1. はじめに
1990 年代初頭以降,生物多様性の重要性に対す る認識が高まっており,生物多様性条約(生物の多 様性に関する条約)の締結を初めとして世界的な取 り組みが進む中で,日本では 1995 年 10 月に最初 の生物多様性国家戦略が策定された(環境省 2012). その後 2008 年の生物多様性基本法の制定,2010 年 の生物多様性条約締約国会議の名古屋での開催を経 て,2012 年には生物多様性基本法に基づく第 5 次 となる生物多様性国家戦略 2012-2020 が策定された (環境省 2012). 生物多様性国家戦略 2012-2020 では国内の環境を 大きく 7 地域に分類しており,里地里山・田園地域 はそのうちのひとつである.農林業など人の手が加 えられることによって特有の自然環境が形成されて きた里地里山・田園地域に生息する種は,かつては 人々の生活に基づく人為的攪乱を受けて結果的に好 適な環境が創出されたために維持されてきた(日本 生態学会 2014).しかし,生活様式の変化にともなっヒシクイの採食行動に水位が与える影響
田尻浩伸
1,2 1公益財団法人日本野鳥の会保全プロジェクト推進室,2片野鴨池自然研究所 要 旨 継続して人の手が加えられてきた里地里山環境では,今後も生物多様性を維持するには保全対象種 の生態を考慮した管理を行なうことが重要である.筆者は,農業用のため池およびカモ猟の猟場とし て利用されている石川県加賀市の片野鴨池において,その鳥類相を特徴付ける希少種ヒシクイ(亜種 オオヒシクイAnser fabalis middendorffii)の採食行動に水位が与える影響を調べ,その結果に基づいた 水位管理手法を検討した.調査期間中,設置した水位計の値(基準水位)は20 cmから65 cmの範囲で 変動した.ヒシクイの採食行動は水深の影響を受けており,採食場所の水深が80 cmより浅い時には採 食個体の割合は0から1までの幅広い値をとっていたが,それより深いと割合は低下した.水位が浅い 時にはヒシを採食する個体と抽水植物を採食する個体のどちらの割合も0から1までの幅広い値をとっ ていたが,ヒシを採食していた個体の割合は基準水位が上昇するにつれて低下し,一方,抽水植物を 採食していた個体の割合は基準水位が高くなるにつれて上昇した.以上より,ヒシクイの採食環境と 食物選択性の幅を維持するためには,浅い水位を維持するように管理するとよいと考えられた. キーワード:ヒシクイ,片野鴨池,採食行動,水位管理,水深 て加えられる人の手は減少し,全国的に里地里山・ 田園地域の環境は変化しつつあり,その特有な環境 に生息する種の維持を目的とした取り組みが行な われるようになっている(日本生態学会 2014).こ のような状況を受けて,生物多様性国家戦略 2012-2020 では里地里山・田園地域の望ましい姿のひと つとして,「ため池や畦が豊かな生物多様性が保た れるように管理され」ることと指摘されている(環 境省 2012). 石川県加賀市にある片野鴨池は,国指定天然記 念物のマガンAnser albifrons やヒシクイ Ans. fabalis, 絶滅危惧 II 類に指定されているトモエガモ Anas formosa が利用することを特徴とする西日本最大級 のガンカモ類の越冬地で,これまでに 34 種類のガ ンカモ類が記録されている(田尻 2009).江戸時代 から水田耕作やカモ猟の猟場として利用されており, 現在でも春から夏にかけては周辺水田に水を供給す るため池として,また冬には伝統猟法である坂網猟 (投げ網猟)の猟場とするため,経験に基づいた植 田尻浩伸 [email protected] (2020 年 6 月 19 日受付,2020 年 12 月 20 日受理)生や水位の管理が行なわれている典型的な里地里山 環境である. ガンカモ類による片野鴨池の利用形態は種によっ て異なる.トモエガモやマガモAna. platyrhynchos などの水面採食性カモ類はおもに日中のねぐらとし て利用しながら,日中でも午前中を中心にしばしば 採食を行なう(田尻 2009,2018).マガンやコハク チョウCygnus columbianus はおもに夜間のねぐらと して利用し,日中でも滞在していれば採食も行なっ ている(田尻 2009).ヒシクイの利用形態は十分に 解明されていないが,ねぐらまたは採食地として終 日利用している. 片野鴨池の生物多様性を維持していくためには, それぞれの種の利用形態に則した管理を行なうこと が重要である.そのためには,科学的根拠に基づい て実施の効果を予測した上で管理手法を決定し,さ らに実施結果の評価を行なって管理手法に反映させ ることが有効である.しかし,これまでの伝統的管 理手法は個人の経験に基づく手法であるため,管理 者の経験の多寡や質の違いなど管理者個人の資質の 影響を強く受け,長期に渡って適切な管理を継続す るには課題がある.その解決のため,これまでに水 面採食性カモ類の採食行動に基づいた水位管理手法 が提案されている(田尻 2018).本研究では,片野 鴨池の鳥類相の特徴のひとつであり,水面採食性カ モ類と同様に片野鴨池を採食地としても利用してい るヒシクイについて,その採食行動を観察し,調査 結果に基づいた水位管理手法を検討する.
