Kagamiyama 1-3-1, Higashi-hiroshima,
724
Hiroshima,JAPAN先史ヨーロッパにおける農耕文化圏拡大に関する数理モデル考察
広島大学理学部 瀬野裕美
Amathematical model ofthedispersal of colonies produced bythe stochasticmigrationprocess
dependingonthe totalpopulation of thegroupis considered. The expected velocity of the spatial expanding ofthe settlementrangeofcolonies is analyzed,utilizing the fractal concept appliedto
the pattern of spatialdistributionofcolonies. The model isused toconsiderthespreadmg phenomenon ofearly farIninnginEurope, with the data of neolithic sites with C-14 dates.
INTRODUCTION
Theexpandingof thedistribution
area
ofsome
animalshasbeen theoretically studiedbymathematical models. As for patterns of spatialdistributionand the expanding velocity,some
diffusion models have been appliedtounderstand suchphenomena (AmmermanandCavalli-Sforza, 1984;Martin, 1973; Mosimannand
Martin, 1975; Okubo, 1980; Skellam, 1951). Those phenomenaconsidered by
diffusion models should have such
a
characteristic that the spatial distributioncan
be regarded
as
continuousinspace.
However,for such phenomena that the spatial distribution essentiallyconsistsofa
number of spatially disconnected islands,thatis, colonies, theanalysis bythediffusion model has torequire
some
additionalassumptions,and should be regarded
as an
approximate approach.Inthis
paper,
fortheexpanding of settlementarea
consistingofa
numberofcoloniesispresented,
a
mathematical model ofstochasticmigrationprocesses
isproposed (Bartlett, 1978). Inorderto givetherelation between thenumber of colonies andthesettlement
area
occupied bythem, the fractalconceptisintroduced. Analyzing themodel,
we
derive the expected velocity of the expanding of settlementarea.
The modelis appliedtothedata of neolithic siteswith C-14dates, which
was
usedby Ammerman and Cavalli-Sforza(1984)inorder todiscuss thespreading phenomenon ofearly farming in Europe. The
expanding velocity of the settlement
area
offarming colonies is estimated byour
COLONY PRODUCTION
Free Migration Process: A
new
colony is produced bya
random migrationprocess
in theexistinggroup
ofcolonies,witha
constantmigration probabilityindependentof
any
otherparameter(Bartlett, 1978). That is,theprobabilityofthe production ofa new
colony isconstant,independent ofany
other parameters.Now,itisassumed thatthe colony doesnotbecomeextinct
once
itis produced. Under theseassumptions, the followingmodelcan
be defined:$\frac{d}{dt}P(k, t)=-\lambda P(k, t)+XP(k-1, t)$ (1)
$P(k, 0)=6_{k)}$,
where
$\lambda dr$
.
theprobabilityofproduction ofnew
colony during $(t, t+dt)$$P(k, t)$
:
theprobability of$k$colony productions during time-period $(0, t)$.
$6_{k0}$isthe kronecker’s delta
so
that the initialconditionmeans
thatthereisno
colony production at$t=0$
.
This colony production system results in the Poissonprobability distribution $P(k, t)$
:
$P(k, t)= ff^{\lambda t}\frac{(\lambda t)^{k}}{k!}$ (2)
Theexpectednumberofcolonies produced during $(0, t)$ is
$k h=\sum_{k=0}kP(k, t)=\lambda t$, (3)
and theexpectedtimeof the k-th colony production is
$rangFig$
.el.byScheemiaz
線
crfietni
躍註灘欝露盤
ent
explanation,seetext.
Size-dependent Migration Process: The migration probability is assumedto
depend
on
thetotal population of thegroup
(Fig. 1). Thismeans
thatthecolony production isenhancedmore
andmore
as
the totalpopulation becomes larger. Under thisassumption,we
consider the followingmodel:$\frac{d}{dt}P(k, t)=-\}\iota N(t)P(k, t)+\mu N(t)P(k-1, t)$ (5)
$P(k, 0)=$ り,
where
$N(t)$
:
the totalpopulation sizeof thegroup
of coloniesattime$t$$\mu N(t)dt$; theprobabilityof production of
new
colony during time-period$(t, t+dt)$.
