuki Miyoshi for invaluable plasmids and antibodies.