DOI: http://dx.doi.org/10.14947/psychono.33.3
視覚運動プライミングにおける空間的距離の効果
吉 本 早 苗
a, b・竹 内 龍 人
a*
a日本女子大学・b日本学術振興会
Effect of spatial distance on visual motion priming
Sanae Yoshimoto
a, band Tatsuto Takeuchi*
aa Japan Women s University, b JSPS Research Fellow
It is known that the perceived direction of a directionally ambiguous test stimulus is influenced by the moving direction of a preceding priming stimulus. To examine the spatial property of motion priming, we manipulated the spatial distance between the priming and test stimuli. Subjects judged the perceived direction of 180-deg phaseshift-ed sine-wave gratings (test stimulus) displayphaseshift-ed immphaseshift-ediately after the offset of a priming stimulus. We found that a brief priming stimulus induced negative motion priming when it was spatially distant from the test stimulus. We also found that positive motion priming was observed when both priming stimulus and test stimulus were presented at the peripheral retina. Negative motion priming was observed regardless of the position of the stimuli when the duration of the priming stimulus was longer. We conclude that the effect of spatial distance on the visual motion priming could be explained by the activation of a center-surround antagonistic motion detecting mechanism when a brief priming stimulus was used.
Keywords: motion perception, visual motion priming, center-surround antagonism, retinal eccentricity
は じ め に 視覚パターンに対する我々の知覚は,時空間的に異な る他の視覚パターンの影響を受ける。視覚系が絶えず入 力される視覚パターンをどのように統合しているかを理 解することは,人間の視覚情報処理の解明に向けた重要 な問題の一つである。運動知覚は視覚情報の時空間的な 変化によりもたらされるが,局所的な運動情報のみに依 存するのではなく,時空間的近傍に存在する運動の影響 を強く受ける(西田・竹内・蘆田,2000)。そういった 運動知覚の時間的特性の一つとして,ある特定の運動方 向をもたない多義運動刺激が提示されたとき,知覚され る運動方向はそれに先行する運動刺激の方向に依存する という現象が挙げられる。先行する運動刺激の持続時間 が短い場合には,後続する多義刺激が先行刺激と同じ方 向に知覚される視覚運動プライミング(visual motion priming)が生じる(Anstis & Ramachandran, 1987; Campana,
Pavan, & Casco, 2008; Jiang, Pantle, & Mark, 1998; Jiang, Luo, & Parasuraman, 2002; Kanai & Verstraten, 2005; Pantle, Gallogly, & Piehler, 2000; Pavan, Campana, Guerresch, Manassi, & Casco, 2009; Piehler & Pantle, 2001; Pinkus & Pantle, 1997; Raymond, O’Donnell, & Tipper, 1998; Takeuchi, Tuladhar, & Yoshimoto, 2011)。なお後述するように,視覚運動プラ イミングには,多義運動刺激が先行刺激とは反対方向に 見える場合もあるため,本稿では多義運動刺激が先行刺 激と同方向に運動して見えるプライミング現象を「正の 視覚運動プライミング」,多義運動刺激が逆方向に運動 して見える現象を「負の視覚運動プライミング」と呼ぶ。
Pinkus & Pantle(1997)は,位相が180 deg反転する正 弦波縞から成る多義運動刺激をテスト刺激として用い た。先行刺激(滑らかに運動する正弦波縞)の持続時間 が300 msよりも短い場合に,テスト刺激が先行刺激と 同方向に知覚される正の視覚運動プライミングが生じる ことを報告した。一方で,類似の刺激布置を用いたPantle et al.(2000)は,先行刺激がより長い時間,例えば0.5 s 以上提示される場合は,運動残効と同様に,テスト刺激 が先行刺激と逆方向に知覚されることを確認した。この ような負のプライミング現象は,テスト刺激として時間 Copyright 2014. The Japanese Psychonomic Society. All rights reserved. * Corresponding author. Department of Psychology, Japan
Women’s University, 1–1–1 Nishi-ikuta, Tama-ku, Kawasaki-shi, Kanagawa 214–8565, Japan. E-mail: [email protected]
的に変動する多義運動刺激が使用されているという点 で,いわゆるフリッカー運動残効(Nishida & Sato, 1995) と密接に関係した現象であると考えられる。先行刺激の 持続時間により正負のプライミングが切り替わること は,その後の研究でも確認されている(Kanai & Verstraten, 2005; Takeuchi et al., 2011)。
運動する正弦波縞を用いたこれまでの視覚運動プライ ミングに関する研究(e.g., Pinkus & Pantle, 1997)では, 先行刺激とテスト刺激(多義運動刺激)は常に同じ空間 位置に提示されていた。そこで本研究では,プライミン グ効果における先行刺激とテスト刺激との空間的な位置 関係の影響について検討することを目的とした。実験で は,先行刺激とテスト刺激との空間距離(2刺激の中心 間距離)を離し,プライミングの生起頻度を推定した。 先行刺激の持続時間が長い場合に観察される負のプラ イミングがフリッカー運動残効と類似の現象であれば, 先行刺激とテスト刺激の空間距離が離れても,負のプラ イミングが生起すると予測される。順応刺激とテスト刺 激の空間位置が異なる場合でも空間転移が生じ,結果と してフリッカー運動残効が観察されるからである(von Grünau & Dubé, 1992; Snowden & Milne, 1997; McGraw & Roach, 2008)。
では先行刺激とテスト刺激の空間距離が離れている場 合,持続時間が短い先行刺激はどのような効果をもたら すであろうか? 二つの可能性が考えられる。静止した パターンの場合と同様,運動刺激に対する感度も周辺に いくほど低下する(McKee & Nakayama, 1984)。テスト 刺激を中心視野に提示した場合,先行刺激の周辺視野提 示により距離が離れるにつれ,正のプライミングの知覚 確率は単調に減少するという予測が成り立つ。Pinkus & Pantle(1997)は,運動検出メカニズムにおける低域通 過型のフィルタ特性により先行刺激とテスト刺激の情報 が統合され,正のプライミングが観察されるというモデ ルを提案した。このモデルに則れば,視覚運動プライミ ングが消失する空間距離が,Pinkus & Pantle(1997)が 仮定するタイプの運動検出器における空間的な大きさの 推定値となりうると考えられる。
一方で,これまでの神経生理学的研究から,運動検出 メカニズムは中心部と周辺部が互いに抑制し合う中心周 辺拮抗型の空間構造を持つことが知られており(Allman, Miezin, & McGuinness, 1985; Born & Tootell, 1992; Eifuku & Wurtz, 1998),対応する知覚現象も報告されている(Bak-er & Graf, 2010; Golomb, And1998),対応する知覚現象も報告されている(Bak-ersen, Nakayama, MacLeod, & Wong, 1985; Ido, Ohtani, & Ejima, 1997; Murakami & Shimojo, 1993, 1996; Sachtler & Zaidi, 1995; Shioiri, Ito, Sakurai, &
Yaguchi, 2002; Tadin, Lappin, Gilroy, & Blake, 2003)。例え ば,運動パターンのサイズ拡大に伴い運動方向弁別感度 は低下するという一見直感に反する現象が報告されてい るが(Tadin et al., 2003),中心周辺拮抗型のメカニズム における周辺抑制を仮定すれば説明できる。中心周辺拮 抗型の相対運動検出メカニズムでは,中心と周辺に同じ 方向の運動刺激が提示された時に,中心視野での運動検 出が抑制される。これを視覚運動プライミングの状況で 考えた場合は,こうしたメカニズムが,先行刺激とテス ト刺激を統合するためにある程度の時間幅をもって働く と仮定する必要がある。この仮定の下では,周辺に提示 された先行刺激と同方向の運動信号へ抑制がかかるた め,中心視野に提示されたテスト刺激には周辺と反対の 運動方向へのバイアスがかかることが予測される。つま り,先行刺激とテスト刺激との間の距離が増加すると, 当初知覚されていた正のプライミングが負へのプライミ ングへと切り変わる可能性がある。これが二つ目の予測 である。 実 験 1 目 的 実験1では,視覚運動プライミングの知覚が上述した 予測に合致するかどうかを調べることを目的とした。先 行刺激を周辺視野に提示し,テスト刺激(多義運動刺 激)を中心視野に提示することで両者の空間位置を異に した。被験者は左右いずれかに運動する先行刺激を観察 した後,中心視野に提示されたテスト刺激の見かけの運 動方向を回答した。 方 法
装置 プログラミング言語(MATLAB ver 7.8, Math-Works Inc.)と視覚実験用の関数群Psychtoolbox 3(Brain-ard, 1997; Pelli, 1997)により作成した視覚刺激をパーソ ナルコンピュータ(MacPro, Apple Inc.)で制御し,CRT (GDM-F520, SONY)に提示した。CRTの時間解像度は 120 Hz, 空間解像度は1024×768ピクセルとした。CRTの ガ ン マ補 正 は 色 彩 輝 度 計(ColorCALII, Cambridge Re-search Systems Ltd.)により行った。視距離は 57 cm で, 画 面 の 平 均 輝 度 は 46.8 cd/m2と し た(CIE1931 x=0.31, y=0.32)。実験は暗幕で覆った室内で行った。被験者の
視線位置の測定には眼球運動計測器(Monocular GigE-60 Eye-Frame Scene Camera System MSE07, Arrington Research Inc.)および眼球運動計測用ソフトウェア(ViewPoint ver 2.8.4, Arrington Research Inc.)を使用した。眼球運動 計測器の時間解像度は 60 Hz, 空間解像度は0.15 degで
あった。視線位置の同定はオフラインで行った。 被験者 4名の被験者が参加した。うち1名は著者で あり,その他3名は実験の目的を知らなかった。 視覚刺激 先行刺激(Primer)としては,水平方向に 運動する垂直正弦波格子(空間周波数0.5 c/deg)を用い た(Figure 1)。 刺 激サ イ ズ は 22.4×3.3 deg と し, 縦の エッジをぼかすためにガウス窓をかけた(σs=3.0 deg)。 先行刺激の輝度コントラスト(マイケルソンコントラス ト)は運動方向弁別コントラスト閾の10倍という比較 的高い値に設定した。その理由は,コントラストが高い ほど中心周辺拮抗型の特性があらわれることを示した先 行研究の結果(Tadin et al., 2003)に基づく。先行刺激の 速度は7.5 deg/sとした(1フレームの持続時間は33.3 ms, フレーム間の移動距離はπ/4 rad)。この速度値は,予備 観察および先行研究(Takeuchi et al., 2011)から視覚運 動プライミングが安定して観察される値として選択し た。 先 行 刺 激 全 体 の 持 続 時 間 は 133.3 ms, あ る い は 766.7 msとした。先行刺激とテスト刺激を同じ位置に提 示した過去の研究(Kanai & Verstraten, 2005; Pantle et al., 2000; Takeuchi et al., 2011)から,前者では正の視覚運動 プライミング,後者では負の視覚運動プライミングが生 じることが予測される。 先行刺激の提示位置は中心から周辺上方 0.0–12.2 deg (全8通り)とした。なお,ここでいう中心とは凝視点 の提示位置(およびテスト刺激の中心)のことであり, CRT画面上の中心ではない。先行刺激を画面上部に提示 する必要から,画面中心から3.3 deg下方を凝視点を提 示する「中心」とした。また先行刺激とテスト刺激間の 距離は,先行刺激とテスト刺激の中心間の距離として定 義した。そこで以降ではこの距離のことを「中心間距 離」と記述する。先行刺激とテスト刺激の中心間距離お よび先行刺激の持続時間は試行ごとにランダムとした。 先行刺激が周辺に提示される場合,中心には白い十字の 凝視点(1.0×1.0 deg)を提示した。 テスト刺激(Figure 1内のtest)としては,見かけの運 動方向が曖昧となる多義運動刺激を用いた。フレームご とに位相が 180 deg変化する垂直正弦波格子を4フレー
ム提示した(Kanai & Verstraten, 2005; Pantle et al., 2000)。 テスト刺激の持続時間は533.3 msとした。先行刺激と同 じ見かけの速度(7.5 deg/s)とするために,テスト刺激 の1フレームあたりの持続時間(133.3 ms)は先行刺激 の4倍とした。テスト刺激の輝度コントラストは0.5に 固定した。テスト刺激は常に中心に提示した。テスト 刺激が提示されている間は凝視点は消去した。視覚運 動プライミングの効果が弱まらないようにするために, 先行刺激とテスト刺激との時間間隔(ISI)は0 sとした (Kanai & Verstraten, 2005)。
コントラスト感度の測定 本実験では,先行刺激をい ろいろな空間位置に提示している。提示位置により検出 のしやすさが変わらないようにするために,先行刺激の コントラストを各提示位置における運動方向弁別コント ラスト閾の10倍とした。閾値の測定は先行刺激と全く 同じ刺激を用い,本実験に先んじて行った。1000 Hzの 純音の後に,刺激は8カ所の提示位置(0.0–12.2 deg)の どこかにランダムで提示された。被験者は強制2肢選択 法により,刺激の運動方向が左右どちらであったかを回 答した。正誤のフィードバックは行わなかった。刺激の コントラストを階段法に則って変化させ,正答率 79% となるコントラスト値を求めた(Levitt, 1971)。正答が3 試行連続したときにコントラストは減少し,誤反応した と き に増 加 し た。 測 定 開 始 後 は コ ン ト ラ ス ト を 0.4 log unitで増減させ,増減の反転を2回繰り返した後 は0.1 log unitで増減した。増減の反転が6回となった時 点で測定は終了した。全6回の反転のうち,最後の3回 分のコントラストの平均を求め,その値をコントラスト 閾とした。各被験者とも,運動方向弁別のコントラスト 閾は刺激が中心から離れるにつれて上昇した。そのため 以下に記述する本実験においては,中心に近い位置に提 示される先行刺激のコントラストは,中心から離れる位 Figure 1. Schematic description of the direction
judg-ment task in Experijudg-ment 1. On each trial, a priming stimulus (primer) moved left or right for 133.3 or 766.7 ms (It was right in the figure). Subsequently a directionally ambiguous test stimulus was presented, and subjects judged the perceived direction of the test stimulus. The distance between the primer and the test stimulus was varied from 0.0 to 12.2 deg.
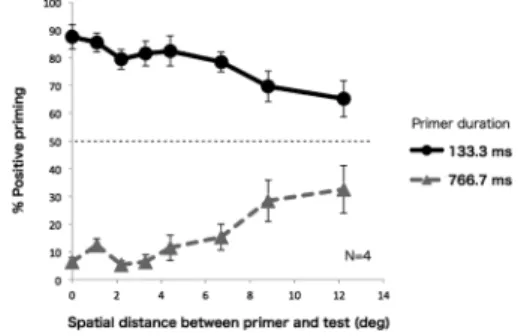
置に提示される刺激のコントラストよりも低くなった。 運動方向判断課題 各被験者につきコントラスト閾を 測定した後に,テスト刺激に対する先行刺激の効果を測 定した。Figure 1に一試行の流れを示す。被験者がキー ボードの5キーを押すと実験が開始された。試行開始時 には1000 Hzの純音が鳴り,凝視点が1 s提示され,その 後先行刺激が中心から 0.0–12.2 degにおけるいずれかの 位置(8通り)に提示された。先行刺激の提示位置およ び持続時間(133.3 msか766.7 ms),そして運動方向(左 か右)はランダムとした。先行刺激提示後すぐにテスト 刺激が中心に提示され,被験者はテスト刺激の見かけの 運動方向をキー押し(左方向判断の場合は4キー,右方 向判断の場合は6キー)により回答した。回答後,1 sの インターバルを挟み次の試行へ移った。1セッションの 試行数は,先行刺激とテスト刺激の中心間距離(8通り)× 先行刺激の持続時間(2 通り)×先行刺激の運動方向 (2通り)×繰り返し(4回)の計128試行であった。なお 20試行ごとに休憩を挟んだ。各被験者とも計4セッショ ンを行った。各被験者とも練習として約20試行を事前 に行った。 視線位置の推定 先行刺激が周辺視野に提示される 時,被験者の注視点が中心に提示された凝視点から外れ ていないかを確認するため,実験中,被験者の視線位置 を眼球運動計測器により測定した。測定は全て右目で 行った。瞬きを除き,視線位置が凝視点から 1.5 deg以 上(テスト刺激の縦方向の長さの0.46倍)外れた試行に ついてはそのデータを除外した。同様の操作を後述する 実験 2,実験 3 でも行った。実験 1 において除外した データは4.9%であった。 結 果 実験1の結果をFigure 2に示す。縦軸はテスト刺激が 先行刺激と同方向に動いて知覚された割合を示す。した がって50%を上回る場合には正のプライミングの報告 数が負のプライミングよりも多かった一方で,50%を下 回る場合は負のプライミングの報告数の方が多かったこ とを示す。二要因分散分析の結果,先行刺激の持続時間 の主効果(F(1, 3)=201.3, p<.001),先行刺激とテスト 刺激の中心間距離の主効果(F(7, 21)=8.2, p<.0001)が それぞれ有意であり,持続時間と中心間距離の交互作用 も有意となった(F(7, 21)=16.2, p<.0001)。 先行刺激とテスト刺激の中心間距離が0 degの時,す なわち両刺激の空間位置が一致する時は,先行刺激の持 続時間が短い場合には正の視覚運動プライミングが生 じ,長い場合には負のプライミングが生じた。これは先
行研究と一貫した結果である(Kanai & Verstraten, 2005; Takeuchi et al., 2011)。先行刺激とテスト刺激の中心間距 離が離れると,先行刺激の持続時間が長い時には常に負 のプライミングが観察された。一方で,先行刺激の持続 時間が短いと,先行刺激とテスト刺激の中心間距離が増 加するにつれて,正のプライミングが負のプライミング へと移行していった。 先行刺激とテスト刺激との中心間距離をさらに離す と,先行刺激の効果が弱まり,先行刺激の持続時間にか かわらずプライミングの生起率が 50%へ収束していく 傾向が見られた。 考 察 先行刺激の持続時間が長い時には,先行刺激とテスト 刺激の中心間距離にかかわらず,負のプライミングが優 位であった。これまでの研究から,運動方向が曖昧なテ スト刺激を用いた時に観察されるフリッカー運動残効 は,先行刺激とテスト刺激の空間位置が異なる場合でも 生じること,つまり空間転移効果があることが報告され ている(e.g., von Grünau & Dubé, 1992)。負のプライミン グは現象的にはフリッカー運動残効と類似していること から(Kanai & Verstraten, 2005),負のプライミングに関 する本実験結果は,先行刺激の提示時間が従来のフリッ カー運動残効研究において検討された順応時間(例えば Figure 2. Result of Experiment 1. The graph plots
per-cent response of positive priming (perceived direction of test stimulus in the same direction as the primer) as a function of the distance between priming and test stimuli in degrees. When higher than 50% of the re-sponses are scored as positive priming, subjects report-ed the motion of the test stimulus to be in the same direction as that of the primer in a majority of trials. Each curve represents the data taken at different prim-ing stimulus duration (133.3 or 766.7 ms). Error bars represent ±1SEM.
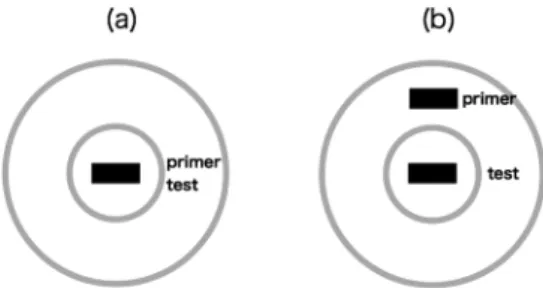
Nishida & Sato(1995)では30秒)より短くても,フリッ カー運動残効でみられる空間転移が生じることを意味し ている。 先行刺激の持続時間が短い時には,もし先行刺激の周 辺視野提示によりその効果が単調減少するのであれば, プライミングの生起率は 50%に収束することが予測さ れる。一方で,中心周辺拮抗型の相対運動検出メカニズ ムがある時間幅をもって機能するのであれば,先行刺激 がテスト刺激から離れるにつれて,プライミングの正負 反転が予測される。実験1の結果は,後者の予測と合致 するものであった。 Figure 3(a)と3(b)には,実験1の結果から推定した 中心周辺拮抗型のメカニズムと視覚刺激との関係を模式 的に示している。灰色で示した同心円は中心周辺拮抗型 の模式的な受容野,黒い長方形は実験で使用した視覚刺 激(Figure 1)を表している。Figure 3(b)に示したよう に先行刺激とテスト刺激の中心間距離を長くした場合, 周辺視野からの抑制作用により,中心視野に提示された テスト刺激における方向知覚が先行刺激とは逆になった (つまり負のプライミングが生じた)のかもしれない。 その場合,中心間距離をさらに延ばせば,先行刺激の効 果はやがてなくなるであろう。このようなメカニズムが 存在するとすれば,中心領域の大きさは正のプライミン グが生成される範囲である約2度である一方,周辺領域 はそこから約10度までの範囲を保ち,負のプライミン グ生成に寄与する構造を持っていると指摘できる。 その一方で,先行刺激の持続時間が長い場合に負のプ ライミング(あるいはフリッカー運動残効)をもたらす メカニズムは,Figure 3に示したような空間構造をもた ないことが実験1の結果(Figure 2)から示唆される。先 行刺激とテスト刺激の空間距離が12.2 degの時にプライ ミングの効果がみられることも,先行刺激の持続時間が 短い時とは異なるメカニズムが関与していることを示唆 している。 実 験 2 目 的 これまでの神経生理学的研究から,視覚野における受 容野のサイズは網膜偏心度と共に増加していくことがよ く知られている(e.g., De Valois & De Valois, 1988)。網膜 偏心度に伴う受容野サイズの増加は,中心周辺拮抗型の 構造を持つ運動方向選択的な神経細胞群においても報告 されている(e.g., Livingstone, Pack, & Born, 2001; Mikami, Newsome, & Wurtz, 1986; Orban, Kennedy, & Bullier, 1986)。 実験1では,テスト刺激を中心視で観察し先行刺激を 中心視野および周辺視野に提示した。実験2では,テス ト刺激を周辺視野に提示し,先行刺激はこのテスト刺激 からさらに周辺に提示した。もし,Figure 3で示したよ うな構造を持つメカニズムが正のプライミングの生成に 関与しているのであれば,先行刺激の持続時間が短い場 合は正のプライミングが生起する可能性が高まると予測 される。というのも,周辺視野では受容野のサイズが増 加することにより,先行刺激とテスト刺激の中心間距離 が開いても,中心周辺の拮抗による抑制がかかりにくく なるはずだからである。 一方でフリッカー運動残効は周辺視においても保たれ ることから(von Grünau & Dubé, 1992),先行刺激の持続 時間が長い場合は,負のプライミングが安定して知覚さ れることが予測される。実験2ではこれらの点を検討す ることを目的とした。 方 法 装置,被験者,実験デザイン,視覚刺激の大きさや時 空間周波数,テスト刺激や先行刺激の持続時間は実験1 と同じとした。実験 1との違いは,刺激の提示位置で あった。テスト刺激は凝視点の上方6 deg(凝視点とテ スト刺激の中心間距離)に提示された。先行刺激とテス ト刺激間の中心間距離は実験1と同様0 degから12.2 deg の8段階とした。そのため,凝視点と先行刺激間の距離 は6 degから18.2 degとなる。 また,実験1に記載した方法で運動方向弁別閾を各網 膜偏心度において測定し,先行刺激の輝度コントラスト を運動方向弁別閾の10倍に設定した。テスト刺激のコ ントラストは実験 1と同様0.5に固定した。被験者は, Figure 3. Schematic description of the stimulus
config-uration and the hypothesized center-surround antago-nistic receptive field. (a) Gray circles represent the hy-pothesized center-surround antagonistic receptive field of the motion detecting mechanism. Both priming stimulus and test stimulus were presented at the cen-tral retina in Experiment 1. (b) The priming stimulus was presented at the peripheral retina while the test stimulus was at the central retina in Experiment 1.
先行刺激とテスト刺激が提示されている間は凝視点から 目を離さないように教示された。被験者が実際に凝視点 を注視しているかを眼球運動計測器で確認し,凝視が外 れた試行はその後の解析から除外した。実験2において 除外したデータは5.4%であった。 結果と考察 実験2の結果をFigure 4に示す。縦軸はテスト刺激が 先行刺激と同方向に動いて知覚された割合を示す。した がって50%を上回る場合には正のプライミングが生じ, 下回る場合は負のプライミングが生じたことを示す。 二要因分散分析の結果,先行刺激の持続時間の主効果 は有意であったが(F(1, 3)=878.0, p<.0001),先行刺激 とテスト刺激の中心間距離の主効果は有意ではなかった (F(7, 21)=0.85, p=.56, n.s.)。持続時間と中心間距離の 交互作用は有意であった(F(7, 21)=13.9, p<.0001)。 以上の結果から,先行刺激とテスト刺激の両者を周辺 視野で観察した場合,先行刺激の提示時間が短い時には 正のプライミングが常に優位になることがわかった。実 験2で検討した空間距離の範囲内では正のプライミング が観察され,実験1の結果とは異なるものとなった。例 えば先行刺激とテスト刺激間が 6.7 deg離れている場合 には,テスト刺激を中心視野で観察した場合は負のプラ イミングが生じる確率が高いが(Figure 2),テスト刺激 (および先行刺激)を周辺視野で観察した場合は正のプ ライミングが知覚されるということである(Figure 4)。 これは,先行刺激とテスト刺激間の絶対距離だけではな く,それぞれが提示される網膜位置がプライミングの生 起確率を決定することを示すと共に,周辺視では視覚運 動情報処理に関連した受容野の空間的なサイズが拡大す るという知見と合致する。 先行刺激の持続時間が長い場合は実験1と同様,負の プライミングが常に優位であった。この結果は,フリッ カー運動残効は先行刺激(順応刺激)とテスト刺激との 間の距離や,提示される網膜位置にかかわらずロバスト な現 象 で あ る と い う 知 見 と 一 貫 し て い る(e.g., von Grünau & Dubé, 1992)。
