日本 応用 動 物 昆 虫 学会 誌(応 動 昆) 第28巻 第2号:68-74 (1984)
カ イ コ の 胚 子 形 成 過 程 に お け る タ ン パ ク 質 合 成 の 変 動1)
河 口
豊 ・藤 井
博
九州大学農学部
Changes in Protein Synthesis during Embryonic Development of the Silkworm, Bombyx mori
(Lepidoptera:
Bombycidae). Yutaka KAWAGUCHI
and Hiroshi FUJII (Laboratory of Sericulture,
Department
of Agriculture, Kyushu University, Hakozaki, Higashi-ku, Fukuoka 812, Japan).
Ja p. J. appl. Ent. Zool. 28: 68-74 (1984)
Protein synthesis in developing embryos of Bombyx mori was studied by means of microinjection
experimens using tritium-labeled precursors and actinomycin D (AD). The incorporation of
3H-leucine into the acid-insoluble fraction was shown to be most active on the first day (stages 3-4)
and during the fifth to ninth days (stages 6-20) after oviposition. In the diapausing egg the
incor-poration of 3H-leucine during stages 3 to 4 was not affected by AD, while the incorincor-poration at the
later stages was appreciably inhibited by AD. Two-dimensional gel electrophoresis followed by
autoradiography using 35S-methionine demonstrated the existence of stage-specific peptides which
were synthesized at different phases of development. The present results suggest that the
trans-lational event in the early stages of embryogenesis depends on pre-existing maternal mRNAs, and
at the later stages of the embryonic development it depends upon newly synthesized mRNAs.
受 精 か ら 胚 子 形 成 に 至 る 胚 発 生 の 過 程 と そ の 機 構 を タ ンパ ク質 や 核 酸 の 合 成 活 性 の 変 化 を 指 標 に し て 解 明 し よ う と す る 試 み は 主 と して ウ ニ や カ エ ル を 用 い て 詳 細 に 行 わ れ て き た(DAVIDOSON, 1976)。 そ の 中 で も 母 性mRNA の 存 在 と そ の 機 能 い か ん に つ い て は 初 期 発 生 の 機 構 を 解 く 重 要 な 鍵 と し て 注 目 を 集 め,多 くの 研 究 が 行 わ れ た (RAFF, 1980)。 そ の 結 果,卵 形 成 中 に 作 ら れ た 母 性 mRNAは タ ン パ ク 質 と結 合 し てmRNPと な り マ ス ク さ れ た 形 で 卵 内 に 貯 蔵 さ れ,受 精 後 胚 子mRNAの 合 成 が 開 始 さ れ る ま で の 胚 発 生 に 関 与 す る こ と,さ ら に mRNPか ら抽 出 したmRNAはin vitroに お い て 翻 訳 活 性 能 を も つ こ と が 明 ら か に さ れ た 。 一方,昆 虫 に お い て はDrosophila melanogastar (GOLDSTEIN and SNYDER, 1973; LOVETT and GOLDSTEIN, 1977; MERMOD et al., 1980)やSmittia sp. (JACKLE, 1979; JACKLE and KAL-THOFF, 1979)あ る い はEuscelis plebejus (SCHMIDT and JACKLE, 1978)を 用 い た 研 究 か ら母 性mRNAは 卵 形 成 後 期 に 合 成 さ れ,mRNPの 形 態 を と りpoly (A)部 分 を も ち,受 精 完 了 後 に お い て 翻 訳 活 性 を 発 現 す る こ と な ど が 明 ら か に さ れ て い る 。 カ イ コで は 休 眠 卵 と発 生 中 の 卵 に お け るpolysomeの 活 性 に つ い て 比 較 分 析 し