2. 調査地と方法
調査は石川県加賀市の片野鴨池(36°19′N,136° 17′E)で行なった.片野鴨池は面積約 10 ha の天 然の池沼で,周囲を標高 40 m 程度の丘に囲まれた すり鉢状の地形となっている.池の西部には水門が あり,おもにため池としての貯水量の調節に利用さ れている.ただし,本調査期間中には水門の操作は 行なわれなかった.調査は,片野鴨池を 38 区画に 分割して設定した 20 m × 20 m の方形区を基準と して行なった(Fig. 1). 片野鴨池の池底は東から西に向かって深くなる 非常に緩やかな傾斜があり,浅い方から順にヨシPhragmites australis,ウキヤガラ Scirpus yagara,マ コモZizania latifolia などの抽水植物が生育する.抽
水植物が生育しない水深になると,オニビシTrapa
natans var. japonica とコオニビシ T. n. var. pumila が 生育する(田尻ら 2014).また,池の東部と南東 部の一部には,環境教育の場およびガンカモ類の 採食地とするため,水田環境が維持されイネOryza sativa が作付けされている(日本野鳥の会 2011). 年による若干の変動はあるものの,概観すればFig. 1 に示した方形区の 1-3 列は抽水植物の分布域,5-7 列はヒシの分布域で,4 列目は両者の分布が重なる 範囲であった. 調査対象としたヒシクイは国指定天然記念物 (1971 年指定),絶滅危惧 II 類(亜種ヒシクイ Ans. f. serrirostris)または準絶滅危惧(亜種オオヒシク イAns. f. middendorffii)に指定されている(環境省 2019)希少な種である.片野鴨池で越冬するヒシク イは亜種ヒシクイと亜種オオヒシクイからなるが, 亜種オオヒシクイが大部分を占める(石川県 2009). 亜種オオヒシクイは,越冬地ではマコモの根茎やヒ シ類の種子などを採食する(呉地 2006,池内 2010). なお,簡便のため,以降はヒシクイと記載した場合 は亜種オオヒシクイを指すものとする. 調査は 2010 / 2011 年(以降 2010 年),2011 / 2012 年( 以 降 2011 年 ) と 2018 / 2019 年( 以 降 2018 年)の 12 月から 3 月にかけて,それぞれ 30 日, 56 日,10 日行なった. 水位の記録は,調査日に 1 回,方形区 A4 と B4 の間に設置した水位計の値を基準水位として 1 cm 単位で目測し記録した(Fig. 1).この水位計には 20 cm ごとに目盛をつけ,水底を 0 cm として設置 した.方形区ごとの水深は,2011 年 4 月 7 日に方 形区ごとに 5 か所ずつ測定した水深の平均と,同日 の基準水位との差異をもとに,調査日ごとに算出し た. 水位がヒシクイの行動に及ぼす影響を明らかにす るため,ヒシクイの行動調査をヒシクイが活発に採 食を行なう午前 9 時から 11 時頃(田尻浩伸未発表 データ)にかけて 1 日あたり 1 回から 4 回行ない, 各越冬期にそれぞれのべ 56 回,78 回,12 回実施し た.同一日に複数回の調査を行なう場合は,調査回
Fig. 1 調査地.図中の正方形は調査用に設定した方形区を示し,各方形区の名称は列番号(1-7)と行
番号(A-F)の組合せで表す.星印は基準水位測定用に設置した水位計の位置を示し,Ob. は観察地点で ある加賀市鴨池観察館を表す.方形区は基準水位 20 cm 時の水深に応じて 5 段階に色分けした.図は国 土地理院の地理院地図(タイル)を加工して著者が作成.
Fig. 1 Map of the study site. Squares on the map indicate the quadrats set up for the study. The ID of each quadrat
was expressed as a combination of the column number (1-7) and row letter (A-F). The star indicates the location of the standard water-level gauge and “Ob.” indicates the observation point (Kaga City Kamoike Observation Center). Quadrats were divided into five levels according to the water depth when the water level is 20 cm and the levels were expressed by the shading. This figure was created by the author based on data from the Geospatial Information Authority of Japan. ごとの独立性を高めるため,1 回の調査終了後にヒ シクイが調査範囲から一旦飛去した後に再び戻って きてから次の調査を行なうなどの配慮を行なった. 観察時のヒシクイの行動の記録は,方形区を単位 として行なった(Fig. 1).ただし,2010 年の調査 では方形区ごとの記録を行なわず,38 区画に相当 する範囲を利用していたヒシクイをまとめて記録し た.方形区の境界の一部には杭を設置し,杭を基準 として方形区を認識した.なお,ヒシクイは 1 列目 のメッシュと鴨池観察館のあいだにある水田もしば しば採食に利用することから,観察館の存在はヒシ クイに影響を与えないか,与えても無視できる軽度 の影響と考えられる. 調査には 7 倍の双眼鏡と 20 倍の望遠鏡をもちい, 調査地の東岸にある加賀市鴨池観察館内から方形区 ごとに走査サンプリング法(Altmann 1974)をもち いて行なった.ヒシクイは採食している個体とそれ 以外の行動をとっている個体に分けて数えた.