This colony production system in time$t$results in thePoissonprobability
distribution$P(k, 7)$ in time$T$
:
$P(k, T)=e-\downarrow 1T_{\frac{(\mu\tau Y}{k!}}$ (6)
where the time$T$is
now
transformed from time$t$as
follows:$T=T(t)= \int_{0^{t}}N(\tau X1\tau.$ (7)
Since
a
colony does notbecomeextinct afterits
production,we
find that $Tarrow\infty$as
$tarrow\infty$.
Theaboveresultintime$T$coincides with that for thepreviouscase, thatis, for the
case
offreemigrationprocess.
Therefore, theexpected numberof colonies produced during $(0, t)$ is$\Psi\lambda=\sum_{k=0}^{\infty}kP(k, T)=[\iota T=\mu\int_{0}^{t}N(\tau)d\tau$, (8)
and the expectedtime of the k-th colony production is
$t^{\tau}k=\int_{0}^{\infty}\tau P(k-1, \tau)\}\downarrow d\tau=\frac{k}{\downarrow 1}$
.
(9)Then, the expectedtimein$t$
can
beobtained through the following relation:$t k=T^{-1}(\{\phi=T^{-1}(\frac{k}{\mu})$
(10)
where$T^{-1}$ denotes theinverse function of$T=T(t)$
.
EXPANSION OF SETTLEMENTAREANext,
we
consider the settlementarea
ofthegroup
of colonies. The settlementarea
attime$t$correspondstothearea
thathas been occupiedby thoseexistingcoloniesatthetime. We characterize thesettlement
range
bytheminimaldiameter,
say
$R$, whichcan
include all existing colonies.In the
case
when thesettlementarea
expands inevery
direction with thesame
probability,the shape of thesettlementarea can
beapproximated by thedisc,andtherefore, when the spheric nature of the earth
can
benegligibleandbeapproximated well bythe plane, the
range
$R$approximatelyhas thefollowingrelation withthetotal number of colonies $M;M\propto R^{2}$
.
However, sincethe expanding of the settlementarea
is constrained by geography,climate, culturalfactors, etc.,theshape is in generalpossibly inhomogeneous in direction. Itis
likely that the shape has
a
fractal
nature (fortheconcept of’ffactal’, see, forinstance, Mandelbrot, 1982). To deal withsuch cases,
we
assume
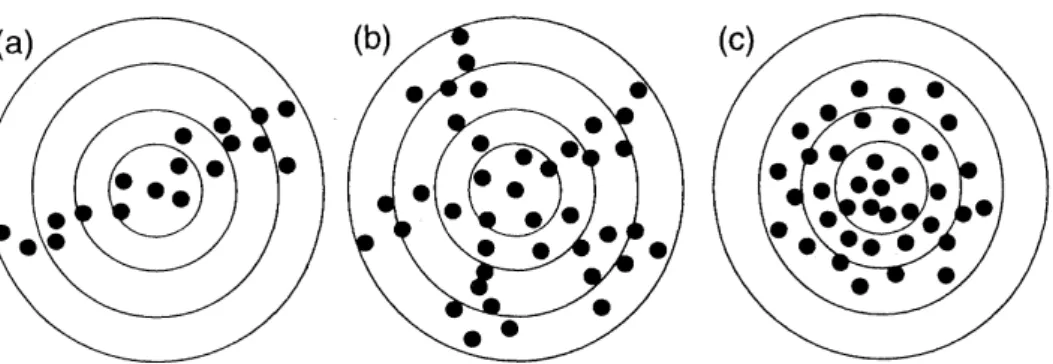
thegeneralizedFig.2. Illustrative explanation of therelation of the fractal dimension$d$tothe spatial pattern of colonydistribution. Each black disc shows each colony. (a)$d\sim 1;(b)1<d<2;(c)d\sim 2$.