たSZY-SZKO and LASSOTA (1977)やGRZELAK et al. (1979)の
研 究 は あ るが,タ ンパ ク質 合成 か らみ た胚 発 生 に つ い て
の研 究 は 少 な い 。
著 者 らは胚 発生 過程 に お け る遺 伝 子 の 発 現 を 翻 訳 レベ ル で解 明す る 目的 で 研究 を 行 って い るが,ま ず カ イ コ未
受 精 卵 か らRNP顆 粒 を抽 出 し,poly (A)部 分 を 有 す る
母 性mRNAが 存 在 し,し か もin vitroにお い て 翻 訳 能 を
もつ こ とを 明 らか に した(藤 井 ・河 口,1982)。 こ の 結 果 を基 礎 に して さ らに胚 子形 成 過 程 に お け る タ ンパ ク質 合 成 の変 動 に つ い て 解 析 を 行 った の で こ こに 報 告 す る。 本 文 に 入 るに先 立 ち,著 者 らの 研 究 に 対 し終 始 懇 切 な る指 導 を い た だ きか つ 本 文 校 閲 の 労 を と られ た 九 州 大 学 農 学 部 坂 口文 吾 教 授 に 衷 心 よ りお 礼 申 し上 げ る。 また, 本 研 究 遂 行 上 種 々の 助 言 を いた だ いた 九 州 大 学 農 学 部 家 蚕 遺 伝 子 実 験 施 設 の 土 井 良 宏 教 授 な らび に木 原 始 氏 に お 礼 申 し上 げ る。 材 料 と 方 法 供 試 蚕 は 九 州 大 学 農 学 部 家 蚕 遺 伝 子 実 験 施 設 保 存 の d41系 統 で あ る。 産 下 後6時 間(stage 1∼2),12時 間(stage 2),産 下 後20時 間 の カ イ コ卵 を 比 重1.075 の塩 酸 溶 液(溶 液 は46℃ に 加温)中 で5分 間 浸 積処 理 (即時 浸 酸 法 に よ る人 工 孵 化 処 理)を 施 した1日(stage 1) 本 研 究 に 要 した 経 費 の一 部 は昭 和54年 度 文 部 省 科 学 研 究 費一 般 研 究(C) , No.356245に よ った. 本 報 告 の一 部 は 昭 和53, 54年 度 日本 蚕 糸 学 会 九 州支 部 大 会(1978年11月,大 分;1979年11月,鹿 児 島)で 口頭 発 表 した. 1983年11月18日 受 領 (Received November 18, 1983) -68
3∼4), 3日(stage 9), 4日(stage 11), 5日(stage 13), 7日(stage 17), 9日(stage 20)の 人 工 孵 化 処 理 卵,お よ び 人 工 孵 化 処 理 を し な い1, 2, 3, 4, 5, 7日 の 休 眠 卵 を 用 い て 実 験 を 行 っ た 。 雌 蛾 の 産 卵 時 間 は30分 間 に 限 定 し,産 下 後 の 卵 は す べ て25℃ に 保 護 した 。 胚 子 の 発 育 階 梯 は 大 槻(1970)の 分 類 に 従 っ た 。 1. 卵 へ の 放 射 性 ア ミ ノ 酸 と ア ク チ ノ マ イ シ ンDの 投 与 産 下 後6, 12時 間,1日,2日,3日,4日 の 卵 に は 背 側 に 細 い タ ン グ ス テ ン針 で2か 所 穴 を あ け る 。 片 方 の 穴 に ガ ラ ス 細 管 で 作 成 し た 注 射 針 を 挿 入 し,3H-ロ イ シ ン(0.02μCi/0.04μl/卵)を 投 与 し た 。 他 方,ア ク チ ノ マ イ シ ンD処 理 区(以 後AD処 理 区 と 記 す)に は3H-ロ イ シ ン(0.02μCi/0.02μl/卵)とAD(0.02μg/0.02 μl/卵)と を 同 時 に 投 与 し た 。 そ の30分,1, 3, 6, 9 時 間 後 に10%冷 ト リ ク ロ ル 酢 酸 を 用 い て 卵 の 酸 不 溶 性 分 画 を 得 た 。 分 画 中 の 放 射 活 性 は トル エ ン:ト リ ト ンX 100:PPOの 溶 媒 系 を 用 い て ベ ッ ク マ ン社 製 液 体 シ ン チ レ ー シ ョ ン ス ペ ク ト ロ フ ォ トメ ー タLS-315Tで 測 定 し た 。 2. 泳 動 用 試 料 の 調 整 産 下6時 間,人 工 孵 化 処 理 を 施 し た1日,3日,7日 の 卵 に 前 述 の 方 法 に よ り35S-メ チ オ ニ ン(0.18μCi/ 0.04μl/卵)を 投 与 した 。3時 間 後,卵 殻 を 除 き 卵 内 容 物 を9.5M尿 素,2%NP-40, 2%ア ン フ ォ ラ イ ト (LKB社 製),5%メ ル カ プ トエ タ ノ ー ル 混 合 液 に 溶 解 さ せ,10,000×9, 5分 間 遠 心 し た 上 清 を 泳 動 用 試 料 と し た 。 3. 2次 元 電 気 泳 動 法 電 気 泳 動 はO′FARRELL (1975)の 方 法 に 準 じて 行 っ た 。 