実 験 3 目 的 実験1ではテスト刺激を常に中心視野で観察した。そ の結果,先行刺激の持続時間が短い場合,先行刺激がテ スト刺激から離れていると負のプライミングが観察され た。実験3では先行刺激とテスト刺激の位置を入れ替え た。もし中心と周辺間で互いに抑制し合っている中心周 辺拮抗型のメカニズム(Figure 3)がプライミングの知 覚に関与しているのであれば,どちらが中心視野に提示 されたかといったことに関わりなく,先行刺激とテスト 刺激の中心間距離が短い時には正のプライミングが,中 心間距離が長い時には負のプライミングが観察されると 予測される。実験3ではこの点を検討した。 方 法 装置,被験者,視覚刺激の大きさや時空間周波数,テ スト刺激や先行刺激の持続時間は実験1と同じとした。 その一方で実験1と異なり,先行刺激を中心視野に提示 した。テスト刺激の提示位置は,中心から周辺上方0.0– 12.2 deg(全8通り)とした。テスト刺激が0 degの場合 は先行刺激とテスト刺激が同じ中心視野に提示されるこ とになり,実験1と同条件となる。先行刺激のコントラ ストを0.5に固定する一方で,テスト刺激のコントラス トをあらかじめ測定した閾値(実験1参照)の10倍とし た。1セッションの試行数は,先行刺激とテスト刺激の 中心間距離(8通り)×先行刺激の持続時間(2通り)× 先行刺激の運動方向(2通り)×繰り返し(4回)の計 128試行であった。各被験者とも計4セッションを行っ た。被験者は,先行刺激とテスト刺激が提示されている 間は凝視点から目を離さないように教示された。実験1 で記した方法により,被験者が実際に凝視点を注視して いるかを確認し,凝視が外れた試行はその後の解析から 除外した。実験 3 において除外したデータは5.3%で あった。 結果と考察 実験3の結果をFigure 5に示す。縦軸はテスト刺激が Figure 4. Result of Experiment 2. The graph plots
per-cent response of positive priming as a function of the distance between priming and test stimuli in degrees. Each curve represents the data taken at different prim-ing stimulus duration (133.3 or 766.7 ms). Error bars represent ±1 SEM.
先行刺激と同方向に動いて知覚された割合を示す。先行 刺激の持続時間が短い時に先行刺激とテスト刺激の中心 間距離に応じて正負のプライミングが切り替わるという 結果は,実験1(Figure 2)と同じ傾向を示した。先行刺 激の持続時間が長い場合は負のプライミングが優位であ るという結果も,実験1と同じであった。二要因分散分 析の結果,先行刺激の持続時間の主効果(F(1, 3)= 216.4, p<.01),先行刺激とテスト刺激の中心間距離の主 効果(F(7, 21)=5.0, p<.01)がそれぞれ有意であり,持 続時間と中心間距離の交互作用も有意となった(F (7, 21)=14.8, p<.0001)。以上の結果は,中心と周辺で 互いに抑制し合う中心周辺拮抗型メカニズムが正のプラ イミングの知覚に関与しているという仮説を支持してい る。 先行刺激の持続時間が長い場合にも,実験1とほぼ同 様の結果が得られている。負のプライミングにおける空 間転移が,先行刺激とテスト刺激のどちらが中心視野で 観察されるかといった位置関係には依存しないロバスト な現象であることが改めて示された。 総合的考察 正 弦 波 格 子 を 用 い た 視 覚 運 動 プ ラ イ ミ ン グ(e.g., Pinkus & Pantle, 1997)に関する研究では,先行刺激(プ ライミング刺激)とテスト刺激は常に同じ位置に提示さ れていた。そこで本研究では,視覚運動プライミングが 先行刺激とテスト刺激の中心間距離の変化によりどのよ うに影響されるかを検討した。視覚運動プライミングに 関するこれまでの研究から,先行刺激の持続時間が長い 時にはフリッカー運動残効(Nishida & Sato, 1995)と類 似した現象(負のプライミング)が観察されるが,先行
刺激が短い時にはAnstis & Ramachandran(1987)が視覚 的慣性(visual inertia)と呼んだ正のプライミングが観 察されると考えられてきた。 しかし本研究における実験から,視覚運動プライミン グ現象は従来考えられていたよりも複雑な現象であるこ とが示された。先行刺激の持続時間が短い場合には常に 正のプライミング(あるいは視覚的慣性)が知覚される わけではなく,先行刺激とテスト刺激の位置関係により 正負の符号,つまりテスト刺激における見かけの運動方 向が反転するのである。先行刺激が周辺視野に提示され た場合には,その持続時間にかかわらず,中心視野に提 示されたテスト刺激が先行刺激の運動方向と反対に動い て見えるという負の視覚運動プライミングが生じる (Figure 2)。この現象は,中心視野に先行刺激を提示し テスト刺激を周辺視野に提示した場合でも観察されたこ とから,どちらかが中心視野に提示された場合は先行刺 激とテスト刺激の空間距離そのものが重要であるといえ る(Figure 5)。 これまでの研究から,運動残効,運動検出の感度,知 覚される速度,曖昧運動の方向知覚,両眼間視野闘争と いった現象は,中心周辺拮抗型の相対運動メカニズムを 仮定すると説明できることが示されている(e.g., Baker & Graf, 2010; Paffen et al., 2005; Sachtler & Zaidi, 1995; Tadin et al., 2003, Takemura & Murakami, 2010)。本研究の結果も 同様に,中心周辺拮抗型の運動検出メカニズムの出力が 視覚運動プライミングの知覚に反映されている可能性を 示している。ただし本研究の結果を説明するためには, このメカニズムが,中心視野と周辺視野に提示される 刺激間に時間のずれがあっても機能する必要がある。つ まり,先行刺激(133.3 ms)とそれに続くテスト刺激 (533.3 ms)を統合するという形で運動情報を処理でき るメカニズムでなければならない。 この仮想的なメカニズムでは,周辺視野においては受 容野サイズが拡大していると考えられる。というのも, 先行刺激とテスト刺激を共に周辺視野に提示した場合 は,先行刺激の持続時間が短い時には正のプライミング が観察されたからである(Figure 4)。実験2と実験3を 比較すると,先行刺激とテスト刺激間の空間距離が 6.6 deg の場合,テスト刺激を全く同じ空間位置(周辺 6.0 deg)に提示したにもかかわらず,実験2では正方向 (Figure 4),実験3では負方向(Figure 5)へテスト刺激 が動いて知覚されており,正反対のプライミング効果が 生じている。先に,先行刺激とテスト刺激の間の空間距 離が重要であると指摘したが,距離の効果は先行刺激の 空間位置に依存していることがわかる。周辺視野におい Figure 5. Result of Experiment 3. The graph plots
per-cent response of positive priming as a function of the distance between priming and test stimuli in degrees. Each curve represents the data taken at different prim-ing stimulus duration (133.3 or 766.7 ms). Error bars represent ±1 SEM.
ては,より拡大した受容野を持つメカニズムが働いてい ると仮定すれば,実験2と実験3におけるプライミング の反転を説明することができる。 なお,ここで比較している実験2と実験3の刺激布置 では,テスト刺激の輝度コントラストが異なっている。 前者は0.5に固定されている一方で,後者では運動方向 弁別閾の10倍としている。この違いが,プライミング 効果の反転に寄与した可能性は捨てきれず,今後の検討 が必要である。 一方で先行刺激の持続時間が長い場合は,全実験にお いて,先行刺激とテスト刺激の提示位置が異なるにもか かわらず,ほぼ同じ結果が得られた。フリッカー運動残 効は二次運動や等輝度運動を順応刺激とした場合でも観 察されることから,静止したテスト刺激が順応刺激とは 逆方向に動いて見える古典的な運動残効より高次の処理 レベルが関与していると考えられている(e.g., Nishida, 2011)。視覚情報処理の高次レベルでは受容野がより拡 大していることは,負のプライミングが刺激の空間要因 にかかわらずロバストに知覚されたという結果とも矛盾 しないと考えられる。 本実験結果から,先行刺激の持続時間が長い時に負の プライミング(あるいはフリッカー運動残効)を生み出 すメカニズムと,先行刺激が短い時に正のプライミング (視覚的慣性)を生み出すメカニズムとは,特にその空 間構造が異なることが示唆される。それぞれのメカニズ ムがどのように視覚系に実装されているのか,そして互 いにどのような関係にあるかといった具体的な機序につ いては,今後のさらなる検討が必要である。 引用文献
Allman, J., Miezin, E., & McGuinness, E. (1985). Direction- and velocity-specific responses from beyond the classical receptive field in the middle temporal visual area (MT).
Perception, 14, 105–126.
Anstis, S., & Ramachandran, V. S. (1987). Visual inertia in ap-parent motion. Vision Research, 27, 755–764.
Baker, D. H., & Graf, E. W. (2010). Extrinsic factors in the per-ception of bistable motion stimuli. Vision Research, 50, 1257–1265.
Born, R. T., & Tootell, R. B. H. (1992). Segregation of global and local motion processing in primate middle temporal visual area. Nature, 357, 497–499.
Brainard, D. H. (1997). The psychophysics toolbox. Spatial
Vision, 10, 433–436.
Campana, G., Pavan, A., & Casco, C. (2008). Priming of first- and second-order motion: Mechanisms and neural sub-strates. Neuropsychologia, 46, 393–398.
De Valois, R. L., & De Valois, K. K. (1988). Spatial Vision.
Cambridge, MA: Oxford University Press.
Eifuku, S., & Wurtz, R. H. (1998). Response to motion in ex-trastriate area MST1: Center-surround interactions. Journal
of Neurophysiology, 86, 2011–2028.
Golomb, B., Andersen, R. A., Nakayama, K., MacLeod, D. I., & Wong, A. (1985). Visual thresholds for shearing motion in monkey and man. Vision Research, 25, 813–820.
Ido, K., Ohtani, Y., & Ejima, Y. (1997). Dependencies of mo-tion assimilamo-tion and momo-tion contrast on spatial properties of stimuli: Spatial-frequency nonselective and selective in-teractions between local motion detectors. Vision Research, 37, 1565–1574.
Jiang, Y., Luo, Y. J., & Parasuraman, R. (2002). Neural corre-lates of perceptual priming of visual motion. Brain Research
Bulletin, 57, 211–219.
Jiang, Y., Pantle, A. J., & Mark, L. S. (1998). Visual inertia of rotating 3-D objects. Perception & Psychophysics, 60, 275– 286.
Kanai, R., & Verstraten, F. A. (2005). Perceptual manifestations of fast neural plasticity: Motion priming, rapid motion af-tereffect and perceptual sensitization. Vision Research, 45, 3109–3116.
Levitt, H. (1971). Transformed up-down methods in psycho-acoustics. Journal of the Acoustical Society of America, 49, 467–477.
Livingstone, M. S., Pack, C. C., & Born, R. T. (2001). 2-Dimen-sional substructure of MT receptive fields. Neuron, 30, 781– 793.
McGraw, P. V., & Roach, N. W. (2008). Centrifugal propaga-tion of mopropaga-tion aftereffects across visual space. Journal of
Vi-sion, 8, 1–11.
McKee, S. P., & Nakayama, K. (1984). The detection of motion in the peripheral visual field. Vision Research, 24, 25–32. Mikami, A., Newsome, W. T., & Wurtz, R. H. (1986). Motion
selectivity in macaque visual cortex. I. Mechanisms of direc-tion and speed selectivity in extrastriate area MT. Journal of
Neurophysiology, 55, 1308–1327.
Murakami, I., & Shimojo, S. (1993). Motion capture changes to induced motion at higher luminance contrasts, smaller eccentricities, and larger inducer sizes. Vision Research, 33, 2091–2107.
Murakami, I., & Shimojo, S. (1996). Assimilation-type and contrast-type bias of motion induced by the surround in a random-dot display: Evidence for center-surround antago-nism. Vision Research, 36, 3629–3639.
Nishida, S. (2011). Advancement of motion psychophysics: Review 2001–2011. Journal of Vision, 11, 1–53.
Nishida, S., & Sato, T. (1995). Motion aftereffect with flicker-ing test patterns reveals higher stage of motion processflicker-ing.
Vision Research, 35, 477–490.
西田眞也・竹内龍人・蘆田 宏(2000).8.運動の知覚 日本視覚学会(編)視覚情報処理ハンドブック 朝倉 書店 pp. 335–366.
(Nishida, S., Takeuchi, T., & Ashida, H.)
Orban, G. A., Kennedy, H., & Bullier, J. (1986). Velocity sensi-tivity and direction selecsensi-tivity of neurons in areas V1 and
V2 of the monkey: Influence of eccentricity. Journal of
Neurophysiology, 56, 462–480.
Paffen, C. L. E., van der Smagt, M. J., te Pas, S. F., & Verstraten, F. A. J. (2005). Center-surround inhibition and facilitation as a function of size and contrast at multiple levels of visual motion processing. Journal of Vision, 5, 571–578.
Pantle, A. J., Gallogly, D. P., & Piehler, O. C. (2000). Direction biasing by brief apparent motion stimuli. Vision Research, 40, 1979–1991.
Pavan, A., Campana, G., Guerresch, M., Manassi, M., & Casco, C. (2009). Separate motion-detecting mechanisms for first- and second-order patterns revealed by rapid forms of visual motion priming and motion aftereffect. Journal of Vision, 9, 1–16.
Pelli, D. G. (1997). The VideoToolbox software for visual psy-chophysics: Transforming numbers into movies. Spatial
Vision, 10, 437–442.
Piehler, O. C., & Pantle, A. J. (2001). Direction-specific chang-es of sensitivity after brief apparent motion stimuli. Vision
Research, 41, 2195–2205.
Pinkus, A., & Pantle, A. (1997). Probing motion signals with a priming paradigm. Vision Research, 37, 541–552.
Raymond, J. E., O’Donnell, H. L., & Tipper, S. P. (1998).
Prim-ing reveals attentional modulation of human motion sensi-tivity. Vision Research, 38, 2863–2867.
Sachtler, W. L., & Zaidi, Q. (1995). Visual processing of motion boundaries. Vision Research, 35, 807–826.
Shioiri, S., Ito, S., Sakurai, K., & Yaguchi, H. (2002). Detection of relative and uniform motion. Journal of the Optical
Soci-ety of America A, 19, 2169–2179.
Snowden, R. J., & Milne, A. B. (1997). Phantom motion after effects-evidence of detectors for the analysis of optic flow.
Current Biology, 7, 717–722.
Tadin, D., Lappin, J. S., Gilroy, L. A., & Blake, R. (2003). Per-ceptual consequences of center-surround antagonism in visual motion processing. Nature, 424, 312–315.
Takemura, H., & Murakami, I. (2010). Visual motion detec-tion sensitivity is enhanced by orthogonal induced modetec-tion.
Journal of Vision, 10, 1–13.
Takeuchi, T., Tuladhar, A., & Yoshimoto, S. (2011). The effect of retinal illuminance on visual motion priming. Vision
Research, 51, 1137–1145.
von Grünau, M., & Dubé, S. (1992). Comparing local and remote motion aftereffects. Spatial Vision, 6, 303–314.