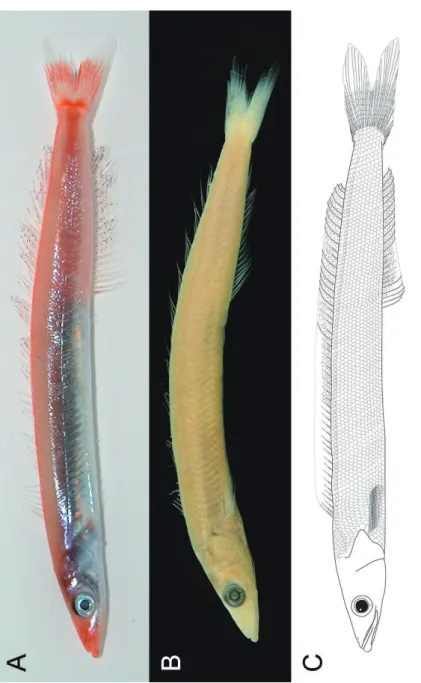
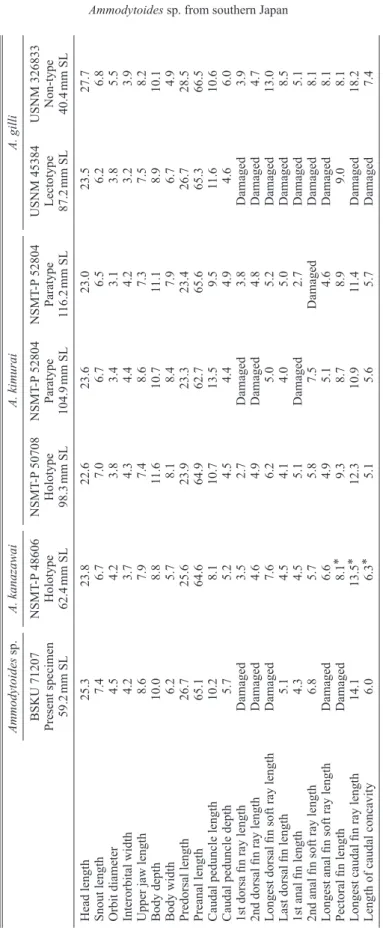
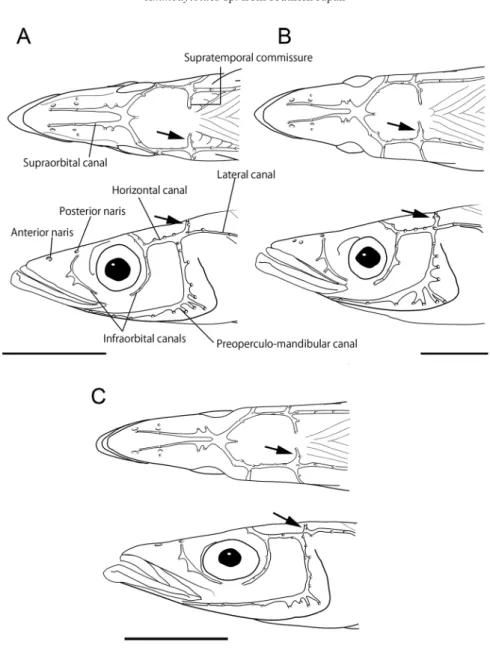
関連したドキュメント
The input specification of the process of generating db schema of one appli- cation system, supported by IIS*Case, is the union of sets of form types of a chosen application system
Laplacian on circle packing fractals invariant with respect to certain Kleinian groups (i.e., discrete groups of M¨ obius transformations on the Riemann sphere C b = C ∪ {∞}),
The only thing left to observe that (−) ∨ is a functor from the ordinary category of cartesian (respectively, cocartesian) fibrations to the ordinary category of cocartesian
Eskandani, “Stability of a mixed additive and cubic functional equation in quasi- Banach spaces,” Journal of Mathematical Analysis and Applications, vol.. Eshaghi Gordji, “Stability
Finally, we give an example to show how the generalized zeta function can be applied to graphs to distinguish non-isomorphic graphs with the same Ihara-Selberg zeta
We use these to show that a segmentation approach to the EIT inverse problem has a unique solution in a suitable space using a fixed point
The main problem upon which most of the geometric topology is based is that of classifying and comparing the various supplementary structures that can be imposed on a
It turns out that the symbol which is defined in a probabilistic way coincides with the analytic (in the sense of pseudo-differential operators) symbol for the class of Feller