す な わ ち,1次 元 目 に は キ ャ リ ア ー ア ン フ ォ ラ イ ト に よ りpH4.0∼8.0のpH勾 配 を 形 成 し た5%ア ク リ ル ア ミ ドゲ ル を 支 持 体 とす る デ ィ ス ク 電 気 泳 動,2次 元 目 は2% SDSを 含 み 濃 度10∼16%の 直 線 的 勾 配 を 形 成 し た ア ク リ ル ア ミ ド ゲ ル に よ る ス ラ ブ 電 気 泳 動 で あ る 。 等 電 点 は1次 元 目 の 泳 動 終 了 後 ゲ ル を5mm幅 の 切 片 に 切 り出 しpHメ ー タ ー で 測 定 し た 。 タ ンパ ク質 の 分 子 量 は 分 子 量 既 知 の タ ンパ ク質 マ ー カ ー(E. coli RNA polymerase α-subunit, 39kd; β-subunit, 165kd; β′-subunit, 155kd; bovine serum albumin, 68kd; try-psine inhibitor, 21.5kd)を 同 じ条 件 で 泳 動 しそ の 相 対 的 移 動 度 よ り 作 成 し た 標 準 線 か ら 求 め た 。
4. オ ー トラ ジ オ グ ラ フ 法
泳 動 終 了 後 の ゲ ル は10%ト リ ク ロ ル 酢 酸 溶 液 で 固 定
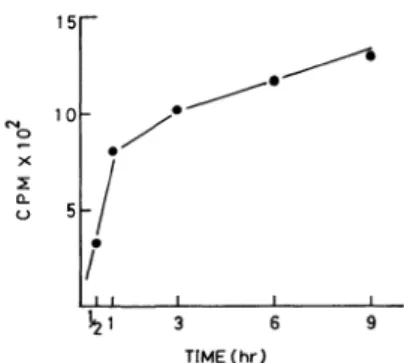
Fig. 1. Time course of 3H-leucine incorporation into the acid-insoluble fraction of the Bombyx egg. Egg: 24hr after oviposition.
な らび に 洗 浄 し,7%酢 酸 に よ り脱 色 した 染 色済 の ゲル
を た だ ち に フ ィル ム状 に 乾 燥 し,Kodak X-OMAT film
を 密着 させ,冷 暗所 に て5日 間露 出 した後 現象 ・定 着 し た。 結 果 1. 胚 子 形 成 過 程 にお け る卵 の酸 不 溶 性 分 画 中へ の 3H-ロ イ シ ン の 取 り込 み 3H-ロ イ シ ン投 与後 に お け る卵 の 酸 不 溶 性 分 画 中 に 検 出 され る放 射 活 性 の経 時 的変 化 をFig. 1に 示 す 。3H-ロイ シ ンを産 下 後1日 の卵 に投 与 し30分,1, 3, 6, 9 時 間 後 に卵 内容 物 の酸 不 溶 性 分 画 中 の放 射 活 性 を調 べ た とこ ろ,投 与 後1時 間 ま で は急 速 に,そ れ 以 後 は徐 々に 上 昇 し,少 な く とも投 与 後9時 間 ま で卵 は3H-ロ イ シ ン を 酸 不 溶 分 画 中 に取 り込 む能 力 を維 持 して い た。 した が って 以 後 の取 り込 み 実 験 で は,投 与6時 間 後 の 酸 不 溶 性 分 画 中 へ の取 り込 み 量 に つ い て比 較,分 析 した 。 胚 子 発 育 経 過 に 伴 う卵 の 酸 不 溶 性 分 画 中 へ の3H-ロ イ シ ン取 り込 み 量 の 変 化 をFig. 2に 示 す。3H-ロ イ シ ンの 取 り込 み は 産 下後6時 間 か ら1日 に か け て増 加 す る が, 2日 に な る と一時 減 少 した。 しか しな が ら,人 工 孵 化 処 理 卵 は3日 以降 再 び増 加 を開 始 し,5日 か ら7日 へ と胚 子 発 育 の進 行 に伴 い取 り込 み は急 激 に上 昇 して最 高 値 に 達 した。9日 に お い て も7日 とほ ぼ 同 じ ぐ らい高 い値 を 維 持 して いた 。 一 方,人 工 孵 化 処 理 を し な い 休 眠 卵 で は,人 工 孵 化 処 理 卵 にみ られ る3日 以 降 の増 加 は ま った く認 め られ ず,き わ め て 低 い 値 で 推 移 した 。 ち な み に 産 下 後2日 の 卵 に お け る3H-ロ イ シ ンの 取 り込 み 能 は 人 工 孵 化処 理 卵 の ほ うが 人 工 孵 化 処 理 を しな い卵 よ りも大 で あ った 。
70 河 口 豊 ・藤 井 博
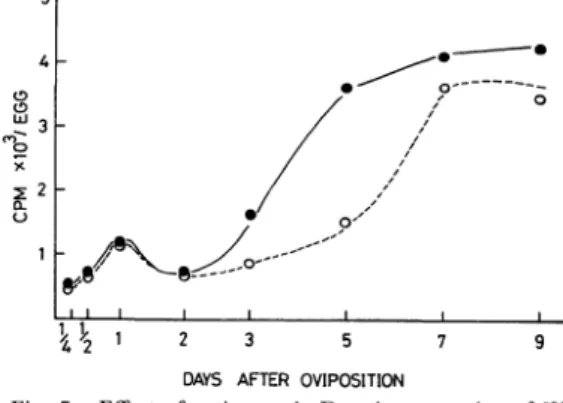
Fig. 2. Changes of incorporation of 3H-leucine into the acid-insoluble fraction of the Bombyx egg through-out the embryogenesis. Eggs that were incubated at 25•Ž were collected six hours after injection. Artificial hatching treatment (HCl treated) was performed at twenty hours after oviposition. •œ: HCl-treated, and non-diapausing egg, •›: dia-pausing egg in which HCl treatment was omitted.