さら に,採食している個体は,嘴にくわえたりついばん だりしている食物の種を望遠鏡で目視観察し,ヒシ を採食している個体,抽水植物を採食している個体 と,それ以外を採食している個体の 3 通りに分けて 記録した.ヒシはオニビシとコオニビシを区別せ ずヒシとした.抽水植物は,マコモ,ウキヤガラ, イネとした.それ以外の食物には,ハスNelumbo nucifera の種子のほか,種を識別できなかった食物 をまとめた.なお,これ以降,採食している個体を 採食個体,ヒシを採食している個体をヒシ採食個体, 抽水植物を採食している個体を抽水植物採食個体, それ以外をその他採食個体と記述する. ヒシクイの採食行動に水位が与える影響を検討す るため,調査回ごとの採食個体の割合,ヒシ採食個 体の割合,抽水植物採食個体の割合のそれぞれを目 的変数とし,調査日の基準水位,12 月 1 日を 1 と した場合の調査日までの経過日数を固定効果,調査 年と調査ごとに割り当てた ID 番号をランダム効果 とし,誤差構造は二項分布,リンク関数を logit と
する 3 種類の一般化線形混合モデル(GLMM)を 構築して効果を検討した.ここで調査回ごとの割 合とは,38 区画に対する 1 回の走査サンプリング で記録されたヒシクイ全体に占める採食個体の割合, ヒシ採食個体の割合および抽水植物採食個体の割合 を指す.なお,本研究は 12 月から 3 月にかけて行 なったので,目的変数に経過日数が与える影響を考 慮するため,経過日数を説明変数に加えた.構築し た GLMM は AIC(赤池情報量基準)に基づくモデ ル選択を行ない,AIC がもっとも小さいモデルを当 てはまりの良いモデルと判断し,説明変数の効果を 検討した.これらの解析は統計ソフト R ver.3.5.2(R Core Team 2018)とパッケージ glmmADMB(Skaug et al. 2016)に含まれる関数 glmmadmb をもちいて 行なった.
3. 結果
3.1. 片野鴨池の基準水位の季節変化と方形区
ごとの水深
基準水位の季節変化は年によって異なっていた (Fig. 2).2010 年,2011 年には調査期間中に増減が 見られた.2010 年は 12 月末をピークに 30 cm から 65 cm の間で変動した.2011 年には 1 月中旬をピー クに 20 cm から 60 cm の間で変動した.2018 年は 12 月中旬から 2 月下旬まで 25 cm で一定していた. 調査時の方形区で記録された水深の平均は,2010 年,2018 年 の 順 に 44.3±32.9 cm( 平 均±SD, 範 Fig. 2 基準水位の季節変化.Fig. 2 Fluctuation of standard water level from December 1st in
each study period. The water level was measured using a standard water-level gauge.
Fig. 3 方形区ごとの採食個体の割合と方形区の水深の関係.
採食個体の割合は,水深 80 cm までは幅広い値を取り,80 cm を超えると低下した.
Fig. 3 Relationship between the proportion of foraging Bean
geese and water depth in each quadrat at the study site. When the water depth were below 80 cm, the proportion of foraging geese ranged from 0 and 1 and when the water levels were above 80 cm, the proportion of them were decreased.
囲 0-142.0,n=2128),35.3±30.3 cm(範囲 0-102.0, n=380)であった.個別の方形区のうち,もっとも 水深が浅かったのは方形区 A1 で,調査期間中の 平 均 水 深 は 0.5±1.9 cm( 範 囲 0-9.2,n=56,2010 年)と 0 cm(n=10,2018 年)であった.もっとも 深かったのは方形区 D7 で,調査期間中の平均水深 が 111.4±13.8 cm(範囲 97.0-142.0,n=56,2010 年) と 102.0 cm(n=10,2018 年)であった.
3.2. 方形区ごとの水深と採食個体の割合の関
係
2011 年に行なった 78 回の調査で記録されたヒシ クイは,のべ 400 方形区における合計 5,749 羽であっ た.そのうち採食していた個体は,のべ 225 方形区 で記録された合計 1,678 羽であった.調査回ごとに 各方形区で求めた採食個体の割合は,0.32±0.36(平 均±SD,範囲 0-1.0,n=400)であった. 2018 年に行なった 12 回の調査で記録されたヒシ クイは,のべ 127 方形区における合計 1,653 羽であっ た.そのうち採食していた個体は,のべ 73 方形区 で記録された合計 462 羽であった.調査回ごとに各 方形区で算出した採食個体の割合は,0.30±0.34(平 均±SD,範囲 0-1.0,n=127)であった. ヒシクイの採食が記録された方形区の水深でもっFig. 4 採食個体の割合と基準水位との関係.実線は GLMM
による予測値を示す.採食個体の割合は,基準水位が高く なるにつれて低下すると予測された.
Fig. 4 Relationship between the proportion of foraging Bean
geese and the standard water level. Solid line indicates the proportion of foraging geese estimated by a generalized linear mixed model (see Table 1). Estimated proportion of foraging geese decreased as the water level increased.