$M\propto R^{d}$ $(1 \leq d\leq 2)$, (11)
wherethe
power
$d$charactenizes the spatial pattern of the settlementarea
occupiedby colonies(Fig. 2). Itiscalled cluster dimension
or mass
dimension,whichisone
offractal
dimensions. When$d\sim 2$, the spatialdistribution of coloniescan
be approximated bya
disc. When$d\sim 1$, thedistributionisone
dimensional, that is, the coloniesare
arrayedon
a
curve.
Forexample, the lattercase
may
correspondtothe
case
whenthecoloniesare
located alonga
river.Through therelation (11),
we
can
consider thevelocity of the expanding of the settlementrange.
That is, thevelocity $V$is given by$V= \frac{dR}{dt}\propto\frac{d}{dt}(M^{1/d})=\frac{1}{d}\cdot M^{(1-yd}\cdot\frac{dM}{dt}$
.
(12)Sincetheexpectedtota1 numberof coloniesattime$t$is given by$kh$, theexpected
range
of thesettlementarea
isproportionalto$k\Psi^{d}$.
Therefore,we
consider theexpected velocity $V_{t}$ attime$t$
as
follows:$\ovalbox{\tt\small REJECT}\propto\frac{1}{d}\cdot\phi h^{(1-dy_{d}\ovalbox{\tt\small REJECT}}d_{dt}$
.
(13)Forthe
case
ofthefreemigrationprocess,
the expected velocityis$\overline{V_{t}}\propto\frac{\lambda^{1/d}}{d}t^{(1-\nu d}$ , (14)
$\overline{V_{t}}\propto\frac{\mu^{1/d}}{d}\cdot N(t)$
.
$[ \int_{0^{t}}N(\tau)d\tau]^{1-dy_{d}}$.
(15)LOGISTICGROWTH OFPOPULATION
As
an
exampleof the size-dependentmigrationprocess,
we
deal withthecase
whenthetotal population size of thegroup
of coloniesgrows
in the logisticmanner
(Fig. $3(a)$):$N(t)=N(0) \cdot\{(1-\frac{N(0)}{K})e^{-}’+\frac{N(0)}{K}\backslash /^{-1}$
, (16)
where$\epsilon$ isthe intninsicgrowthrateofthepopulation and$K$isthecarrying
capacityforthetotal population ofthe
group.
In thiscase,some
fundamental calculations show(17) $k)_{l}= \frac{\mu}{\epsilon}\cdot K\cdot\ln\{\frac{N(0)}{K}\cdot(\not\in-1)+1\}$
$\#k=\frac{1}{\epsilon}\ln\{\frac{K}{N(0)}\cdot(e^{(\epsilon 1\downarrow\downarrow)k/\kappa_{-1)+}}1\}.$
(18)
Generic feature oftheseexpectedvaluesis shownin Fig. 3(b) and Fig. 3(c). The
time interval betweenthe nearest twocolony productions is givenby
$\#k+1-\#k=_{\frac{1}{\epsilon}}\ln\{\frac{e^{\epsilon/(\mathscr{O}}+\frac{N(0)}{K}(1-\frac{N(0)}{K})\cdot e^{-(\epsilon 1\downarrow\downarrow)k/K}}{1+\frac{N(0)}{K}(1-\frac{N(0)}{K})\cdot e^{-(\epsilon 1\downarrow\iota)k/K}}\{$
.
(19)Thisvalue tendsto
a
constant $l/pK$as
$karrow\infty$, whichmeans
thatthecolonyproductionisexpectedto
occur
periodically. In addition, from (17) and (18), for sufficiently large$t$and sufficiently large$k$,(a)Typica1time-development of logistic populationgrowth;(b)Typical Typical
lk
$\cdot$$\phi h^{\sim}t$ (20)
$\#k^{\sim k}$
.