Fig. 4. Time course of 3H-leucine incorporation (2•~10-10ƒÊg/egg) into the acid-insoluble fraction of the Bombyx egg in the presence of actinomycin D. The age of the egg was the same as in Fig. 3.
2. 胚 子 形成 過 程 に お け る 卵 の 酸 不 溶 性 分 画 中 へ の 3H-ロ イ シ ン取 り込 み に 及 ぼ す ア ク チ ノマ イ シ ン Dの 影 響 産 下 後5日 の 人工 孵 化 処 理 卵 に3H-ロ イ シ ンの み を投 与 した6時 間 後 の 対 照 区 に お け る取 り込 み に対 す る3H-ロ イ シ ン とADと を 同 時 に 投 与 したAD区 に お け る取 り込 み の 割 合 か ら阻 害 率 を 求 めADの 影 響 を 調 べ た (Fig. 3)。 卵1粒 当 りADを2×10-5μg与 え る と14%, 2×10-4μgで28%, 2×10-3μg 45%, 2×10-2μgで は 58%も の 阻 害 効 果 が 認 め られ,AD量 の 対 数 に 比 例 して そ の阻 害 効 果 は 高 くな った 。 次 に3H-ロ イ シ ン 取 り 込 み に及 ぼ すADの 阻害 作 用 の経 時 的 変 化 をFig. 4に 示
Fig. 3. Effect of actinomycin D at varying doses on the incorporarion of 3H-leucine into the acid-insoluble fraction of the Bombyx egg. Five days old eggs were used.
Fig. 5. Effect of actinomycin D on incorporation of 3H-leucine into the acid-insoluble fraction in the Bombyx egg throughout the embryogenesis. Eggs were collected six hours after injection. •œ: 3H-leucine , •›: 3H-leucine+actinomycin D. す 。AD(2×10-2μg/1粒)投 与後 作用 時 間 の 経 過 に 伴 い 阻害 効 果 は 徐 々に 高 くな り,投 与6時 間後 に は 阻 害 率 56%と な った 。 しか し投 与後9時 間 目に な る と や や 低 下 した。 上 述 の結 果 に基 づ き胚 子 形 成 中 の 卵 の3H-ロ イ シ ン取 り込 み に対 す るADの 影 響 は,濃 度2×10-2μg (Fig. 3),作 用 時 間6時 間(Fig. 4)の 条件 で 分析 す る こ と と した。 そ の結 果 をFig. 5に 示 す 。AD投 与 区 に お け る3H-ロ イ シ ンの取 り込 み は産 下 後6か ら12時 間,さ らに1日 へ と発 育 の進 行 に伴 っ て 徐 々 に増 加 し,2日 目 に な る と一 時 減 少 した。 この 間 に お け る取 り込 み の上 昇 は対 照 区 の それ と比 較 して ほ とん ど差 異 は 認 め られ な か った 。 一 方,産 下 後3日 か ら5日 お よび7日 に か け て対
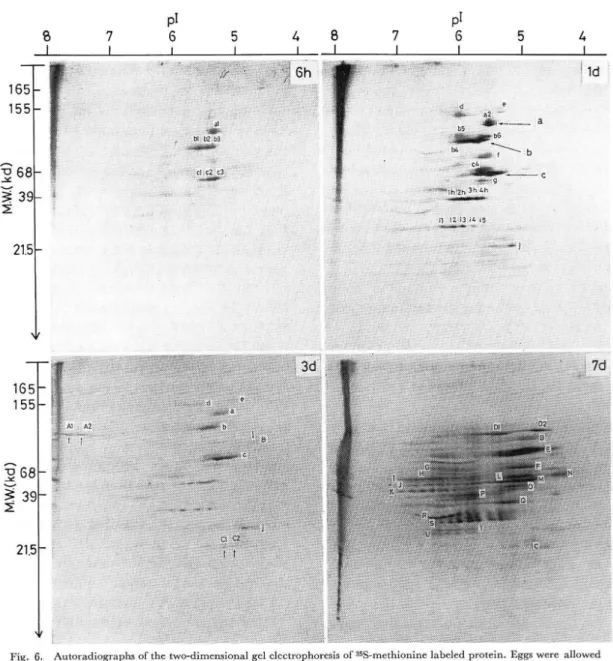
Fig. 6. Autoradiographs of the two-dimensional gel electrophoresis of 35S-methionine labeled protein. Eggs were allowed
a three-hour incorporation of the labeled precursor. 6th, 1d, 3d and 7d refer to six hours, one-, three- and
seven-day old eggs, respectively. One- to seven-day old eggs were previously subjected to the artificial hatching
treatment at twenty hours after oviposition.