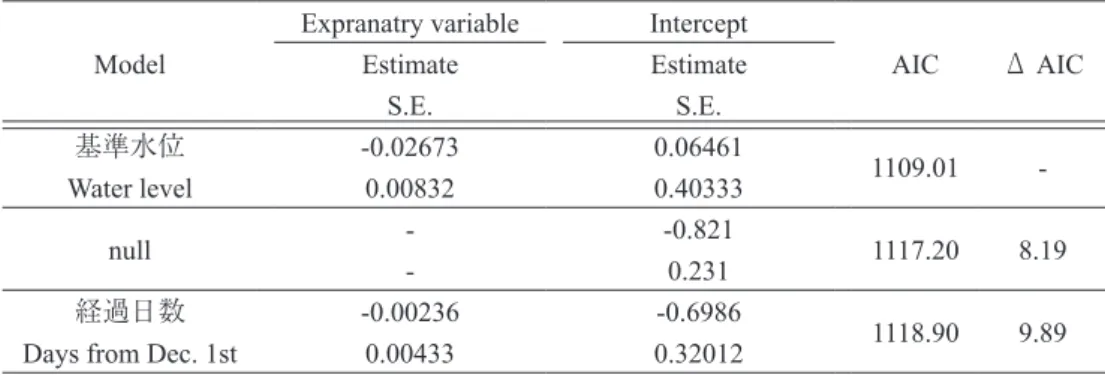
Table 1 GLMM をもちいた採食個体の割合に影響を与える要因の検討結果.上段の数字は推定値,
下段は標準誤差を表す.
Table 1 Generalized linear mixed model analysis of factors affecting the proportion of foraging Bean geese.
In each row, upper values indicate estimated coefficients and lower values indicate standard errors (S.E.). とも深かったのは,2011 年の調査では 2012 年 1 月 6 日の方形区 E6 の水深 93.1 cm で,13 羽のヒシク イのうち 3 羽(採食個体の割合 0.23)がマコモを採 食していた.2018 年の調査では 2018 年 12 月 17 日 の方形区 D6 の水深 82.8 cm で,3 羽のヒシクイが ヒシを採食していた(採食個体の割合 1.0). 調査回ごとに各方形区で算出した採食個体の割 合と,調査時の方形区の水深の関係をFig. 3 に示し た.2011 年,2018 年ともに類似した傾向が認めら れ,方形区の水深が 0 cm からおよそ 80 cm までの 範囲では,採食個体の割合は 0 から 1 までのさまざ まな値をとった.一方,およそ 80 cm を超えると採 食個体の割合の最大値が低下し,100 cm を超える と 0 となった.
3.3. 基準水位とヒシクイの行動
つぎに,調査回ごとに,調査対象とした 38 方形 区全体で記録されたヒシクイについて述べる. 2010 年には,採食個体は 56 回すべての調査で 記録され,その平均個体数は 25.5±13.8 羽(範囲 4-63)であった.調査回ごとの採食個体の割合は平 均 0.43 ± 0.19(範囲 0.07-0.94,n=56)であった. 2011 年には,採食個体は 78 回中 70 回の調査で 記録され,その平均個体数は 21.5 ± 23.2 羽(範囲 0-117,n=70)であった.調査回ごとの採食個体の 割合は平均 0.30±0.22(範囲 0-1.0,n=70)であった. 2018 年には,採食個体は 12 回中 11 回の調査で 記録され,その平均個体数は 37.6±21.3 羽(範囲 0-69)であった.調査回ごとに記録されたヒシクExpranatry variable Intercept
Model Estimate Estimate AIC Δ AIC
S.E. S.E. 基準水位 -0.02673 0.06461 1109.01 -Water level 0.00832 0.40333 null - -0.821 1117.20 8.19 - 0.231 経過日数 -0.00236 -0.6986 1118.90 9.89 Days from Dec. 1st 0.00433 0.32012
イに占める採食個体の割合は平均 0.29±0.17(範囲 0-0.68,n=12)であった. 調査回ごとの採食個体の割合を目的変数として, GLMM を構築して説明変数である基準水位と経過 日数の効果を調べた(Table 1).なお,説明変数間 の相関が高かったので(スピアマンの順位相関係数 rho=-0.692,P<0.001),ふたつの変数を同時に含む フルモデルは構築せず,それぞれの変数のみからな るモデルと変数をひとつも含まない null モデルを 構築し,AIC 値で効果の比較を行なった. 説明変数が基準水位のみからなる GLMM の AIC 値は 1109.01 でもっとも小さかった.null モデルの AIC 値は 1117.2,経過日数のみからなるモデルの
AIC 値は 1118.9 であった.採食個体の割合に影響 を与える変数として基準水位が選択され,基準水位 は採食個体の割合に負の影響を与えていた(Fig. 4).