(21)Thatis, these expected valuesincrease linearly in
a
sufficientlygrown group,
whichis the
same as
for thecase
of the freemigrationprocess.
In this case, the expanding
way
of settlementrange
essentiallydependson
thefractal dimension$d$(Fig. $4(a,$$b)$). Theexpected velocity of expanding of settlement
range
isexpressedas
follows:$\overline{V_{t}}\propto\frac{\epsilon}{d}(\frac{\mu K}{\epsilon})^{1/d}\frac{N(0)}{K}\cdot\frac{e^{\sigma}}{\{\frac{N(0)}{K}\cdot(\theta-1)_{/}+1^{\backslash }[\ln\{\frac{N(0)}{K}\cdot(\theta-1)+1^{1_{(}}]^{1-11d}}$
.
(22)For $1<d\leq 2$, thisexpected velocity decreasesto
zero
ata
sufficiently largetime(Fig.$4(c,$$d)$). This
means
that, fora
sufficientlygrown group,
thevelocity of theexpanding of settlement
range
isvery
small,whilethe number of colonies continuously increases; thatis, thenew
coloniestend to beproduced within the vacantareas
among
the pre-settledcolonies. Ontheotherhand, thetime-development oftheexpectedvelocity intheearlier period depends
on
theinitialpopulation sizeofthe
group
(Fig. $4(d)$). Inthecase
when the initial populationissufficiently large, the expectedvelocity monotonically decreases in time, while in the
case
when itis small, theexpectedvelocity increasesin theearlierperiod and decreases after peaking. Analytically,if thefollowing conditionis satisfied, the formercase
occurs, and otherwisethelatter(Fig. $4(d)$):Fig. 4. Inthesize-dependent immigrationprocessmodel for the logistic population growth, thecontnibution ofthefmctal dimension$d$and the initialpopulationsize
$N(O)/K$to: $(a, b)$thetime-development of the expected settlementrange$R$,thatis,
$kt^{/d}$
:
$(c, d)$thetime-development of the expected velocity17.
For(c),$N(O)/K=$ 0.1,and for(d),$d=1.5$. The graph shape of theexpected velocity $\overline{V_{t}}$depends on$d$ and$N(O)/K$asshown totallyinthefigure attachedto(d).
This condition
can
beeasily derived by examining thesignof the t-derivative of(22).
Intheperiod when thepopulation size$N(t)$ is sufficientlysmall, the
population growth
can
be well-approximated by exponential growth. Inthisperiod,the
same
argumentas
abovegives theapproximateresultson
thebehavior ofcolonydispersalas
follows:$N(t)\approx N(0ffl$ (24)
$k h^{\approx}\frac{\mu}{\epsilon}N(0X\not\in-1)$ (25)
$\#k^{\approx}\frac{1}{\epsilon}\ln(\frac{\epsilon/\mu}{N(0)}k+1)$ (26)
$\#k+1-\#k^{\approx}\frac{1}{\epsilon}\ln(\frac{N(0)+\epsilon(k+1y\mu}{N(0)+\epsilon H\mu})$ (27)
longitude
Fig. 5. 106neolithic European sitesinthegeographiccoordinates,used by AmmermanandCavalli-Sforza(1984). The blacksquareindicates the oldestsite,
Aswad(9690B.C.,C-14date). Black discs are forthosesites before5800B.C.
(C-14date),and whiteonesforthose after5800B.C.
In this case, theexpectedvelocity increasesexponentially while the number of coloniesgrows exponentially.
SPREAD OFEARLYFARMING INEUROPE
Ammerman and Cavalli-Sforza(1984) calculated theisochron
map
of the spread of early farming inEurope from the data of106
neolithic European sites withC-14 dates(9690 B.C.