照 区 で は取 り込 み が 急激 に上 昇 す るの に 対 し,産 下 後3 日,5日 のAD区 に お いて は3H-ロ イ シ ン の 取 り込 み は対 照 区 の よ うな 著 しい上 昇 は 認 め られ ず 低 い 値 で あ っ た 。7日 のAD区 で は か な り取 り込 み が 増 加 し,9日 のAD区 に お い て も7日 と同様 の 取 り込 み が 認 め られ た。 しか し,い ず れ の 場 合 も対 照 区 よ り高 くな る こ と は な か った 。 3. 胚 子 発 育 に伴 う卵 タ ンパ ク質 の2次 元 電 気 泳 動 オ ー トラ ジ オ グ ラフ 像 の 解 析 オ ー トラ ジ オ グ ラ フ法 に よ る卵 タ ンパ ク質 の2次 元 電 気 泳 動 像 をFig. 6に 示 す 。X線 フ ィル ム上 に現 わ れ た 黒 色 の ス ポ ッ トは タ ンパ ク質(正 確 に はポ リペ プ タ イ ド で あ るが,こ こ で は タ ンパ ク質 成 分 と して表 示 す る。) 中 に取 り込 まれ た35S-メ チ オ ニ ンの放 射 活 性 に 基 づ く
72 河 口 豊 ・藤 井 博 も の で あ り,新 生 タ ン パ ク 質 の 存 在 を 示 し,ス ポ ッ トの 大 小 は 合 成 さ れ た タ ン パ ク 質 の 量 を 反 映 し て い る と一 応 解 釈 す る 。 産 下 後6時 間 の 卵(Fig. 6, 6h)に はa1 (140kd, pI 5.4)とb (b1, b2, b3を 含 む,100kd, pI 5.7∼5.3), グ ル ー プ の 成 分 と10数 個 の 微 量 成 分 と が 検 出 さ れ た 。 産 下 後1日 の 卵(Fig. 6, 1d)に お い て は6時 間 の 卵 で 検 出 さ れ たa, b, c(矢 印)グ ル ー プ を 構 成 す る 成 分 の 量 が 大 と な っ た 。 さ ら にaグ ル ー プ に はa2成 分 が,b グ ル ー プ に はb4, b5, b6の 成 分 が,Cグ ル ー プ に は c4が そ れ ぞ れ 加 わ り,hグ ル ー プ(h1, h2, h3, h4, 43 kd, pI 6.2∼5.5)とiグ ル ー プ(i1, i2, i3, i4, i5, 25kd, pI 6.4∼5.7)が 新 た に 出 現 し た 。 そ の 他 微 量 成 分 と し てd, e, f, g, jが 検 出 さ れ た 。 産 下 後3日(Fig. 6, 3d)に な る とaとb, c, h, iグ ル ー プ の 成 分 の うち あ る も の は 痕 跡 を と ど め る 程 度 に ま で 減 少 し,ま た あ る も の は 消 失 して し ま っ た 。一 方, 微 量 で は あ る が6時 間 あ る い は1日 の 卵 に は 検 出 さ れ な か っ たA1, A2 (120kd, PI 7.6; pI 7.4)とB (105kd, PI 4.7), C1, C2 (22kd, PI 5.0; pI 4.8)の 成 分 が 出 現 した 。 さ ら に 産 下 後7日 に な る とDとE, F, G, H, I, J, K, L, M, N, O, P, Q, R, S, T, U,の よ う な 多 数 の 成 分 を 包 含 し た グ ル ー プ が 検 出 さ れ る に 至 っ た 。 そ の 中 で も D1, D2 (130kd, pI 5.3∼pI 4.8), E (83kd, pI 5.3∼ 4.5), F (50kd, pI 5.2∼4.8), L (65kd, PI 5.2∼4.8), M (50kd, pI 5.5∼4.7), P (36kd, pI 5.8∼6.2)や11 成 分 か ら な るIグ ル ー プ(50kd),9成 分 か ら な るJグ ル ー プ(45kd),7成 分 か ら な るKグ ル ー プ(39kd),8 成 分 か ら な るRグ ル ー プ(28kd),7成 分 か ら な るSグ ル ー プ(25kd)等 が 主 要 な グ ル ー プ で あ っ た 。 こ の よ うに 分 子 量130kdか ら22kd,等 電 点7.5か ら 4.3に わ た る 広 範 囲 な 分 子 量 な ら び に 等 電 点 を も つ 多 数 の 成 分 が 検 出 さ れ る に 至 っ た の で あ る 。 考 察 カ イ コ 卵 の 形 成 は 主 と し て 蛹 期 の 卵 巣 に お い て 行 わ れ る 。 