仁淀川下流域の砂疎堆にみられる植物群落*
石 川 慎 吾.・石 田 明 儀**
(理学部生物学科)
Plant communities on the bars in the lower course of the
Niyodo River, Shikoku*
Shingo ISHIKAWA and Akiyoshi ISHIDA**
(Department ofBiology,FacultyofScience)
Abstract
The plant communities developed on the bars were investigated in the lower course of
the Niyodo River where the river bed gradient is about 1 ‰and alluvial deposits are
composedmainly of gravels and coarse sand. Eight community types and two subtypes were
recognized "by the comparative study of the dominants and the floristic composition as
follows: 1) Polygonum九ydropφer comm., 2-a)p. hydropiper‘・-Polygonumthunberがi
commら2-b) DittoSalix graciltstylo-Pんragmites japonica type, 3) P. h:ydropiper-S
gracilistylacomm., 4) P. h:ydropiper-P.japonicacomm., 5) Miscan£hus sacchariflorus
comm・, 6) P.japoaicacomm., 7-a)Kummerouia stφulaceacomm., 7) Ditto.
P.μipo-nicatvpe,8)S.gracilistyla comm.1)t0 4) occupy wet habitats' along the stream. 7)
occurs in semi-dry flat places far from the stream. 5) is found on mesic places where
medium and fine sands are deposited by floods. 6) tends to develop on various habitats.
8) is restricted mainly to the places near the stream, and big mounds are constructed of
gravels and sands deposited by floods on and around the scrubs of this type.
`As the result of the sieving size analysis of the surface deposits of each stand, any
differences among those collected from the layers deeper than 2 cm are not recognized
except some stands of 5) and 6).
Four species groups are classified by means of cluster analysis based on interspecific
association, and are closely correlated with the community types O□)t0 3), 5), 7) and 8)
respectively. は じ め に 西南日本の河川は比較的小規模のものが多く,東北日本に見られるような平野の中をゆったり流 れる大きな河川は少ない(小出, 1970).従って氾濫原も狭く,そこに成立する植物群落も小規模 である.今回調査した仁淀川も,平野部を流れる流域をほとんど持たず,いわゆる急流河川の一つ である.このような河川の河床に発達する植物群落を扱った研究は少なく,猶原(1951)が東海地 方の富士川,天龍川,相模川などの数河川で行った研究のほか,西日本の河川では,瀬野川での波 田(1972),小瀬川での中西・関(1979)などがあるにすぎない. 急流河川では,特に洪水や増水による河床の変勁が激しく,そこに生育する植物もその物理的撹 *昭和58年度文部省特定研究経費(高知大学)の一部を使用した * * 1984年10月逝去
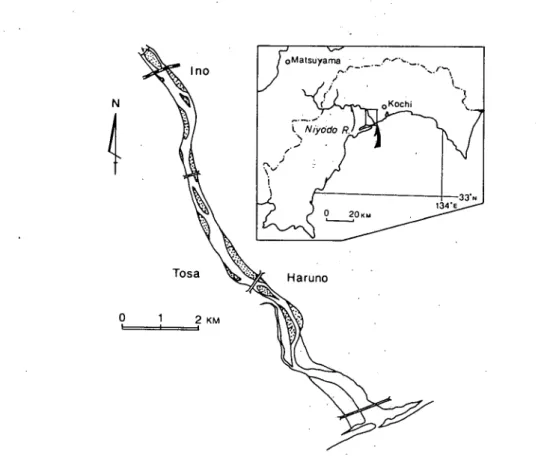
266 高知大学学術研究報告 第34巻 n985)自然科学 乱の影響を強く受けている.非常に卓越した流水による破壊作用のために,植生は貧弱であるが, このような変動の激しい環境に成立している植物群落は,その環境の変動にある部分では同調して 生活しているはずである.それがどのような生態的特性によるものなのか,そしてそこに生活する 植物たちがお互いにどのような関連を持ちつつ,植物社会を構成しているのかを研究することは, 遷移や変動といった,植物社会の持つ動的な側面の解明に少なからず役立つと考える.このような 視点から,仁淀川下流域の砂喋堆上にみられる植物群落について調査を行った.本報では,まず成 立している植物群落の総合的な把握,それぞれの群落と立地との対応関係,さらに主要構成種の個 体群相互の分布相関について検討した結果を報告する. なお,種名は大井(1975)に従ったが,帰化植物の一部は長田(1979)に従った. 調査地の概況 仁淀川は西日本の最高峰である石鎚山に発し,高知県の中央部を南東に貫流し,土佐市新居,春 野町仁野付近で土佐湾に注ぐ.四国では吉野川,四万十川につぐ第三位の長流で,全長125.9 km (うち面河川は49.3 km),流域面積は1585 、におよぶ.四国山地の中を流れる中流域では峡 谷を形成し,河床は狭い.吾川郡伊野町より下流域になると,河床には大小の砂榛堆が形成され, 長いものでは2kmに達するものもみられる(Figバ).それらの砂疎堆上に発達する植物群落は小 N4ト
Fig. 1 Map showing the river course investigated. Dotted areas indicate the bars studied. .、 し● ;
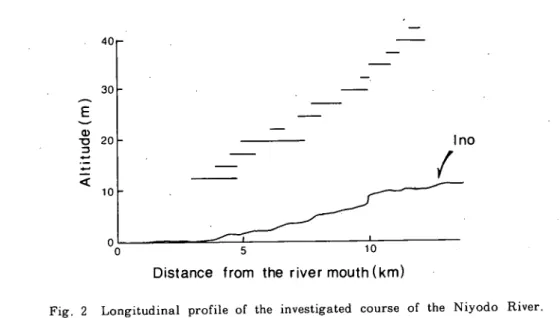
仁淀川下流域の砂疎堆にみられる植物群落 (石川・石田) 267 規模ではあるが,伊野町より上流と比較すると種類,量ともにはるかに豊富である.河川敷で は広く畑作が行われ,これらの畑と砂喋堆の間や,堤防に近い部分には,エノキームクノキ林やモ ウッウチク林が成立している(山中, 1981).調査対象としたのは,伊野町から下流の約10 km の範囲に‘ある砂磯堆である.また河口部付近の砂疎堆は人為的破壊が著しいため,調査対象から除 外した. ’ 調査範囲の河床縦断面図および砂疎堆の位置をFig. 2に示す.河床勾配は河口付近でやや緩 くなるが,調査範囲内では大きな変化は見られず,約1‰の勾配を示す.ただし,砂疎堆により, 瀬や淵が形成されるため,詳細にみると勾配は小さく変動する.図中の不連続な地点は八田堰にあ たる. 40 30 20 10 ︵E)apnmiv 0 0 5 1 0
- Distance from the river mouth (km)
Fig. 2 Longitudinal profile of the investigated course of the Niyodo River 13 lines over the profile show the extent of bars.
方 法
現地調査は1981年から1983年にかけて行ったが,主として1981年の夏から秋にかけて行った.植 生調査の資料をもとに群落組成表を作成し,群落相互の種組成の比較検討をした.また各スタンド間 の類似度を比較するため,32のスタンドについて森下の類似度指数Cλ(p)を求めた(Morisita, 1959).その際,被度の値はそれぞれの被度階級の中央値を求めて使用した.これをBray-Curtis
序列法(Bray & Curtis, 1957)によって二次元の図に展開した.これらのスタンドの立地条
件を比較するため,表層堆積物を採取し粒径分析を行った.表層堆積物は,0∼2 cm, 2∼10 cm, 10∼20 cm, 20∼30 cm の4つの深さの層からそれぞれ300 g以上採取した.深い層では中樫も含 まれていたが,採取対象としたのは小磯以下の・matrixに限った.これらの資料を電熱乾燥機を用 いて約1時間乾燥させためちに,ふるいで分析した.ふるいは−2φ(境界粒径4 mm) ,' ― 1φ (同2 mm), 0φ(同1 mm), 1φ(同0.5mm),2φ(同0.25 mm)の5つを使用し,2φより 細粒な物質の詳細な分析は省略した.一方,調査地域の植生を構成する主要な24種類について,そ れぞれの分布相関を森下のR5(p) (Morisita, 1959)を用いて検討した.
268 高知大学学術研究報告 第34巻(1985)自然科学
結果および考察
各群落の立地と種組成
砂疎堆上に成立する植物群落を植生図に示したものがFig. 3である.図中の砂篠堆のうちA とCはlongitudinalbar, Bはpoint bar と呼ばれるものである(Lewin, 1978).また,群 落の総合常在度表をTable l に示す.これらの資料をもとに優占種と他の構成種によって区別 された群落について,立地と種組成を中心に以下に述べる. 一 B ノ
ノ
ノ C // / ●i 。。 f φ 匹 12 34。5 一回国一″一回皿□6
0 100 mFig. 3 Vegetation maps of three bars. A and C : longitudinal bars,
B: point bar, 1:Kummerouio Stφulaceacomm., 2: Phragmitesiaponica
com°・・ 3: Salix gracilisりla CO°m・,4; Misca几tht↓s sacchariμort↓sCOmm・,
5: Polygonum hvdropipercomm., P.hjidropiper-Polygonum thixabergii
co°m・ぺP・ hjidropiper-S.gracilistyla comm. and P: Kvdropiper-P.japoaica
・comm. 6: Bare land ゛。 1)ヤナギタデ群落 −. . ヤナギタデが優占し,ツルノゲイトウ,ミゾソバ,イヌタデ,イヌ,ビエなどの一年生草本が多く, 出現する.この群落は,砂疎堆の河道に接した部分を縁どるように発達する.河道から離れ,地下 水位が低くなると急激に少なくなる.また,流水の破壊作用を直接受けやすい砂疎堆の上流側には 少ない.前年度の枯死した茎の分布を調査した結果からも,この群落は,毎年変化する河道や,砂 疎堆の発達あるいは移動に伴う地下水位の変動により,その立地を移動させていると考えられる. 2-a)ヤナギタデーミソソバ群落 ヤナギタデとミゾソバが優占する群落で,ヤナギタデ群落より,堆積物の粒径の細かい砂疎堆の
仁淀川下流域の砂陛堆にみられる植物群落 (石川・石田) .269 最下流部などに成立することが多い.他の構成種はヤナギタデ群落とほぼ同1じである. 2−b)同,・ネコヤナギーツルヨシ型 ヤナギタデーミソソバ群落に分類されるスタンドの中には,ヒメクグ,カヤツリグサなどの多年 草,あるいはネコヤナギの実生が多数侵入しているのが認められる.ネコヤナギの実生が生長して いく過程で,そのまわりに砂疎か堆積し,ネコヤナギが低木林を形成するころには,まわりとの比 高が40∼50 cm に達するかなり大きなマウンドに発達す・る.この群落型には,その間の発達段階 の違う群落が含まれている.ネコヤナギが大きくなるに従い,ヤナギタデ,ミゾソバなどは減少し, ツルヨシやヨモギなどの出現頻度が高くなる. 3)ヤナギタデーネコヤナギ群落 上記の2―b)の群落とほぼ同じであるが,ミゾソバが優占しない点で区別される.ネコヤナギ を核とするマウッドの下流側に形成されることが多く,厚くはないが,表層には細粒な物質が堆積 している立地が多い.マウンドが高くなるに従い,ヨモギ,メヒシバ,マルバヤハズソウなどが侵 入してくる. 4)ヤナギタデーツルヨシ群落 ッルヨシは河道縁辺や砂疎堆の中に見られる帯状の凹地(増水時には河道になる)に群落を発達 させることが多い.このうち,河道縁辺の地下水位の高い立地には,ヤナギタデを伴うことが多い. 一方,ヤナギタデ群落の中に漂着したと思われる走出枝や,実生などが混在することも多く,これ らのうちヤナギタデとツルヨシの優占度が高いスタンドをこの群落型にまとめた.ミゾソバ,イヌ タデ,ヤノネグサなどのタデ属の他に,ヒナタイノコズチ,ツルノゲイトウなどの湿潤な立地に出 現頻度の高い種が多い.しかし,優占度は低いが,マルバヤハズソウ,オオアレチノギク,コセン ダングサなどの乾燥した立地に多い種もかなり高い頻度で出現する. 5)オギ群落 オギは河道から離れた場所で,中砂や細砂が厚く堆積した立地に多い.オギが密に生育するため か下層の植被率は低く,オギの純群落といっても差し支えないスタンドが多い.他の群落に高頻度 で出現する,メヒシバ,ヒメムカショモギ,アメリカセンダングサ,エノコログサなどは見られな いが,ヤナギタデ,ミゾソバに加え,マルバヤハズソウ,オオアレチノギクも混生し,当調査地域 に限って言えば,種組成的には,はっきりした性格を示さない. 6)ツルヨシ群落 ッルヨシは,調査地全域に広く出現し,多様な立地に生育する.しかし,最も発達.した群落が見 られたのは,オギ群落に接した河道側で,オギ群落と水平に帯状に広く・成立している.前述のよう に,砂疎堆の中ほどから下流部にかけての河道側には,ネコヤナギ低木林を核とするマウンドが所々 に発達しているが,これらのマウンドを取り囲むようにして成立している群落も多く見られる.ま た,ツルヨシのみが生育する大型のマウンドも一部で見られた.ツルヨシは非常に生長の速い走出 枝を持っており,より条件のよい立地に素早く侵入,定着でき,河川のような不安定な環境下では, きわめて有利な種であると考えられる.そのような走出枝の侵入による,未発達の群落も随所に見 られる・.この群落型には,ツルヨシが優占する発達段階の違うスタンドをすべて含めたので,種組 成的には,はっきりした性格を示さない.他の構成種の中では,ケアリタソウ,ヨモギ,ホウキギ ク,メヒシバなどの出現頻度が高い. 7−a)マルバヤハズソウ群落 マルバヤハズッウが優占する群落で,しばしばヤハズソウが混生する.ヨモギ,メヒシバ,コマ ツヨイグサ,ヒメムカショモギなどの優占度が高い.河道から離れだ平らな場所で,中疎の間隙に 小磯や粗砂など粒径の荒いmatrixが密につまった立地に広く成立する.河道から離れているので
270 高知大学学術研究報告 第34巻(1985)・一自然科学
Community type
Number of Quadrats Average number of species
’2 7 ̄下 9 9 1 0 9 3 4 5 25 16 28 14 14 15 6 7 8 - a b、 i r t r t 6 11 3 1 1 2 13C O ζ J q ″ p o l リ g o n u f f lh y d r o p i p e r R ] 1 y g o n u n t む 力 u n b e r g i i l C 力 y r a n t 力 e s f a u r i e i f り 2 y g o n u m l o n g i s e t u m E C ね i n o c h l o a c r u s - g s l l i A l t e r n a n t 力 e r a s e s s i l i s x y l j j n 9 ∂ b r e v j f o j j a P o l y g o n u m n i p p o n e n s e R o r i p p aI n d i e s A r t 力 r a x o n 力 i s p i d u s C h e n o p o d i u m a m b r o s i o i d e s 乙 A j d w i g i a p r o s t r a t a B c l i p t a p r o s t r a t a S K l i x g r a c l l i s t y l a H i s c a n ぐ 力 l a s s a c c h a r i f l o r u s P ね \ r a g m i と e s j り 珍 n i c a K u n m e r o v i a s t i p u l a c e a E r i g e r o n s u B i a t r e n s i s B i d e n s p i l o s a H i s c a n t h u ss i n e n s i s K v n m e r o v i a s t r i a t a O c n o t h o r a J a c i n i a t a 乙 ^ s p e d e z a p 』 ・ l o s s C u s c u t a p e n と a g o n a A r t e m i s i a p r i n c c p s D i g i t a r i a s a n q u i n a l i s E r i g e r o n c a n a d e n s i a B i d e n s f r o n d o s a S e t a r i a v i r i d l s 人 S と e r s u b u l a t u s M o s l a p u n c t u l e t a R u m c xJ B p o n j c u s D i g i t a r i a t i m o r e n s i s P a n i c u m b i s u l c a t u m L a c t u c a i n d i c a P l a n t a g o a s i a t i c a ' X a n t 力 i u m c a n a d e n s e A s t r a g a l u s s i n i c u s J U S と i a t a p r o c u m b e n s F a t o u a v i l l o s a C y p o r u s m i c r o i r i a C Q 閲 ・ e l i n a c o m m u n i s C r s s s o c e p 力 a j U m c r e p i d i o i d e s H u m u l u s j a f り n i c a s S e d u a b u l b i f e r u m Y o u n g i a d e n t i c u l a t a E l e u s i n e i n d i c a v e r o n i c a p e r s i c a ぷ e t a r i a f a b e r i Y o u n g i a j a μ o n j c a S p o r o b o l u s f c r t i l i a S t e l l a r i ∂ n e g l e c t s C x a l i s c o r n i c u l e t a D i g i t a r i a v i o l a s c e n s R o s a m u l t i f l o r a O c n o t 力 e r a b i e n n i s G j り c i n e s o j a A s t e r a g e r a t o i d e s R o r i p p , ∂ i s l a n d i c a S t e l l a r i a s g u a t i c a R u m e x c r i s p u s s o i l d a g o a l t i a s i m a C e l o s i a a r g e n t e a ・ T r i f o l i u m r a p e n s N a l l o t i i s j a μ j n i c u s 乙 ■ e p i d i u j n v i r g i n i c u m P o l y g o n u i a c u s p i d a t u m 戸 ∂ η j C U md i c h o む α m i f l o r u m Z o y3 i a j & p o n i c a j ? u m e x a c e t o s a P o a a n n u a P h a l a r i s a r u n d i n a c e a O e n a n t h a j a v a n i c a E u p h o r ね i a p s e u d c 》 ・ c h a m a e s y c e D o p a t r i u m j u n c e u m P O と e n t i i l a f r e t / n i ∂ n a 乙 ■ u d w i g i ∂ d e c u r r e n s C e r a s t i u o i g l c 肖 O S l a d i a n む 力 e r a A g r o p り r o n t s u k u s h i e n s G S o n c h u ss s p e r C e j と i ss j n e n s j s V i c t a a n g u s t i f o l i a S i i g i n a j a p o n i c a S a l i x e r i o c a r p a M O Z U 5 i a p o n i c u s β Γ a s a i c a n a p u s L e e r s i a j a μ o n i c a V v m m V a ︱ c a ・ -■ H ・ ・ V t a ■ -' Ⅲ a e I I ・ 一 ・ 一 ⅢW ⅣⅡ.III・・nl.lnl・nlnl.I・・.n.・U..・.11.1 nmlnⅢnna ■I‘・・・・・I 一 s s 1 1 ・ VVH-BB--'-0aaHv・VI・・・・・・‘ ⅢP-ca≪H'-'HH'-*'-cJ ■--II III VIIIⅢnllla B -■ aV・・III・II■ ■ aS'-Boca'-'ca-ci ■o-'HCj -I・I・IIIIIIIIII・・・・I・I ln11I・・・・・・︱・・IV’I皿・1・・.・・ V11UⅡ11111nⅡnlll‘・︲Ⅲnnl1 いIT‘√ W V WⅢnmnUnnnlnUllillllr1!llUI・lill・lli・I・III!111111 I︲II ‘I・II■."■・・・ p ■ ja "-・■ H ■-. . 1 . ・ . ' a a . . U I ∼ ・ n ・ I I Ⅲ : ■ B ・ = * ■ t a -・ WⅢnl■EiHH*-"-'^'-'-'a'-'-' ■'-ΦII・IΦII ■■I一泰一一s lssl一1 41弗ΦII U ・ 1 T s u r u n o g e l t o H l m c k u g u V a t i o n e g u s a I n u g a r a s h l K o b u n a g u s a K e a r l c a s o C h o j i c n d e T s k a s o b u r S N e k o y a n a g i O g i T s u r u y o s h i M a r u b a y a h a z u s o 6 a r e c h l n o g i k u K o s c n d a n g u s a S u s u k l Y a h a z u s o ‘ ニ K o m a t B u y o l g u s a M c d o h s g i A n e r l k a n e n a s h l k a z u r a Y o m o g i H c h l s h l b a H l m c m u k a s h l y o m o s l A m c r i k a s e n d a n g u s o E り o k o r o g u s a ・ H o k l g t k u I n u k o j u C l s h l g l s h l K o m e h l s h l b a N u k a k l b l A k ・ l n o n o g e s h l O b a k o 6 o n a i n 。 m 1 C e n g e K l c s u n e n o m a g o K u u a k u s a K a y a t s u r l g u s a .T s u y u k u s a S c n l b a n a b d r o g l k u K a n a n u g u r a K o m o c h l m a n n e n g u s a Y a k u s h l s o O h l s h l b a 8 i n u り o f u g u r l A k i n o e n o f c o r o g u S B O n l t a b l r a k o N c z u m l n o o ・ H l d o r l h a k o b e K a t a b a m l A k i n c h i s h i b i i N o i b a r a A r e c h l n a t s u y o l g u s a ‘ T s u r u m a m e N o k o n g l k u S u k a s h i t a g o b o U s h l h i k o b c ・ N a g a b a g i s h l g l s h l S e 1 C a k a a w a d a c h i s 6 N o g ' e l c S S h l r o t s u m c k u s a A k a m C g a s h l w a H a r a e g u n b a 1 n a z u n a I c o d o r l Q k u s o k l b l S h i b n S u l b a S u z u i n e n o k a c a b l r a , K u s o y o s h l S e r l N I s h i k l s o A b u n a n c M l t s u b a t s u c h l o u r l H l r c C a g o b o O r a n d a m l m L n a g u s a H l m c i l a o K f i m o J I g u s a O n l n o g c s h i E n o k i K a r a s u n o e n d 6 T s u n c k u s a J o y a n o g i T o k l u n h n z a S e l y o a b u r a n a A s h i k a k l 。
仁淀川下流域の砂疎堆にみられる植物群落 (石川,石田)
Table l Synthetic table for the plant communities developed on bars along the
Niyodo River. h Po1:ygonitm. fiydropipercomm. 2−a: p.
hydropφer- Polygonumびiunbergii comm., 2-b: Ditto:Salix gracilisりla- PKragmites
japonica type, 3: j).hydropφe「-S.gracilistjila comm.,
4: P.hvciro- piper-p. joponica comm., 5: Miscanthus sacchariμoruscomm., 6: j).
japonicacomm・, 7-a: Kummeroひ1a s£φulacea comm。7-b: Ditto. p.
ja- ponica type, 8: S. gracilisりla comm.
271 地下水位は低いが・平らな場所で, matrixが密なだめ,完全な乾燥状態になることはなく,夏期 でも.10 cm より深いところでは湿り気を保っている. 7−b)同,ツルヨシ型 ツルヨシ群落に隣接した所では,マルバヤハズソウ群落の中に,ツルヨシの走出枝が侵入してき て,小さな株を定着させている.緩い傾斜を持ち, matrixが密でない立地が多く, 7-a)の立 地よりは乾燥している.そのため,ヤナギタデ,イヌビエ,ツルノゲイトウなどの湿潤な立地に出 現頻度の高い種は全く見られなかった.コセンダングサ,ヤハズソウ,メヒシバ,ヒメムカショモ ギなどの出現頻度が高い/ 8)ネコヤナギ群落 ネコヤナギが優占する群落で,高さ1mから2mの低木林を形成する.平均出現種数はわず かに3種程度で,ネコヤナギ以外の種は優占度も低く,ネコヤナギの純群落といえる.この低木林 を核として,時には比高1mを越えるマウンドが形成されることがある.砂疎堆の中央部にはな く,河道の縁辺に点々と成立する.この理由としては,地下水位が高いごとが,発芽と実生が育つ 条件として必要であることが考えられる. その他の群落として,アカメヤナギ群落が認められる.アカメヤナギは,砂呻堆の後背側の湿潤 な立地や,放棄された河道に細砂やシルトが堆積した場所に,低木林,時には高木林を形成するが, 得られた資料が少なく,本報ではその詳細については触れない. ‥’ 主な出現種の分布相関 グい 調査地域の植生を構成する主な24種類の分布相関の指数を求め,クラスター分析を行った結果を Fig. 4 に示す. ●j プラスの分布相関を示す種群を分類すると8群が認められるが,これらのうち1∼4はレベルに 大きな差はなく,分析の過程で違う群に分類された種の中にも,プラ・スの相関を示すペアが多い. 7と8にも同様のことが言える.そこで,分布相関の指数のレベルを−0.2に下げると, A, B, C,Dの4群にまとまる.以下この4群について検討する.Aに含まれるツルノゲイトウ,ヒ ナタイノコズチ/ミゾソバ,ヤナギタデ,ヤノネグサ,チョウジタデ,ケアリタソウ,イヌビエ, イヌタデ,イヌガラシ,ヒメクグ,タカサブロウの13種は, Table 1 では河道縁辺の湿潤な立地 に成立する1).∼4)の群落に出現頻度の高い種である.Cに含まれるのはオギのみであり,どの 種とも低い分布相関しか示さず,その分布は他の種群とはかなりかけ離れていると言える.Dには メドハギ,ツルヨシ,アレチマッヨイグサ,オオアレチノギク,ススキ,コセンダングサ,マルバ ヤハズッウ,ヤハズソウの8種類が含まれる.ツルヨシを除くいずれの種も,Aに含まれる種群と 比べ,より乾燥した立地に生育しTable 1 では7)と4)の群落に出現頻度の高い種である.
272 高知大学学術研究報告 第34巻・(1985 )自然科学 AIternonthera sessitis Achyranthes fauriei ArtKraxon hispidus Polygoれぱ7 thunbergii Potygonum hydropiper Polygonum nippo?lense Luduigia prostrata CHenopodium・ambrosioides Eahinochtoa CnλS−galH P011ugcnwn longisetum Rorippa iれ直ca Kytlinqa brevifoZia Eolipta prostrata Sdlix gracilietyla Moslalpunctutatc Miscanthus sacchariflorus Lespedeza pit08a P知卯吋{.te8 jdponiea Oenothera Vaainiata Erigeraれsumatrensis M・む8canihu8 sinenais Bidens pito8a Kurnnerouia8tr・iata Kunmerヽouiaetipulaoea 1 2 3 4 5 6 7 8 A B C D −1 −0.5
0
R5(P)
十〇.5 +1Fig. 4 Dendrogram of 24 species derived from cluster analysis. Morisita's R(5(p) is used as similarity coefficientbet‘weenspecies. 1 to 8 are species clusterswith positive coefficients.Based' on level a (coefficient= - 0.2), these species are grouped in four clusters as A,B, C and D.
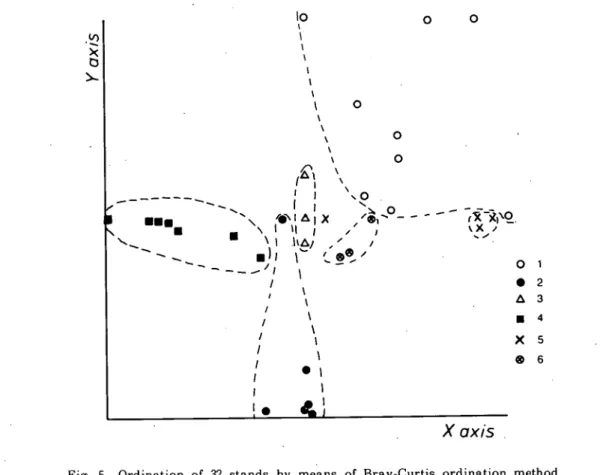
ツルヨシの立地の範囲は広く,ヤナギタデやマルバヤハズソウとも群落を形成する.しかし,分布 相関からみると,Dの種群に近い立地に生育すると考えられる.Bに含まれるイヌコウジュは,調 査地域全体に広く出現する種であるが,ネコヤナギとの分布相関がやや高く,この2種は,種群D よりも種群Aにより高い分布相関を示す・ Fig. 4とTable 1 を比較してみると,1)∼3)の 群落とAの種群,7)の群落とBの種群が対応し,4)゛は両方の種群を含む移行帯にあたる群落と いえる. .’ 群落のオーディネーション 各群落型の中から選定した32のスタンドについてオーディネーションを行った結果を■ Fig. 5 に示す.この図に見られるように,各群落は比較的良いまとまりを示す,ヤナギタデ群落は図の左
仁淀川下流域の砂即堆にみられる植物群落 (石川・石田) 273 側に,ツルヨシ群落は1つのスタンドを除いて,図の下に集中した.マルバヤハズソウ群落は,図 の右上に集ま'るが,比較的分散する傾向を示している.オギ群落は中心にまとまっているが,これ は,他の5つの群落型のいずれとも類似度が低いためである.ネコヤヂギ群落は,マルバヤハズソ ウ群落に近い位置にまとまるが,ヤナギタデを伴った,ヤナギタデーネコヤナギ群落は,ヤナギタ SIXD X "ミミ= ヘペ ■■●■ y、 ヘヘヘ・、 − ・ −ぺゝ ● \ X I’] 一 一 一 一 令 111 勺 / / 旧∼I X I I χ χ ゝ χ χ χ £ ヽ / | △ き I∼II ・ ` ”い’/、 `゛11111 1−IMい八I X ゝ ○ / / / 旦 9 . ○ \4 、 j / / ○ ○ 、O_ ○ ○
'∩が腎
χ【】χis ○ ● △ 扁 X @ 2 3 4 5 6Fig. 5 Ordination of 32 stands by means of Bray-Curtis ordination method.
Morisita's Cバp) is used as similarity index between communities.
l:Kummeroり必s£φulacea comm., 2: PhragiTvitesjapoai£acomm., 3:
Mis- canthussacchariμoruscomm., 4: Polygonumんydropiper comm., 5:Salix
gracilistylacomm., 6: P.hydropφe「-S,gracilistylacomm. r . i
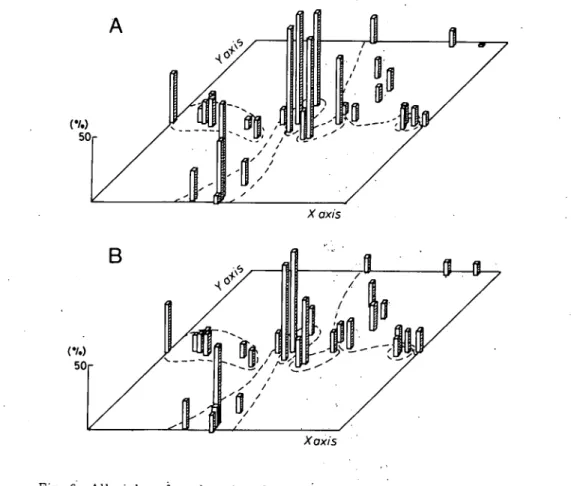
デ群落とネコヤナギ群落の中間に位置し,類似度指数も両群落の中間を示す. 各群落とも比較的良いまとまりをみせたのは,群落の類似度指数に,個体の量(ここでは被度を 使用)を考慮したCλ(p)を使用したため,優占度の高い種に類似度が大きく左右された結果だ と思われる.ここには図示しなかったが,種の存在のみで決定される,野村・シンプソンの指数 (木元, 1976)などを用いた場合,マルバヤハズソウ群落を除いて,各群落型のスタンドはかなり 混在する傾向を示す.この傾向は, Table 1 の種組成を比較しても容易に理解できる. 表層堆積物の粒径組成 各スタンドの表層堆積物を粒径分析した資料からOφ以下,すなわち1mm以下の粒径の堆積物 を重量パー々ントで示しだのがFig. 6である.つまり,バーの高さが高いスタンドほど細かい
274 高知大学学術研究報告 第34巻(1985)自然科学 堆積物が多いことを示す.図中のスタンドの位置はFig. 5に示・したものと同じである.Aは O∼2cmの深さから,Bは2∼10 cm の深さから採取した堆積物について図示したものである. O∼2cmの深さでは,オギ群落,ヤナギタデーネコヤナギ群落で細かい粒径の物質が多く,マル バヤハズソウ群落,ネコヤナギ群落,ヤナギタデ群落で少ない・‥ツルヨシ群落ではスタンドごとの XぴxiS Xaxi゜5
Fig. 6 Alluvial surface deposits of each stand as shown in Fig. 5. Height ofbars indicates percentage in weight of deposits smaller than l mm. A:O-2cm in depth, B: 2-10 cm in depth 」・
差が大きい.2∼10 cm の深さでも同様の傾向があるが,各群落の粒径の差はかなり縮まっている といえる.オギ群落と2∼3のスタンドを除けば,他の5つの群落型における堆積物の粒径の差は ほとんどない.さらに深い,10∼20 cm, 20∼30 cm の層の堆積物では,その差はさらに縮まる. オギ群落では,細粒な物質が厚く堆積している立地が多い.これ,らの立地は,河道から離れた所に あり,流水面との比高も高く,増水時には,浮流物質を多量に含んだ水が,植物体の密度の高いオ ギ群落を通過する際にそれらを沈積させた結果と推定できる(野満・瀬野, 1959).ツルヨシ群落, ネコヤナギーヤナギタデ群落で高い値を示す2∼3のスタンド伝大きな群落で植物体の密度も高 い. 1 本報の調査地域は下流域であるにもかかわらず,河床堆積物には映か多く,・扇状地河川に似た河 状を呈する.大きな砂疎堆が発達するが,植生は貧弱でFig. 3 tc示したように,裸地の占める
仁淀川下流域の砂榛堆にみられる植物群落 (石川・石田) 275 割合が非常に高い.これは,台風による洪水や,梅雨時の増水などの頻度が高く,流水や河床堆積 物の移動による植物群落の破壊が頻繁に行われているためであろう. 1982年の洪水で,前年に調査 した植物群落の半分近くが,砂喋に埋められたり,根こそぎ流されたりして消失した.非常に短い 時間的サイクルの中で,植生が変動しているわけであるが,このように撹乱が卓越した環境下では, ヤナギタデ,ミゾソバ,マルバヤハズソウなどの一年生の草本群落や,ネコヤナギ,ツルヨシなど 物理的破壊に比較的耐性のある植物が群落を維持できるのであろう(Grime, 1979).中でもネコヤ ナギの生長に伴って,それを核にしながら形成されるマウンドの周囲にツルヨシが群落を発達させ たり,マウyドの下流側に細粒な物質が堆積することによって,ヤナギタデーネコヤナギ群落等に 見られるように他の植物が生育できる立地が作り出されているという事実は,河川という環境の中 での植物の立地を考える上で興味深い.今後,これらの群落が相互にどのような関連を持ちつつ, どのような変動をしながら生活をしているのかを明らかにするためには,河川の諸々の環境作用を 考えながら,短いインターバルで詳細な調査を行う必要がある. 摘 要 1.高知県の中央部を流れる仁淀川の下流域において,砂疎堆上に成立する植物群落の調査を 行った. 2. 全体に裸地の占める面積が広く,植生は貧弱であるが,優占種および種組成によって,以 下の8群落型と2亜型が認められた.1)ヤナギタデ群落. 2-a)ヤナギタデーミソソバ群落, 2−b)同,ネコヤナギーツルヨシ型,3)ヤナギタデーネコヤナギ群落,4)ヤナギタデーツル .ヨシ群落,5)オギ群落,6)ツルヨシ群落, 7-a)マルバヤハズソウ群落, 7-b)同,ツル ヨシ型,8)ネコヤナギ群落 3.1)−4)は河道縁辺の地下水位の高い湿潤な立地に,5)は河道から離れた細粒物質の 堆積が厚い立地に,7)は河道から離れた地下水位の低いやや乾燥した立地に成立する.6)の立 地は幅広いが,河道近くに多い.8)は河道近くに多く,砂疎か堆積してマウンドを形成する. 4.堆積物の粒径分析の結果,表層O∼2cmでは差が見られたが,それ以深の層では,5) と6)の一部のスタンドを除いて,大きな差は認められなかった. 5.主要な出現種の分布相関を検討した結果,1)∼3)の群落に出現する頻度の高い種群と 7)の群落に出現する頻度の高い種群が明瞭に区別できた. 引 用 文 献
Bray, J. R. and Curtis, J. T. 1957. An ordination of the upland forest communities of southern Wisconsin. Ecol. Monogr・, 27: 325-349.
Grime J. p. 1979. Plant strategies & vegetation processes. 222 pp. John Wiley & Sons. 波田善夫. 1972.瀬野川の河床植生.広島大学生物学会誌,39: 18-21.
木元新作. 1976.動物群集研究法I一多様性と種類組成-192 pp- 共立出版,東京. 小出博. 1970.日本の河川. 248 pp- 東京大学出版会,東京.
Lewin, J. 1978. Floodplain geomorphology. Progr. Phys. Geogr・,2: 408-437.
Morisita, M. 1959. Measuring of interspecific association and similarity between communities. Mem. Fac. Sci., Kyushu Univ. Ser. E (Biol.), 3: 65-80.
中西弘樹・関太郎. 1979.小瀬川の河辺植生.弥栄峡の自然総合学術調査研究報告書. 397-422,広島・ 山口.
276 高知大学学術研究報告 第34巻 n985 自然科学 野満隆治・瀬野錦蔵. 1959.新河川学. 348 pp- 地人書館,東京. 大井次三郎. 1975.日本植物誌. 1560 pp. 至文堂,東京. 長田武正. 1979.原色帰化植物図鑑. 425 pp- 保育社,大阪. 山中二男. 1981.南四国における暖温帯の河辺林. Hikobia Suppl., 1: 257-264. (昭和60年9月30日受理) (昭和61年3月29日発行)