-4160
B.C.). Thecomputer-generatedisochronmap
givesthe impressionthatearly farming might have spread in
a
spatiallycontinuousmanner
in Europe. This isan
approximationtothe spatial spread through the analogy of diffusionprocess. However,in contrast to thespatial spread ofvariousspecies ofanimals, insects, and plants, the spatial spread of
a group
of humans frequentlyinvolves theproduction spatially disconnectedunits,thatis,colonies. The spatial distribution expandsessentially bya
seriesof productions ofnew
colonies. The spread of early farming in Europe, dealt with by Ammerman and Cavalli-Sforza(1984),
can
be regardedas
sucha
case.
Inthis section,
we
applyour
mathematical model described aboveto thedata and estimate the parameters of the model totrytodiscuss
some
features of the spread ofearly farming in Europe.As forthe
way
ofpopulation growth,we
assume
theexponentialone
givenby (24). This is appropriate inthe
case
when the population growthdoes nottime
Fig.6. Time-development of the number of neolithic sites,which iscumulatedafter the oldestsite,Aswad(9690B.C.,C-14date). Timeaxisshowsthe C-14 date passed$aRer$9690B.C. All 106neolithic sitesareplotted forthedata of Ammerman
andCavali-Sforza(1984). $k1$ curveforthe exponential growthinthe
size-dependentmigrationprocessmodelisoverlaid,fitto76data of neolithicsites
$($before5800B.C.(C-14 date). $\mu N(O)/\epsilon=3.352$and$\epsilon=8.233x10^{-4}$in
(25).
Fig.
5
showsthe106
neolithic European sites in the geographic coordinatesusedbyAmmerman andCavalli-Sforza(1984). Theirspatialdistribution
seems
toshow inhomogeneity in direction. Beginning with the oldestsite, Aswad(9690 B.C.;
33.
$36N,$ $36.30E$),we
countthecumulative number ofcolonies inorderofdescending C-14 date
as
shownin Fig.6.
Plotsin the figure indicate that thecontinuityof the time-development of the number of coloniesseems
tobreakataround3800 years
after Aswad (i.e.,around5800
B.C.). Thus,we use
only the76
data before5800
B.C.,up
tothesite Reichtett(5940B.C.;48.
$6N,$$7.75E$).SinceFig.
6
can
be regardedas
coIrespondingtothe time-developmentof$k)_{t}$ ,
we
trytofit$\ell_{\{}$}
$i$ given by (25)to the data. Theresultis overlaid in Fig.
6.
Theestimated parameters result in$\mu N(O)/\epsilon=3.572;\epsilon=8.233^{x}10^{-4}$
.
Next,
we
try toestimate the ffactal dimension$d$thatcharacterizes thepatternof spatial distribution. From(11), the
range
$R$ of thesettlementarea
andthe number$M$of colonieswithin ithavetherelation: $\log M=d\cdot\log R+const$
.
Therefore,
we
can
estimate$d$fromthe slope ofthe linefitto theplots of log$M$against$\log R$
.
Thediametercan
be calculated from the data of the locations ofneolithicsites (Fig. 7). We
use
thegyration-radius methodtoestimatetheparameter$d$(as forthe method, see, forinstance, Mandelbrot, 1982). All
76
sitesbefore
5800
B.C.are
considered. The numberofsitesdistributed withinthedisc centeredat theoldest site, Aswad,iscounted. For disc radius large enoughtotime
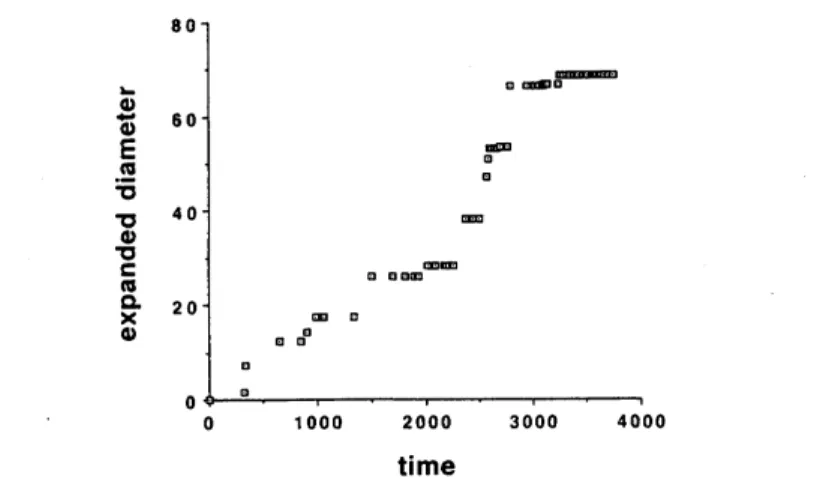
Fig.7. Expandeddiameterof thesettlementrange. Time axis shows the C-14 date passed after9690B.C.
$\underline{\frac{\wedge oc\wedge}{*\vee oooo}}$
log(r) Iog(expanded diameter)
Fig. 8. (a)Number ofsiteswithin thedistance$r$from the oldestsite, Aswad,in log-log coordinates. For the distance$r$thatcontainsmorethan 10sites,plots fit wellto theline with the slope 1.671. (b)Relation between the expanded diameter of settlementareaand the number of coloniesinlog-log coordinates. The overlaidline
indicates theslope 1.671. Theunitof measured distance is conventionallyselected.
radius
can
befittedwellbya
straight line with slope 1.671,as
estimatedby the least-square method(Fig. $8(a)$). Hence,the spatialdistribution ofneolithicsitesisestimated to have thecharacteristicfractal dimension$d=1.671$
.
Since thediameter and the number of neolithic sites
are
time-dependent,it islikely that thepaIameter$d$might change in time in the periodconsidered
now.
However,as
Fig.8(b) shows,the time-dependent relation between the expanded diameter and the
number of colonies in log-logcoordinates, theestimated$d=1.671$
even
holds well. Therefore,we
dealwith$d$as
time-independent constant:$d=1.671$.
From (28) withthese estimated parameters, the time-development ofthe
expected velocity of the expanding ofsettlementrange
can
bedrawn,resulting in Fig. 9(a). Itcan
beseen
that the velocity isrelatively small in the first century after Aswadandthenincreasesexponentially. In Fig. 9(b), thesame
expectedtime #colony
Fig. 9. Time-development of the expectedvelocity(28)of the expanding of the settlementarea,for$\mu N(O)/\epsilon=3.352;\epsilon=8.233x10^{-4};d=1.671$. $(a)$
time-development of the expected velocity(28); (b)the expected velocity(28)againstthe expected number of colonies(25).
velocityisplotted against thetime-development of the expected number of
colonies, which follows(25). As thenumberof colonies becomes sufficiently
large, thevelocityofthe expanding ofthe settlement
area
increases. REFERENCESAmmerman,A. J.andCavalli-Sforza,L. L. (1984) Neolithic Transition and The Genetics
of
Populations in Europe, PrincetonUniversityPress, Princeton,New Jersey.
Bartlett,M.S.,F.R.S. (1978) AnIntroductiontoStochasticProcesses, CambridgeUniversity
Press,Cambridge.
Britton,N. F. (1986)
Reaction-Diffision
Equations and Their ApplicationstoBiology, AcademicPress,London.
Mandelbrot,B. B. (1982) $Th\ell$Fractal Geometry ofNature, Freeman,SanFrancisco.
Martin,P. S. (1973) The discovery of America. Science. 179,969-974.
Mosinam,J. E.andMartin,P. S. (1975) Simulating overkill by paleoindians. Am SCL 63,
304-313.
Murray, J.D. (1989) Mathematical Biology, Springer-Verlag, New York.
Okubo,A. (1980)
Diffision
and Ecological Problems: MathematicalModels, Springer-Verlag,New York.