蛹 期 の 卵 巣 は 包 卵 細 胞 や 栄 養 細 胞 が 体 液 中 か ら種 々 の 物 質 を 取 り込 み,か つ 活 発 に 物 質 の 合 成 を 行 い つ つ 卵 細 胞 内 に 蓄 積 す る(KAWAGUCHI and DOIRA, 1974; TEL-FER, 1975;河 口 ・藤 井,1983)。 そ の 結 果,卵 黄 中 に は タ ン パ ク 質 を 初 め 糖 質 や 脂 質,ア ミ ノ 酸 な ど将 来 胚 子 を 形 成 す る た め に 必 要 な 素 材 と初 期 発 生 に 重 要 な 役 割 を 演 ず る 母 性mRNAやrRNA, tRNAが 具 備 さ れ る よ う に な る。 これ らの素 材 が どの よ うな機 構 で胚 子 形 成 に役 立 つ か を 明 らか にす る 目的 で胚 子 形 成 過 程 に お け る タ ンパ ク質 合 成 の変 動 を放 射 性 ア ミノ酸 の取 り込 み能 で も っ て 比 較 し,さ らに タ ンパ ク質 合 成 に及 ぼす ア クチ ノ マ イ シ ンDの 影 響 につ い て分 析 を試 み た。 胚 子 形 成 過 程 に お け る放 射 性 ア ミノ酸 の取 り込 み か ら み た タ ンパ ク質 合 成 を胚 子 の発 育 階 梯(大 槻,1970)と 対 比 させ てみ る と受 精 完 了後 に起 こる分 割 期(stage 2) か ら胚 帯 形 成 期(stage 3∼4)に か け て の上 昇 とダ ル マ 胚 あ る い は コケ シ形 胚 期(stage 6∼7)を 経 て反 転 期 (stage 11∼15)以 降 に生 じる大 き な上 昇 の二 つが 認 め ら れ る。 これ に対 して,人 工 孵 化処 理 を行 わ な い休 眠 性 卵 で は発 生 初 期 の上 昇 は認 め られ る もの の ダ ル マ形 胚 子 期 (stage 6)以 後 合 成 能 の上 昇 は ま っ た く認 め られ ず,き わ め て低 い水 準 で推 移 す る の み で あ っ た。 す な わ ち 前休 眠 期 に お け る タ ンパ ク質合 成 は非 休 眠 卵 で も休 眠 卵 で も 同 じ程 度 で あ る の に対 し,休 眠期 の胚 子 で は タ ンパ ク質 合 成 能 の増 加 は ほ とん どな い もの と考 え られ る。 一方,ア クチ ノマ イ シ ンDは 受 精 直 後 の 発 生初 期 に お け る タ ンパ ク質合 成 に は ま った く影 響 を お よぼ さな か っ た が,産 下 後3日 か ら5日 に か け て の タ ンパ ク質合 成 に 対 し著 しい 阻害 効 果 を示 した。 しか しな が ら,発 生 後 期 に な る と阻 害 効 果 は低 下 し,タ ンパ ク質 合 成 は ほ とん ど 影 響 され な い。 こ の事 象 をDNA依 存 性 のRNA合 成 を阻 害 す る ア クチ ノ マ イ シ ンDの 作 用 と考 え合 わ せ る な らば,発 生 初 期 に生 じる タ ンパ ク質 合 成 はRNA新 生 を 伴 わ な い 合 成 で あ り,そ れ 以 後 の合 成 は新 生RNAに よ る もの で あ る とい うこ とが で き る。 また,低 濃 度 の ア ク チ ノマ イ シ ンDに お いて も タ ンパ ク質 合 成 の阻 害 効 果 が 認 め られ るが,こ の こ と は卵 の よ うな閉 鎖 系 で あ る ため に 生 じる もの なの か,あ る い は昆 虫 卵 の特 異 性 に よ る も の か 不 明 で あ る の で今 後 の よ り詳 細 な解 析 が 必 要 で あ る。 発 生 初 期 の タ ンパ ク質 合 成 は藤 井 ・河 口(1982)が 指 摘 し た 翻 訳 活 性 能 を も つmRNAとrRNAお よび tRNAを 含 め た 母 性 由来 のRNAを 素 材 に して行 わ れ る もの で あ り,発 生 中期 以 降 の合 成 は受 精 核 由 来 の 新 生 RNAに よ って 行 わ れ る も の にほ か な らな い。 さ らに発 生 後 期 に お け る タ ンパ ク質 合 成 は ア クチ ノ マ イ シ ンDの 影 響 を 受 け る こ とが 少 な い とい う事 象 は,こ の時 期 で は す で にRNA合 成 は 停 止 して い るか あ る いは きわ め て 低 く,旺 盛 な タ ンパ ク質 合 成 は そ れ 以 前 に 作 られ たRNA に 依 存 して 行 わ れ る こ とに よ る と判 断 され る。 この こ と は 人 工 孵 化 処 理 卵 のRNA合 成 は 産 下 後3日 か ら始 ま り