3.4. 基準水位とヒシクイの食物
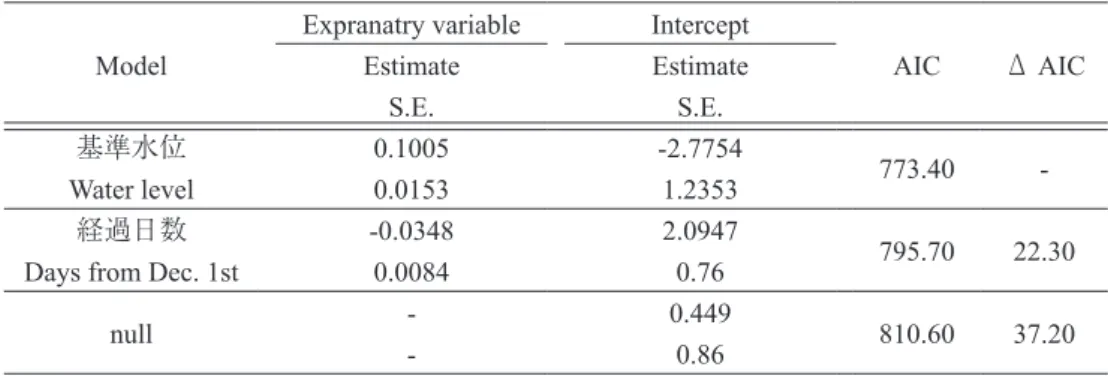
つづいて,調査回ごとに調査対象とした 38 方形 区全体で記録されたヒシ採食個体,抽水植物採食個 体について述べる. 2010 年には,採食個体が観察された 56 回の調査 回におけるヒシ採食個体数の平均は 13.2 ± 8.5 羽 (範囲 0-41,n=56),抽水植物採食個体数の平均は 12.3±11.5 羽(範囲 0-43,n=56)であった.その他 採食個体は記録されなかった.採食個体に占める ヒシ採食個体の割合は平均 0.56 ± 0.28(範囲 0-1.0, n=56),抽水植物採食個体の割合は平均 0.44 ± 0.28 (範囲 0-1.0,n=56)であった. 2011 年には,採食個体が記録された 70 回の調査 回におけるヒシ採食個体数の平均は 12.3±13.4 羽 (範囲 0-61,n=70),抽水植物採食個体数の平均は 11.4±17.3 羽(範囲 0-93,n=70),その他採食個体 数の平均は 0.2±1.3 羽(範囲 0-10,n=70)であった. 採食個体に占めるヒシ採食個体,抽水植物採食個体, その他採食個体の割合は,それぞれ平均 0.62±0.37 (範囲 0-1.0,n=70), 0.36±0.35(範囲 0-1.0,n=70), 0.01±0.07(範囲 0-0.53,n=70)であった. 2018 年には,採食個体が記録された 11 回の調査 におけるヒシ採食個体数の平均は 4.8±4.5 羽(範囲 0-13),抽水植物採食個体数の平均は 36.2±18.6 羽 (範囲 0-69)であった.採食個体に占めるヒシ採食 個体の割合は平均 0.12±0.12(範囲 0-0.39,n=11), 抽水植物採食個体の割合は平均 0.88±0.12(範囲 0.61-1.0,n=11)であった. Table 3 GLMM をもちいた抽水植物採食個体の割合に影響を与える要因の検討結果.上段の数字 は推定値,下段は標準誤差を表す.Table 3 Generalized linear mixed model analysis of factors affecting the proportion of rhizomes of
emerging plants foraging Bean geese. In each row, upper values indicate estimated coefficients and lower values indicate standard errors (S.E.).
Table 2 GLMM をもちいたヒシ採食個体の割合に影響を与える要因の検討結果.上段の数字は推
定値,下段は標準誤差を表す.
Table 2 Generalized linear mixed model analysis of factors affecting the proportion of Water chestnuts
foraging Bean geese. In each row, upper values indicate estimated coefficients and lower values indicate standard errors (S.E.).
Expranatry variable Intercept
Model Estimate Estimate AIC Δ AIC
S.E. S.E. 基準水位 0.1005 -2.7754 773.40 -Water level 0.0153 1.2353 経過日数 -0.0348 2.0947 795.70 22.30 Days from Dec. 1st 0.0084 0.76
null - 0.449 810.60 37.20
- 0.86
Expranatry variable Intercept
Model Estimate Estimate AIC Δ AIC
S.E. S.E. 基準水位 -0.1115 3.0842 769.60 -Water level 0.0159 1.2637 経過日数 0.0382 -2.293 795.60 26.00 Days from Dec. 1st 0.0088 0.7459
null - -0.628 1797.80 1028.20
Fig. 5 ヒシ採食個体の割合と基準水位との関係.実線は
GLMM による予測値を示す.ヒシ採食個体の割合は,基準 水位が高くなるにつれて低下すると予測された.
Fig. 5 Relationship between the proportion of foraging Bean
geese foraging the fruits of Water chestnuts Trapa natans and the standard water level. Solid line indicates the proportion of Water chestnut foraging geese estimated by a generalized linear mixed model (see Table 2). Estimated proportion of Water chestnut foraging geese decreased as the water level increased.
Fig. 6 抽水植物採食個体の割合と基準水位との関係.実線
は GLMM による予測値を示す.抽水植物採食個体の割合は, 基準水位が高くなるにつれて高まると予測された.
Fig. 6 Relationship between the proportion of foraging Bean
geese foraging the rhizomes of emerging plants and the standard water level. Solid line indicates the proportion of rhizomes foraging geese estimated by a generalized linear mixed model (see Table 3). Estimated proportion of rhizomes foraging geese increased as the water level increased.
調査回ごとのヒシ採食個体の割合および抽水植物 採食個体の割合を目的変数として,GLMM を構築 して説明変数である基準水位と経過日数の効果を調 べた(Table 2).なお,説明変数間の相関が高かっ たので(スピアマンの順位相関係数 rho=-0.666, P<0.001),ふたつの変数を同時に含むフルモデルは 構築せず,それぞれの変数のみからなるモデルと変 数をひとつも含まない null モデルを構築し,AIC 値 で効果の比較を行なった. ヒシ採食個体の割合を目的変数とする GLMM の AIC 値は,基準水位を含むモデルが 769.6 でもっ とも小さく,ついで経過日数を含むモデルの 795.6, null モデルの 812.3 であった.ヒシ採食個体の割合 に影響する変数として基準水位が選択され,基準水 位はヒシ採食個体の割合に負の影響を与えていた (Fig. 5). 抽 水 植 物 採 食 個 体 の 割 合 を 目 的 変 数 と す る GLMM の AIC 値は,基準水位を含むモデルが 773.4 でもっとも小さく,ついで経過日数を含むモデルの 795.7,null モデルの 810.6 であった.抽水植物採食 個体の割合に影響を与える変数として基準水位が選 択され,基準水位は抽水採食個体の割合に正の影響 を与えていた(Fig. 6).
4. 考察
4.1. ヒシクイが採食することができる水深
採食の際,ヒシクイを含むガンカモ類は,潜水採 食性の種を除けば,水深に合わせてさまざまな姿 勢で採食を行なう(del Hoyo et al. 1992,Hughes and Green 2005).採食が可能な水深は種によって異な り,一般に大型種ほど深い水深に対応できる(Green 1998,Hughes and Green 2005).本研究では,ヒシクイの採食個体の割合は水深 80 cm 程度まではさまざまな値を取り,およそ 80 cm を超えると採食個体の割合の最大値が低下し て 100 cm を超えると 0 となった.また,採食が記 録された方形区の最大水深は 2011 年では 93.1 cm, 2018 年では 82.8 cm であった. ヒシクイがヒシを採食する場合,水底に沈んでい る種子を食物とすることから,嘴が水底に到達し なくては採食することができない.2011 年にヒシ を採食するヒシクイが記録された方形区の水深は 91.9 cm がもっとも深く,ついで 88.9 cm,86.9 cm, 85.6 cm と続いた.2018 年では 82.8 cm が最も深く, 68.7 cm,68.6 cm と続いた.したがって,80 cm か ら 90 cm ほどの水深が,ヒシクイが採食を行なうこ とができるもっとも深い水深であると考えられる. なお,加賀市鴨池観察館所蔵のヒシクイの剥製をも
ちいて嘴の先端からかかとまでの長さを脊椎に沿っ て測定したところ,およそ 80 cm(田尻浩伸未発表 データ)であった.倒立採食を行なうヒシクイは勢 いをつけて倒立したり,倒立後に足をかいてより深 い水深に到達しようとするので,水深 90 cm で倒立 採食を行なった場合,水底に到達することが十分可 能であると考えられる.
4.2. ヒシクイの採食に基準水位が与える影響
片野鴨池の基準水位の高さは,ヒシクイの採食個 体の割合に負の影響を与えていた.また,方形区の 水位が 80 cm を超えると採食個体の割合の最大値が 低下していた.これらは,ヒシクイの採食環境を創 出するためには,片野鴨池内の採食場所の水深が 80 cm を超えないように管理すべきであることを示 唆している. 最適採餌理論では,動物の食物選択や採食地選択 は採食効率が最大になるように調節されていると考 えられており(たとえば Charnov 1976a,1976b,嶋 田ら 2006),食物資源の現存量,エネルギー量や含 有する栄養素(Amano et al. 2004,2006,Baldassarre and Bolen 2006),利用しやすさに影響を与える水深 や積雪深(嶋田・植田 2006,植田 2007,嶋田 2010, 田尻ら 2013),採食に割り当てることができる時間 (Amano et al. 2004),採食に必要となるエネルギー 支 出(Nolet et al. 2001,2006), 捕 食 リ ス ク(Fox and Kahlert 2000,Hughes and Green 2005)などの影 響を受ける. 基準水位は,ヒシ採食個体の割合には負の影響, 抽水植物採食個体の割合には正の影響を与えていた. 基準水位が低い場合には,どちらも 0-1.0 のあいだ のさまざまな値をとっていたが,基準水位が高くな るにつれてヒシ採食個体の割合は低下し,抽水植物 採食個体の割合が増加した.これは基準水位の上昇 にともなってヒシが生育している比較的沖合に位置 する範囲での採食効率が低下したり,ヒシクイが採 食可能な水深 80-90 cm を超えたりしたため,水深 が浅く抽水植物が生育する方形区を中心に採食する ようになったことによると考えられる. したがって,採食個体の割合の幅に加えて食物選 択の幅を広げる意味においても,基準水位を低く保 つことが有効であると考えられる. 本研究では,前述した水深以外の採食に影響を与 える要因についての調査を行なわなかったので,そ れらの比較からヒシクイの食物選択を検討すること ができなかったが,既存のデータが存在する捕食者 と積雪深の影響について検討を行なった. 片野鴨池にはヒシクイの捕食者としてクマタカ Nisaetus nipalensis が越冬し,頻度は高くないものの マガンやヒシクイを捕食する(中川ら 2005)ので, クマタカの存在はヒシクイの採食方法の選択に影響 を与えているかもしれない. ガンカモ類では,倒立採食のような目が水面下に 沈む採食方法は,濾し取りやついばみなど目が水面 上に出ている他の採食方法に比べて,捕食者への対 応にコストがかかることが知られている(Guillemain et al.2000,2001,2007).調査を行なった 12 月か ら 3 月におけるクマタカの出現頻度は,2010 年が 9.9%,2011 年が 13.9%,2018 年が 23.1%(加賀市 鴨池観察館未発表データ)で,ヒシ採食個体の割合 が低かった 2018 年でもっとも高かった. したがって,2018 年には捕食者への対応として, 倒立採食を必要とするヒシの採食を避け,より水深 の浅いところで採食できる抽水植物を採食する傾向 にあったのかもしれない. 調査地にもっとも近いアメダス観測地点は福井県 地方気象台で,調査期間中の月ごとの最深積雪は, 2010 年は 119 cm(2011 年 1 月),2011 年は 61 cm(2012 年 2 月),2018 年は 14 cm(2019 年 1 月)であった (気象庁 a).水が存在すると,降った雪は地面が干 出している場所と比べて早く融けるので,2010 年, 2011 年は,基準水位が高い時期には採食しやすい 場所が広がったと考えられる.2010 年で基準水深 が深くなるほど抽水植物採食個体の割合が増加する 傾向が顕著だったのはそのためかもしれない.4.3. 片野鴨池の水位管理
本研究により,片野鴨池をヒシクイの採食地とし て管理する場合には水位を低く保つべきであること が明らかとなった.その際の基準水位は,採食個体 の割合がもっとも高かった 20 cm 程度が適当と考え られる.ただし,20 cm より浅い基準水位は本研究時には記録されなかったので,20 cm 未満における 採食個体の割合の把握と水位の評価が必要だろう. 秋から冬にかけて片野鴨池ではヒシクイ以外にも 希少種であるマガンやトモエガモ,狩猟の対象種と なっているマガモなども多数が越冬することから, ヒシクイ以外のガンカモ類の越冬環境にも配慮した 管理が求められる. 田尻(2018)は,水位と水面採食性カモ類の採食 行動の関係を調べ,片野鴨池の流出部にある水門の 管理を通じて初冬にはこれまでよりも 20 cm 水位を 低く設定し,季節の進行にともなって水門に堆積す る植物遺骸を利用して水位を徐々に上げ,越冬期終 盤にかけて岸近くに水際が来るように管理する手法 を提案した.この方法を採用すると,本研究でもち いた基準水位は越冬期初期に約 10 cm,終盤には 30 cm 程度となり,ヒシクイの採食環境創出に適した 基準水位である 20 cm を中心に前後 10 cm の範囲に 収まる.したがって,田尻(2018)が提案した管理 方法は,水面採食性カモ類とヒシクイそれぞれに適 した水位管理を両立させる妥当な管理方法と言える だろう.
謝 辞
2018 年の調査時には,加賀市鴨池観察館レン ジャーの皆さんに協力いただきました.記して感謝 申し上げます.引用文献
Altmann J (1974) Observational study of behavior: Sampling methods. Behaviour, 49(3-4), 227-267.
Amano T, Ushiyama K, Fujita G and Higuchi H (2004) Alleviating grazing damage by White-fronted geese: An optimal foraging approach. Journal of Applied Ecology, 41, 675-688.
Amano T, Ushiyama K, Fujita G and Higuchi H (2006) Foraging patch selection and departure by non-omniscient foragers: A field example in White-fronted geese. Ethology, 112, 544-553.
Baldassarre GA and Bolen EG (2006) Waterfowl ecology and management Second edition. Krieger Publishing Company. Charnov EL (1976a) Optimal foraging: Attack strategy of a
mantid. The American naturalist, 110, 141-151.
Charnov EL (1976b) Optimal foraging: The marginal value theorem. Theoretical population biology, 9,129-136. del Hoyo J, Elliott A and Sargatal J (eds) (1992) Handbook of
the Birds of the World. Vol. 1. Lynx Edicions.
Fox AD and Kahlert J (2000) Do moulting Greylag geese Anser anser forage in proximity to water in response to food availability and/or quality? Bird study, 47, 266-274. Green AJ (1998) Comparative feeding behaviour and
niche organization in a Mediterranean duck community. Canadian Journal of Zoology, 76(3), 500-507.
Guillemain M, Fritz H and Blais S (2000) Foraging methods can affect patch choice: an experimental study in mallard (Anas platyrhynchos). Behavioural Processes, 50, 123-129. Guillemain M, Duncan P, and Fritz H (2001) Switching to
a feeding method that obstructs vision increase head-up vigilance in ducks. Journal of Avian Biology, 32, 345-350. Guillemain M, Arzel C, Legagneux P, Elmberg J, Fritz H,
Lepley M, Pin C, Arnaud A and Massez G (2007) Predation risk constrains the plasticity of foraging behaviour in teals, Anas crecca: a flyway-level circumannual approach. Animal Behaviour, 73(5), 845-854.
Hughes B and Green AJ (2005) Feeding ecology. Ducks, geese and swans volume 1 (ed. Kear J), 27-56. Oxford University Press. 池内俊雄(2010)菱喰雁留多(ヒシクイカルタ)資料 編 オオヒシクイ Q&A.雁の里親友の会. 石川県(2009)改訂・石川県の絶滅のおそれのある野 生生物 いしかわレッドデータブック<動物編> 2009.石川県環境部自然保護課. 環境省(2012)生物多様性国家戦略 2012-2020 ~豊か な自然共生社会の実現に向けたロードマップ~.環 境省. 環 境 省(2019)【 鳥 類 】環 境 省 レ ッ ド リ ス ト 2019. <https://www.env.go.jp/press/files/jp/110615.pdf>( 参 照 2020 年 6 月 10 日) 気象庁 a, 過去の気象データ.<https://www.data.jma.go.jp/ obd/stats/etrn/index.php?prec_no=57&block_no=47616& year=2019&month=&day=&view=p1>( 参 照 2020 年 6 月 1 日) 呉地正行(2006)雁よ渡れ.どうぶつ社. 中川直之・田尻(山本)浩伸・大畑孝二(2005)片 野鴨池におけるクマタカの水鳥の補食例.Strix,23, 181-183. 日本生態学会(編)(2014)エコロジー講座 7 里山のこ れまでとこれから.日本生態学会. 日本野鳥の会(2011)平成 22 年度(2010 年度)加賀市 鴨池観察館年次報告書.日本野鳥の会.
Nolet BA, Fuld VN and van Rijswijk MEC (2006) Foraging costs and accessibility as determinants of giving-up densities in a swan-pondweed system. Oikos, 112, 353-362.
Nolet BA, Langevoord O, Bevan RM, Engelaar KR, Klaassen M, Mulder RJW and van Dijk S (2001) Spatial variation in tuber depletion by swans explained by differences in net intake rates. Ecology, 82(6), 1655-1667.
R Core Team (2018) R: A language and environment for statistical computing. R foundation for statistical
computing. 嶋田正和・山村則男・粕谷栄一・伊藤嘉明(2006)動 物生態学 新版.海游社. 嶋田哲郎(2010)気象条件にともなうヒシクイの短期 的移動.BirdResearch,6,S7-S11. 嶋田哲郎・植田健稔(2006)2005/06 年の寒波がガン カモ類の個体数変動に与えた影響.BirdResearch,2, A11-A17.
Skaug H, Fournier D, Nielsen A, Magnusson A and Bolker B (2016) Generalized Linear Mixed Models using 'AD Model Builder'. <http://glmmadmb.r-forge.r-project.org> (参照 2020 年 6 月 8 日)
田尻浩伸(2009)北陸地方におけるカモ群集の越冬生 態と保全に関する研究-特に絶滅危惧種トモエガモ
The effect of water level on the foraging behaviour of Bean geese
H
ironobuT
AJIRI1,21Preservation Projects, Wild Bird Society of Japan 2Katano-kamoike Institute of Nature
Abstract: In SATOYAMA environments that have been varied due to changes in the lifestyle of local people, the habitat of some species has also been altered. Therefore, to conserve the biodiversity of such environments, including Katano-kamoike pond where is managed as reservoir for rice cultivation and traditional hunting area, management plans that consider the ecology of target species are required. In the present study, I investigated the effect of water level on the foraging behavior of Bean geese Anser fabalis middendorffii at Katano-kamoike pond in Kaga City, central Japan; based on the results, I propose a water-level management plan. During the study periods, the water level at the site, which was measured with a standard water-level gauge, fluctuated between 20 and 65 cm. In general, the proportion of foraging Bean geese ranged from 0 and 1 when water depth was lower than 80 cm and those were low at higher water depth. In addition, the proportion of Bean geese foraging for the fruits of Water chestnuts Trapa natans decreased as the water level rose, while that of geese foraging for the rhizomes of emerging plants increased. These results were confirmed statistically using a generalized linear mixed model. Because the foraging behavior of Bean geese was affected by the water level at the study site, I suggest that it would be advisable to maintain a low water level in order to conserve the foraging grounds and the flexibility in the food selection of Bean geese.
Key words: Anser fabalis middendrffii, Bean goose, foraging behaviour, water depth, water level management について-.平成 21 年度博士論文.金沢大学. 田尻浩伸(2018)水面採食性カモ類の採食方法と水深 の関係から検討する池沼の水位管理手法-ラムサー ル条約湿地・片野鴨池を例に-.Strix,34,37-57. 田尻浩伸・松本潤慶・田米希久代・中野夕紀子(2014) ラムサール条約湿地・片野鴨池の溶存酸素量の経時 変化と水生植物の関係.伊豆沼・内沼研究報告,8, 23-34. 田尻浩伸・櫻井佳明・組頭五十夫・大西五十二・鈴川文夫・ 田米希久代・山本芳夫(2013)風力発電施設周辺に おけるマガンの飛行コース選択と気象条件および採 食場所の位置の関係.Strix,29,1-16. 植田睦之(2007)ハクチョウ類やカモ類の越冬数に積 雪や気温がおよぼす影響.BirdResearch,3,A11-A18.