4日 に 最 高 に達 し,6日 か ら7日 に か け て 急 速 に低 下 す る もの で 発 生初 期 と後 期 とに お け る合 成 は き わ め て 低 く,休 眠 卵 で は この よ うなRNA合 成 の 高 ま りは ほ とん どな い とい う倉 田 ・坂 口(1978)の 結 果 と よ く合 致 して い る の で あ る。 オ ー トラジ オ グ ラフ法 を併 用 した2次 元 電 気 泳 動 像 か ら合成 産 物 を解 析す る と,ま ず産 下 後6時 間(6h)の 卵 で はaとb, cの3グ ル ー プが 主 要 産 物 で あ り,1日 (1d)の 卵 に な る とa, b, cグ ル ー プ にhとiグ ル ー プ が加 わ り5グ ル ー プが主 要 産 生 物 とな った 。6hと1d との 間 に はa, b, cグ ル ー プ にみ られ る よ うな産 生 物 に 共 通 性 が あ る こ と,合 成 能 の増 加 と平 行 して 質 的 に も量 的 に も6h<1dの 関 係 が 認 め られ る点 が 特 徴 的 で あ る。 産 下 後3日 の卵 で は前 述 の5グ ル ー プ の主 要 産 生 物 は 減 少 か つ 消 失 して しま う一 方,微 量 で は あ る が ま っ た く 新 しい産 生 物 と してAとB, C成 分 が 出現 した。 さ らに 産 下 後7日 の卵 で はDか らUま で の18グ ル ー プ に 代 表 され る よ うな,き わ め て多 種 類 か つ 多 量 の成 分 が合 成 さ れ る点 に特 徴 が あ る。 産 下 後3日 と7日 の卵 の合 成 産 物 に認 め られ る質 的 な差 異 は3日 の卵 に お け る タ ンパ ク質 合 成 に寄 与 す るmRNA種 と7日 の卵 に お け る タ ンパ ク 質 合 成 に寄 与 す るmRNA種 と の間 に質 的 差 異 の あ る こ とを示 す も の で,こ の こ とは 胚 子 形 態 形 成 に経 時 的 な変 化 が あ る こ とか ら も考 え られ る こ と であ り,そ こ に はあ る種 の調 節 機 構 が 介 在 す る も の と想 像 され る。 以 上 の こ とか ら,母 性mRNAの 情 報 と胚 子 由 来 の mRNAの 情 報 とは 質 的 に大 き な違 いが あ る こ とを 明 示 す る もの で あ り,さ らに胚 子 の発 育 時 期 に よ り特 有 の産 生 物 が あ る こ とは と りもな お さず 胚 子 発 育 過 程 に お け る 時 期 特 異 的 な 遺 伝 子 の 発 現 を 如 実 に 物 語 って い る も の で あ る。 か く して 発 生 初 期 に お け る タ ン パ ク 質 合 成 は 母 性 mRNAに 依 存 す る もの で あ り,発 生 中 期 以 降 に 起 こ る 合 成 は 時 期 特 異 的 な 遺 伝 子 の 活 性 化 に 伴 う胚 子 由 来 の mRNAに よ る もの で あ る とい うこ とを 明示 す る こ と が で きた が,産 生 タ ンパ ク質 の 機 能 的 役 割,分 布 な ど につ い て は 今 後 さ らに 追 求 す る 予 定 で あ る。 摘 要 1) カ イ コ卵 の 胚 子 形 成 過 程 に お け る タ ンパ ク質 合 成 能 の経 時 的 変 化 を放 射 性 ア ミノ酸 の 取 り込 み に よ り調 べ た結 果,発 生 初期(産 下 後1日)と 中 期 以 降(産 下 後 5∼7日)と の 二 つ の 時 期 に 合 成 能 の上 昇 が 認 め ら れ た 。 休 眠 に 入 る卵 で は 発 生 初 期 の 上 昇 のみ が 認 め ら れ る。 2) ア クチ ノ マ イ シ ンD投 与 に よ って も発 生 初 期 の タ ンパ ク質 合 成 は 影 響 を受 け ず 母 性mRNA依 存 性 で あ る が,中 期 以 降 の 合 成 は 著 し く阻 害 され,新 生RNA依 存 性 で あ る と判 断 され る。 3) 胚 子 の 発 育 時 間 に よ り合 成 され る タ ンパ ク質 成 分 は 量 的 に 異 な るの み な らず,質 的 に も異 な って い る こ と を2次 元 電 気 泳 動 オ ー トラ ジ オ グ ラ フに よ り明示 した 。
引 用
文
献
DAVIDOSON, E.H. (1976) Gene activity in early
develop-ment. 2nd ed. New York: Academic Press, p. 452.
藤 井 博 ・河 口 豊(1982)カ イ コ未 受 精 卵 に お け る 母 性 mRNAの 存 在.日 蚕 雑 51: 503-509.
GOLDSTEIN, E.S. and L.A. SNYDER (1973) Protein synthesis
and distribution of ribosomal elements in ovarian
oo-cytes and early developmental stages of Drosophila melano-gaster. Exp. Cell Res. 81: 47-56.
GRZELAK, K., E. SZCZESNA and Z. LASSOTA (1979)
Poly-adenylated RNA in diapausing and developing embryos
of Bombyx mori. Insect Biochem. 9: 125-128.
JACKLE, H. (1979) Degradation of maternal poly
(A)-con-taining RNA during early embryogenesis of an insect
(Smittia spec. Chironomidae, Diptera). Wilhelm Roux's
Arch. Entwicklungsmech. Org. 187: 179-193.
JACKLE, H. and K. KALTHOFF (1979) RNA and protein
synthesis in developing embryos of Smittia spec. (Chiro-nomidae, Diptera). Wilhelm Roux's Arch. Entwicklungs-mech. Org. 187: 283-305.
KAWAGUCHI, Y. and H. DOIRA (1974) Incorporation and
synthesis of protein by the ovaries of Bombyx mori. J.
Fac. Agr., Kyushu Univ. 18: 139-142.
河 口 豊 ・藤 井 博(1983)カ イ コ の退 化 型 小 形 卵 卵 巣 に お け るRNAの 合 成 と蓄 積.日 蚕 雑 52: 233-241.
倉 田 俊 一 ・坂 口文 吾(1978)カ イ コ の非 休 眠 卵 及 び 休 眠 初期 卵 に お け るRNA合 成 能 の変 化.日 蚕 雑 47: 371-374.
LOVETT, J.A. and E.S. GOLDSTEIN (1977) The cytoplasmic
distribution and characterization of poly(A)+RNA in
oocytes and embryos of Drosophila. Dev. Biol. 60: 70-78.
MERMOD, J.J., G. SCHATZ and M. GRIPPA (1980) Specific
control of messenger translation in Drosophila oocytes
and embryos. Dev. Biol. 75: 177-186.
O'FARRELL, P.H. (1975) High resolution two dimensional
electrophoresis of protein. J. Biol. Chem. 250:
4007-4021.
74 河 口 豊 ・藤 井 博
精 編),東 京:三 省 堂, PP. 104-120.
RAFF, R.A. (1980) Masked messenger RNA and the
regu-lation of protein synthesis in egg and embryos. In:
Cell Biology. Vol.4, New York: Academic Press, pp.
107-136.
SCHMIDT, O. and H. JACKLE (1978) RNA synthesis during
oogenesis and early embryogenesis in an insect egg
(Euscolis plebejus). Wilhelm Roux's Arch.
Entwicklungs-mech. Org. 184: 143-153.
SZYSZKO, M. and Z. LASSOTA (1977) Polysomes in
dia-pausing and developing embryos of Bombyx mori. Insect
Biochem. 7: 469-475.
TELFER, W.H. (1975) Development and physiology of the
oocyte-nurse cell syncytium. Adv. Insect Physiol. 11: