666 日 内 分 泌 会 誌,(Folia endocrinol.)67.666∼681.1991
絨 毛 癌 細 胞(BeWo)のhCG(α,β)生
成,
分 泌 制 御 機 構 に お げ る細 胞 内刺 激伝 達 系 の役 割
神戸大学医学部産科婦人科学教室
(指導 望月眞人教授)
西
野
理 一 郎,丸
尾
猛
松
尾
博
哉,望
月
眞
人
The Role of Signal Transduction Systems in the
Regulation of the Production and Secretion of hCG
(ƒ¿,ƒÀ) by Cultured Human Choriocarcinoma Cells (BeWo)
Riichiro NISHINO, Takeshi MARUO,
Hiroya MATSUO and Matsuto MOCHIZUKI
Department of Obstetrics and Gynecology,
Kobe University School of Medicine, Kobe 650, Japan
(Director: Professor Matsuto Mochizuki)
The BeWo cell line, derived from choriocarcinoma, produces and releases human chorionic gonadotropin (hCG) and its ƒ¿-and ƒÀ-subunits. The authors have already reported that cAMP and EGF stimulated the production and secretion of hCG and its subunits by cultured BeWo cells. Therefore, in order to elucidate the role of signal transduction systems (cAMP-A- kinase system, DG - C - kinase system and Ca2+-calmodulin system) in the regulation of hCG (ƒ¿, ƒÀ) synthesis by human choriocarcinoma cells, effects of cholera toxin (CT) , an activator of adenylate cyclase, phorbol 12-myristate 13-acetate (PMA) , a protein kinase C activator, and Ca2+ ionophore A23187, an activator of Ca2+ modulation on hCG (ƒ¿, ƒÀ) production and secretion by BeWo cells cultured in a serum - free condition were evaluated. Immunoreactive hCG ƒ¿, hCG ƒÀ and hCG in the media and cultured cells were measured by each homologous RIA for hCG ƒ¿, hCG ƒÀ and hCG, respectively.
Addition of CT at a concentration of 100ng/ml into the medium caused extreme increases in the cellular levels of hCG ƒ¿ , hCG ƒÀ and hCG together with remarkable increases in hCG ƒ¿, hCG ƒÀ and hCG levels in the medium. This stimulatory effect of CT was first observed on the increase of hCG ƒ¿ levels in cultured BeWo cells and medium at 3h, then observed on the increase of hCG ƒÀ levels at 6h and was last detectable on the increase of hCG levels in the cultured cells and medium at 12h. Addition of PMA at a concentration of 100ng/ml into the medium caused an increase in the cellular and medium levels of hCG ƒ¿,
hCG /3 and hCG shortly (3h) after the exposure to PMA. Addition of A23187 at a concentration of 100ng/ml into the medium caused a slight increase in hCG ƒ¿ levels in the medium at 6h without accompanying the increase in those cellular levels. When added together, PMA potentiated the stimulatory effect of CT on hCG ƒ¿, hCG ƒÀ and hCG levels in the cultured BeWo cells and medium, while PMA did not potentiate the effect of A23187 in this experimental condition.
These findings suggest that cAMP-A-kinase system plays a major role in the signal transduction of hCG (ƒ¿, ƒÀ) synthesis and secretion by BeWo choriocarcinoma cells, and that DG - C - kinase system interacts synergistically with cAMP-A- kinase system in the regulation of hCG (ƒ¿, ƒÀ) synthesis and secretion by BeWo cells. Ca2+-calmodulin system appears to participate in the regulation of hCG ƒ¿ secretion without affecting the synthesis of hCG (ƒ¿, ƒÀ) in BeWo cells.
I緒 言
細 胞 に 対 す る 外 因 性 刺 激 と そ れ に 対 す る 細 胞 応 答 に 関 す る 研 究 は,cyclicAMP(cAMP)の 発 見
以 来 急 速 に 進 展 し,現 在,少 な く と も,細 胞 内 刺 激 伝 達 機 構 と し て,cAMP依 存 性protein kinase
(cAMP-A-kinase系),Ca2+-calmodulin系 な ら び に 西 塚17)18),高 井 ら9)10)に よ り 提 唱 さ れ
たdiacy-lglycerol-C-kinase系(DG-C-kinase系)な ど3種 の 伝 達 系 の 存 在 が 知 ら れ る よ う に な っ た 。
こ れ ら の 系 の う ち,特 にDG-C-kinase系 は,細 胞 外 シ グ ナ ル が 受 容 体 に 結 合 し た あ
とphos-pholipaseCの 作 用 に よ っ てphosphoinositideか らDGが 生 成 さ れ11)24),こ のDGが 生 理 的 濃 度 の
Ca2+の 存 在 下 に 細 胞 膜 リ ン 脂 質(phosphatidylserine)と 協 同 し てC-kinaseを 活 性 化 さ せ16)25),活
性 化 さ れ たC-kinaseの 触 媒 作 用 が 種 々 の 蛋 白 質 を リ ン 酸 化 す る と い う,DGをsecond
messen-gerと し た 新 し い 細 胞 膜 受 容 伝 達 機 構 で あ る 。
い ず れ に し て も,細 胞 外 シ グ ナ ル に 対 す る 細 胞 応 答 は 多 彩 で あ り,C-kinaseやA-kinaseな ら
び にCa2+受 容 蛋 白 質 の 作 用 点 も そ れ ぞ れ の 細 胞 に よ っ て 異 な っ て い る と 考 え ら れ る 。
著 者 ら2)14)はす で に,絨 毛 癌 細 胞 由 来 のce111ineで あ るBeWo細 胞 に お い てcAMPな ら び に
EGF添 加 が,培 養 液 内 な ら び に 培 養 細 胞 内hCG(α,β)レ ベ ル を そ れ ぞ れ 著 明 に 増 加 さ せ,
cAMPで は 添 加 開 始 後 早 期 に,他 方,C-kinase系 を 活 性 化 さ せ る9)EGFの 添 加 で は 添 加 終 了 後 約
24時 間 のlagtimeを 経 て,hCG(α,β)レ ベ ル の 増 加 が 起 こ る と い う 事 象 を 報 告 し た 。
そ こ で 著 者 ら は,絨 毛 癌 細 胞(BeWo)19)に お け るhCG(α,β)生 成 分 泌 制 御 機 構 に 果 た す
cAMP-A-kinase系,Ca2+-calmodulin系 な ら び にDG-C-kinase系 の 役 割 と そ の 相 互 関 係 を 明
ら か に す る 目 的 で,cAMP-A-kinase系 の 促 進 物 質 で あ るcholeratoxin6),Ca2+Ncalmodulin系 の
促 進 物 質 で あ るCa2+ionophore A23187 21)な ら び にDG-C-kinase系 の 促 進 物 質 で あ るphorbol
myristate acetate8)の 同 時 系 統 的 添 加 比 較 実 験 を 行 っ た 。
II材 料 お よ び 方 法
1)実 験 材 料
668 絨毛 癌 細胞(BeWo)のhCG(α,β)生 成,
分泌 制 御機 構 にお け る細 胞 内 刺激 伝 達 系 の役 割(西 野,他3名)
2)培 養 方 法
BeWo細 胞 の 培 養 は,CO2-incubator(AstecCo.,BL-320)内 で37℃,5%CO2-95%airの 条 件
下 に 行 い,培 養 液 と し て は10%fetal bovine serum(FBS),50%Waymouth,40%Gey's
bal-anced salt solutionを 用 い た 。 す な わ ち,ま ず10%FBS含 有 培 養 液 を 用 い て 細 胞 数5万 個/mlに 調
整 さ れ たBeWo細 胞 懸 濁 液 の2mlを12well tissue culture cluster(Costar Corp.)の1 wellご と に
散 布 し て 培 養 を 開 始 し,位 相 差 顕 微 鏡(Nikon Corp.,diaphot TMD)に よ る 観 察 で ほ ぼ50%の
confluencyと な っ た 時 点 で 培 養 液 を10%FBS含 有 培 地 か ら 無 血 清 培 地 へ 交 換 し,cholera toxin
(CT),Ca2+ionophore A23187(A23187)な ら び にphorbol myristate acetate(PMA)を そ れ ぞ れ
24時 間 添 加 し,培 養 液 内 な ら び に 培 養 細 胞 内hCG(α,β)レ ベ ル に 及 ぼ す 影 響 を 検 討 し た 。
BeWo細 胞 培 養 系 に 添 加 さ れ たcholera toxin(Calbiochem Hoechst Co.Ltd., vibrio cholerae
Type Inaba)は1mgを 蒸 留 水1mlで 溶 解 し,membrane filter(Sartorius GmbH., poresize:
0.2μm)に て 濾 過 滅 菌 し た も の を 用 い,他 方,Ca2+ionophore A23187(Sigma Chemical Co.)と
phorbol myristate acetate(Sigma Chemical Co.)は い ず れ も1mgを 共 に99.5%ethanol 1mlで
溶 解 し,同 様 に 濾 過 滅 菌 し た 物 を 用 い た 。
3)培 養 液 内 な ら び に 培 養 細 胞 内hCG(α,β)の 測 定
培 養 液 内immunoreactivehCGα,hCGβ,hCGは,著 者 ら が 抗hCGα 血 清,抗hCGβ 血 清,
抗hCG血 清 を 用 い て 確 立 し た そ れ ぞ れ のhomologousRIA系12)に よ っ て 測 定 し,hCGα,hCGβ,
hCGの 分 泌 レ ベ ル は,す べ てBeWo細 胞1万 個24時 間 あ た り の 分 泌 量 に 換 算 し て 表 現 し た 。
培 養 細 胞 内hCGα,hCGβ,hCGレ ベ ル は,培 養 細 胞 をsonicator(Kontes Co.Ltd.,
microul-trasonic cell disrupter)に て 超 音 波 処 理 し,低 温 遠 心 分 離 器(Hitachi Co.Ltd., 20PR-52D)に て
3000rpm,15分 間 遠 心 分 離 し て 得 ら れ た 上 清 中 のimmunoreactivehCGα,hCGβ,hCGを そ れ
ぞ れ のhomologous RIA系 で 測 定 す る こ と に よ っ て 求 め,す べ てBeWo細 胞1万 個 あ た り の 量 に
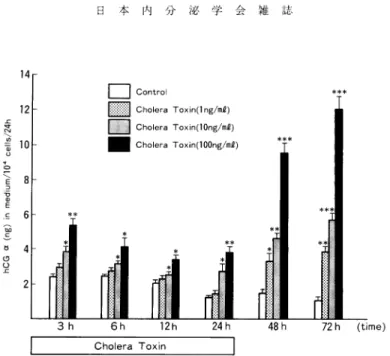
換 算 して 表 現 し た 。 4)統 計 学 的 検 討 各 群 の デ ー タ はmean±SDに て 表 現 し,有 意 差 の 検 定 は,Student'st-testに よ っ て 行 い, P<0.05を 有 意 水 準 と し た 。 III実 験 成 績 1)BeWo細 胞 のhCG(α,β)分 泌 レ ベ ル に 及 ぼ す 影 響 a)choNera toxin(CT)の 影 響 hOGα 分 泌 レ ベ ル はCT添 加 に よ っ て 用 量 依 存 的 に 増 加 し,CT100ng/ml添 加 系 で のhCGα 分 泌 レ ベ ル はCT添 加 開 始 後3時 間 でcontrolに 比 し有 意 に 増 加 し(P<0.01),24時 間 のCT添 加 終 了 か ら24時 間 後 でcontrolの 約6.4倍,48時 間 後 で は 約12倍 に 増 加 し た(Fig.1)。 ま た,hCGβ 分 泌 レ ベ ル もCT添 加 に よ っ て 用 量 依 存 的 に 増 加 し,CT100ng/ml添 加 系 で の hCGβ 分 泌 レ ベ ル はCT添 加 開 始 後6時 間 でcontrolに 比 し有 意 に 増 加 し(P<0.01),24時 間 のCT 添 加 終 了 か ら24時 間 後 でcontrolの 約6倍,48時 間 後 で は 約13倍 に 増 加 し た 。 hCG分 泌 レ ベ ル もCT添 加 に よ っ て 用 量 依 存 的 に 増 加 し,CT100ng/ml添 加 系 で のhCG分 泌 レ ベ ル はCT添 加 開 始 後12時 間 よ り 著 増 し(P<0.01),CT24時 間 添 加 終 了 か ら24時 間 後 でcontrol の 約21倍,48時 間 後 で は 約40倍 に 増 加 し た(Fig.2)。
Fig. 1 Effect of Cholera Toxin on immunoreactive hCG a release into the medium per 104 cells in cultures of BeWo cells. Values are the mean•}SD of triplicate cultures. Asterisks indicate significant differences from control values: *, P<0.05; **, P<0.01; ***, reliable at least to a level of P<0.005.
Fig. 2 Effect of Cholera Toxin on immunoreactive hCG release into the medium per 104 cells in cultures of BeWo cells. Values are the mean•}SD of triplicate cultures. Asterisks indicate significant differences from control values: *, P<0.05; **, P<0.01; ***, reliable at least to a level of P<0.005.
670 絨 毛癌 細胞(BeWo)のhCG(α,β)生 成,
分泌 制 御機 構 にお け る細 胞 内刺 激 伝 達 系 の役 割(西 野,他3名) b)phorbol myristate acetate(PMA)の 影 響
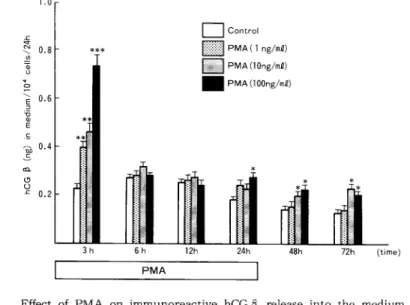
hCGα 分 泌 レベ ル な らび にhCGβ 分 泌 レベ ル はPMA添 加 開 始 後3時 間 で い ず れ も用 量 依 存 的 増 加 を 示 し,PMA100ng/ml添 加 系 で のhCGα 分 泌 レベ ル はcontrolの 約2.4倍,hCGβ 分 泌 レベ ル はcontrolの 約3.2倍 に そ れ ぞ れ 増 加 した が,以 後 のhCGα,hCGβ 分 泌 促 進 効 果 は軽 微 で あ っ た(Fig.3)。
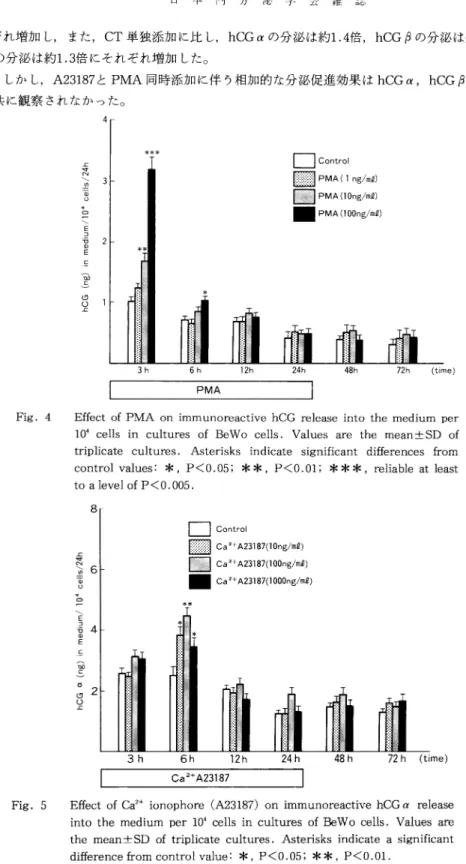
ま た,hCG分 泌 レベ ル はPMA添 加 に よ り,添 加 開 始 後 早 期 に,3時 間 で 用 量 依 存 的 増 加 を 示 し, PMA100ng/ml添 加 系 で のhCG分 泌 レベ ル はcontrolの 約3倍 に増 加 し た(Fig.4)。 こ のPMA添 加 に 伴 うhCG分 泌 レ ベ ル の増 加 はPMA添 加 開 始 後 早 期(3時 間)に 最 も顕 著 で,以 後 のhCG分 泌 促 進 効 果 は 軽 微 で あ っ た 。
c)Ca2+ ionophore A23187(A23187)の 影 響
hCGα 分 泌 レ ベ ル は,A23187 100ng/ml添 加 開 始 後6時 間 の 時 点 で 用 量 依 存 的 増 加 を 示 した が (P<0.01),以 後 はA23187添 加 に 伴 う有 意 な 変 化 を 示 さ な か っ た(Fig.5)。 しか し,hCGな ら び にhCGβ 分 泌 レベ ル はA23187添 加 に 伴 う有 意 な 変 化 を 示 さな か っ た 。
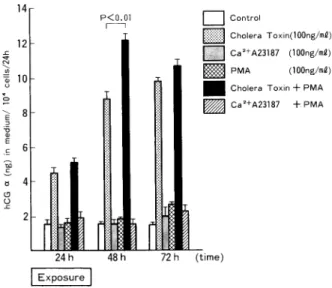
d)cholera toxin(CT),phorbol myristate acetate(PMA),Ca2+ionophore A23187(A23187)の 相 互 作 用 CT100ng/ml添 加 に伴 うhCGα,hCGβ,hCG分 泌 レベ ル の 増 加 は,PMA 100ng/mlを 同 時 に 添 加 す る こ と に よ っ て 一 層 顕 著 と な る こ と を 認 め た 。 す な わ ち,CTとPMA同 時 添 加 に 伴 うhCGα(Fig.6),hCGβ(Fig.7)な らび にhCG分 泌 レ ベ ル の 相 加 的 増 加 は,24時 間 に わ た るCTとPMA同 時 添 加 終 了 か ら24時 間 経 過 後 に最 大 と な り, controlに 比 し,hCGα の 分 泌 は 約7.6倍,hCGβ の 分 泌 は 約10倍,hCGの 分 泌 は 約27倍 へ と そ れ
Fig. 3 Effect of PMA on immunoreactive hCGƒÀ release into the medium per 104 cells in cultures of BeWo cells. Values are the mean•}SD of triplicate cultures. Asterisks indicate significant differences from control values: *, P<0.05; **, P<0.01; ***, reliable at least to a level of P<0.005.
ぞ れ 増 加 し,ま た,CT単 独 添 加 に 比 し,hCGα の 分 泌 は 約1.4倍,hCGβ の 分 泌 は 約1.7倍,hCG の 分 泌 は 約1.3倍 に そ れ ぞ れ 増 加 した 。
し か し,A23187とPMA同 時 添 加 に 伴 う相 加 的 な 分 泌 促 進 効 果 はhCGα,hCGβ な らび にhCG 共 に 観 察 さ れ な か っ た 。
Fig. 4 Effect of PMA on immunoreactive hCG release into the medium per 10 4 cells in cultures of BeWo cells. Values are the mean•}SD of triplicate cultures. Asterisks indicate significant differences from control values: *, P<0.05; * *, P<0.01; ***, reliable at least to a level of P<0.005.
Fig. 5 Effect of Ca2+ ionophore (A23187) on immunoreactive hCGƒ¿ release into the medium per 10•@4 cells in cultures of BeWo cells. Values are the mean •} ESD of triplicate cultures. Asterisks indicate a significant difference from control value: *, P<0.05; ** , P<0.01.
672 絨 毛 癌 細胞(BeWo)のhCG(α,β)生 成,
分泌 制 御機 構 にお け る細 胞 内刺 激 伝 達系 の役 割(西 野,他3名)
Fig. 6 Co-operative effects of Cholera Toxin, PMA and Ca2+ ionophore (A23187) on immunoreactive hCGƒ¿ release into the medium per 104 cells in cultures of BeWo cells. Values are the mean•}SD of triplicate
cultures.
Fig. 7 Co-operative effects of Cholera Toxin, PMA and Ca2+ ionophore (A23187) on immunoreactive hCGƒÀ release into the medium per 104 cells in cultures of BeWo cells. Values are the mean•}SD of triplicate cultures.
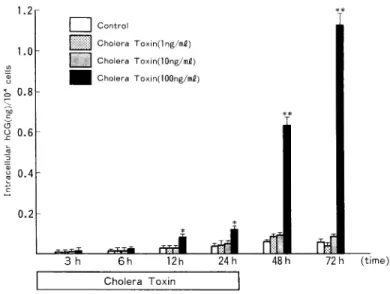
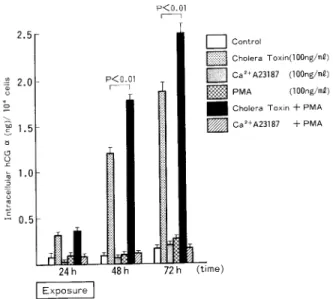
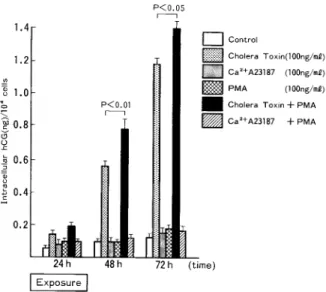
2)BeWo細 胞 の 細 胞 内hCG(α,β)レ ベ ル に 及 ぼ す 影 響 a)cholera toxin(CT)の 影 響 細 胞 内hCGα レ ベ ル は,CT添 加 開 始 後3時 間 よ りCT添 加 量 と 用 量 反 応 的 に 増 加 しは じめ(P< 0.01),CT100ng/ml添 加 系 で は24時 間 に わ た るCT添 加 終 了 か ら24時 間 後 の 時 点 でcontrolの 約 14倍,CT添 加 終 了 か ら48時 間 後 の 時 点 で はcontrolの 約11倍 へ と増 加 し,CT添 加 に 伴 い 細 胞 内 hCGα レベ ル が 著 増 す る こ と を 認 め た(Fig.8)。 他 方,細 胞 内hCGβ レ ベ ル は,CT添 加 開 始 後6時 間 よ り増 加 し(P<0.01),CT100ng/ml添 加 系 で は24時 間 に わ た るCT添 加 終 了 か ら48時 間 後 の 時 点 でcontrolの 約8倍 に 著 増 し た(Fig.9)。 ま た,細 胞 内hCGレ ベ ル は,CT添 加 開 始 後12時 間 よ りCT添 加 量 と 用 量 依 存 的 増 加 を 示 し, CT100ng/ml添 加 系 で は24時 間 に わ た るCT添 加 終 了 か ら24時 間 後 の 時 点 でcontro1の 約11倍, CT添 加 終 了 か ら48時 間 後 の 時 点 で はcontrolの 約20倍 へ と 増 加 し,CT添 加 に 伴 っ て 細 胞 内hCG レベ ル が 著 増 す る こ と を 認 め た(Fig.10)。 b)phorbol myristate acetate(PMA)の 影 響
細 胞 内hCGα,hCGβ な ら び にhCGレ ベ ル はPMA添 加 開 始 後 早 期 に3時 間 で,い ず れ も用 量 依 存 的 増 加 を示 し,PMA100ng/ml添 加 に よ っ て 細 胞 内hCGα レベ ル はcontrolの 約2.3倍,細 胞 内hCGβ レベ ル はcontrolの 約14倍,細 胞 内hCGレ ベ ル はcontrolの 約1.9倍 へ とそ れ ぞ れ 増 加 した(Fig.11)。 こ のPMA添 加 に伴 う細 胞 内hCGα,hCGβ,hCGレ ベ ル の 増 加 はPMA添 加 開 始 後3時 間 に お い て 最 大 で あ り,そ の 促 進 効 果 はPMA添 加 開 始 後6時 間 ま で 軽 度 な が ら観 察 さ れ た
が,そ れ 以 後PMA添 加 に 伴 う有 意 な変 化 は 観 察 され な か っ た 。
Fig. 8 Effect of Cholera Toxin on intracellular immunoreactive hCGƒ¿ levels per 104 cells in cultures of BeWo cells. Values are the mean•}SD of triplicate cultures. Asterisks indicate significant differences from control values: *, P<0.05; **, P<0.01; ***, reliable at least to a level of P<0.005.
674 絨 毛癌 細 胞(BeWo)のhCG(α,β)生 成,
分 泌 制御 機 構 に お け る細 胞 内刺 激 伝 達系 の役 割(西 野,他3名)
Fig. 9 Effect of Cholera Toxin on intracellular immunoreactive hCGƒÀ levels per 10 4 cells in cultures of BeWo cells. Values are the mean•}SD of triplicate cultures. Asterisks indicate significant differences from control values: *, P<0.05; **, P<0.01; ***, reliable at least to a level of P<0.005.
Fig. 10 Effect of Cholera Toxin on intracellular immunoreactive hCG levels per 104 cells in cultures of BeWo cells. Values are the mean•}SD of triplicate cultures. Asterisks indicate significant differences from control values: *, P <0.01; **, reliable at least to a level of P<0.005.
Fig. 11 Effect of PMA on intracellular immunoreactive hCGƒÀ levels per 104 cells in cultures of BeWo cells. Values are the mean •} SD of triplicate cultures. Asterisks indicate significant differences from control values: *, P<0.01; **, reliable at least to a level of P<0.005.
Fig. 12 Co-operative effects of Cholera Toxin, PMA and Ca2+ ionophore (A23187) on intracellular immunoreactive hCGƒ¿ levels per 104 cells in cultures of BeWo cells. Values are the mean•}SD of triplicate cultures.
676 絨 毛 癌細 胞(BeWo)のhCG(α,β)生分 成,
泌 制御 機 構 に お け る細 胞 内刺 激 伝 達系 の役 割(西 野,他3名)
Fig. 13 Co-operative effects of Cholera Toxin
, PMA and Ca2+ ionophore (A23187) on intracellular immunoreactive hCGƒÀ levels per 104 cells in cultures of BeWo cells. Values are the mean•}SD of triplicate cultures.
Fig. 14 Co-operative effects of Cholera Toxin, PMA and Ca2+ ionophore (A23187) on intracellular immunoreactive hCG levels per 104 cells in cultures of BeWo cells. Values are the mean•}SD of triplicate cultures.
c)Ca2+ionophore A23187(A23187)の 影 響
細 胞 内hCGα,hCGβ な らび にhCGレ ベ ル はA23187添 加 に よ る有 意 な 影 響 を 示 さ な か っ た 。 d)cholera toxin(CT),phorbol myristate acetate(PMA),Ca2+ionophore A23187(A23187)の 相 互 作 用 CT 100ng/ml添 加 に 伴 う細 胞 内hCGα,hCGβ な らび にhCGレ ベ ル の増 加 はPMA100ng/ml を 同 時 に 添 加 す る こ と に よ っ て 一 層 顕 著 と な る こ と を 認 め た 。 つ ま り,cTとPMA同 時 添 加 に 伴 う細 胞 内hCGα(Fig.12),hCGβ(Fig.13)な ら び にhCG (Fig.14)レ ベ ル の 相 加 的 増 加 は,24時 間 に わ た るcTとPMA同 時 添 加 終 了 か ら24時 間 経 過 後 に そ の 増 加 は 最 大 と な り,controlに 比 し,細 胞 内hCGα は 約20倍,細 胞 内hCGβ は 約5.3倍,細 胞 内hCGは 約8倍 に 増 加 し,ま た,CT単 独 添 加 に比 し,細 胞 内hCGα は 約1.5倍,細 胞 内hCGβ は 約1.4倍,細 胞 内hCGは 約1.4倍 に増 加 した 。 し か し,A23187とPMA同 時 添 加 に 伴 う細 胞 内hCGα,hCGβ な ら び にhCGレ ベ ル の 有 意 な 変 化 は観 察 され な か っ た 。 IV考 察 著 者 ら は す で に 絨 毛 癌 細 胞(BeWo)培 養 系 で,cAMPとEGFの 添 加 実 験 を 行 いhCG,hCGα, hCGβ の 分 泌 レベ ル な らび に 細 胞 内 レベ ル がcAMPで は 添 加 開 始 後 早 期 に,他 方,EGFで は添 加 終 了 後 約24時 間 の1agtimeを 経 て 増 加 す る こ と を 確 認 し,報 告 した2)。 この こ と よ り,細 胞 内情 報 伝 達 機 構 と して のA-kinase系 及 びC-kinase系 がBeWo細 胞 に お い て そ れ ぞ れ の 刺 激 伝 達 機 構 を 担 う こ と が 推 察 さ れ た 。 そ れ 故 本 研 究 で は,cAMP-A-kinase系 を 賦 活 す るcholera toxin,DG-C-kinase系 を 賦 活 す るPMA,Ca2+uptakeを 促 進 しCa2+-calmodulin系 を 賦 活 す るCa2+iono-phore(A23187)を 用 い て,絨 毛 癌 細 胞 の 腫 瘍 関 連 抗 原 で あ るhCG(α,β)産 生,分 泌 に お け る cAMP-A-kinase系,Ca2+-calmodulin系 な ら び にDG-C-kinase系 の 役 割 と そ の 相 互 作 用 を 検 討 した 。
と こ ろ で,細 胞 内 情 報 伝 達 系 に お い てcAMPとCa2+は 重 要 なsecond messengerで あ る が, cAMPはCa2+,DGを 介 す る受 容 伝 達 機 構 と は 別 種 の 受 容 伝 達 系 を 構 成 して い る 。cAMPを 介 す
る受 容 伝 達 系 は,そ れ 単 独 で 細 胞 機 能 を 調 節 す る が,Ca2+,DGを 介 す る 受 容 伝 達 系 と互 い に 協 調 して 同 一反 応 を 伝 達 す る場 合 や,あ る い は 逆 に互 い に 拮 抗 して 作 動 す る 場 合 が あ る 。 こ の2種 類 の 受 容 伝 達 系 が 互 い に無 関 係 に 作 動 す る か,あ る い は 協 調 的 に 作 動 す る場 合 は,monodirectional control 9)18)と呼 ば れ,逆 に互 い に 拮 抗 的 に 作 動 す る場 合 はbidirectional control 9)18)と呼 ば れ て い る。
今 回 著 者 ら が 行 っ た 実 験 に よ る と,cholera toxin添 加 に よ っ てhCGα,hCGβ,hCGのBeWo
細 胞 内 レベ ル な ら び に 培 養 液 内 レベ ル は い ず れ も増 加 す る こ と が 観 察 さ れ,特 に,細 胞 内 レ ベ ル の増 加 はcholera toxin添 加 開 始 後hCGα が 最 も早 期 に 起 こ り,そ れ に ひ き つ づ い てhCGβ さ ら に は,hCGが 増 加 す る こ と が 認 め られ た 。 この 成 績 は,hCGα とhCGβ のdimerで あ るhCG 7)の BeWo細 胞 内 生 成 に先 立 って,cholera toxinに よ っ て ま ずhCGα 生 成 が,次 い でhCGβ 生 成 が 促 進 さ れ,そ れ に 引 き 続 い てhCG生 成 が 促 進 され る こ と を示 唆 し,cholera toxinに よ って 促 進 され るcAMP-A-kinaseが,絨 毛 癌 細 胞 で のhCGα,hCGβ の 生 成 を そ のmRNA発 現 レベ ル で 促 進 す る可 能 性 が 極 め て 高 い と い え る 。 ま た 著 者 ら の 成 績 で は,choleratoxin添 加 に 伴 うBeWo細 胞 内hCGα,hCGβ,hCGレ ベ ル の 増 加 の 時 間 的 推 移 とhCGα,hCGβ,hCG分 泌 レベ ル の 増 加
678 絨毛 癌 細胞(BeWo)のhCG(α,β)生 成, 分泌 制 御 機構 にお け る細 胞 内 刺激 伝 達 系 の役 割(西 野,他3名) の 時 間 的 推 移 との 問 に差 が み ら れ ず,BeWo細 胞 で のhCG(α,β)生 成 促 進 は た だ ち に そ の 分 泌 促 進 に 連 動 す る こ と が う か が わ れ た 。 著 者 らは す で に トロ ホ ブ ラ ス トで 生 成 さ れ るhCGと そ の subunitは 細 胞 内 に 蓄 積 さ れ る こ と な く分 泌 さ れ る と い う特 性 を有 し,悪 性 トロ ホ ブ ラ ス トで あ る 絨 毛 癌 細 胞 で は そ の 傾 向 が 一 層 顕 著 に な る こ と を 報 告 して い る が13)15)26),今回 認 め たcholera toxin 添 加 に 伴 うhCG(α,β)の 生 成,分 泌 の 連 動 関 係 は 先 の 知 見 に よ く符 号 して お り興 味 深 い 。
と こ ろ で,cholera toxin添 加 に伴 うBeWo細 胞 内cAMP濃 度 はcholera toxin添 加 後 用 量 依 存 的 に増 加 し,cholera toxin(1ng/ml)添 加 後90分 でcontrolの 約300倍,3時 間 で 約900倍 に増 加 す る こ と をRitvosら22)は 報 告 して い る。 こ の こ と は,cholera toxin添 加 が ,BeWo細 胞 内 で 増 加 し たcAMPを 介 してA-kinase系 を 賦 活 し,そ の 結 果 と してBeWo細 胞 で のhCGα ,hCGβ,hCG
の 産 生,分 泌 が 高 ま る こ と を裏 付 け る もの で あ る 。 一 方 ,PMA添 加 に よ っ て,BeWo細 胞 内hCGα,hCGβ,hCGレ ベ ル と そ の 培 養 液 内 レ ベ ル は 共 に 添 加 開 始 後 早 期 に 軽 度 な が ら増 加 す る こ と や,Ca2+ionophore A23187添 加 に よ っ て は, BeWo細 胞 培 養 液 内 のhCGα レ ベ ル が 添 加 開 始 後 早 期 に 軽 度 な が ら増 加 す る こ と を 認 め た 。 つ ま り,PMAに よ っ て 賦 活 され るDG-C-kinaseはBeWo細 胞 で のhCGα,hCGβ,hCGの 生 成 と 分 泌 を 共 に 促 進 す る が,そ の 促 進 のmagnitudeはcAMP-A-kinaseに よ る促 進 のmagnitudeに 比 較 して 小 さ く,ま た,Ca2+ionophore A23187に よ っ て 賦 活 さ れ るCa2+-Ca2+受 容 蛋 白 質 はBeWo 細 胞 に お い てhCGα の 分 泌 レベ ル を 軽 度 増 加 させ る の み で,そ の 細 胞 内 レ ベ ル す な わ ち 生 成 レ ベ ル に は 有 意 な影 響 を 与 え な い こ と が うか が わ れ た 。
さ ら に 著 者 ら はBeWo細 胞 培 養 系 に お い てcholera toxinとPMA ,Ca2+ionophoreとPMAの 同 時 添 加 実 験 を 行 い,hCG(α,β)のBeWo細 胞 内 レベ ル と そ の 分 泌 レベ ル に及 ぼ す 影 響 を検 討 し た 。cholera toxin添 加 に伴 うhCG(α,β)生 成 分 泌 促 進 はPMA同 時 添 加 に よ っ て 一 層 増 強 され る こ と を 認 め た が,Ca2+ionophore A23187とPMAの 同 時 添 加 で は 有 意 な 変 化 を 観 察 しな か っ た 。 以 上 の 成 績 よ り,絨 毛 癌 細 胞BeWoに お け るhCG(α,β)生 成 分 泌 制 御 機 構 は ,cAMP-A-kinase系 が 主 動 的 役 割 を 担 い,Ca2+-calmodulin系 の 関 与 がhCGα 分 泌 に 限 られ,希 薄 で あ る が, DG-C-kinase系 はcAMP-A-kinase系 と協 調 的 に 作 用 して,hCGα ,hCGβ な ら び にhCGの 生 成,分 泌 に対 して 促 進 的 に 働 く こ と が 推 察 さ れ た 。
と こ ろ で,Ritvosら22)はBeWo細 胞 に お い てcholera toxinがhCG分 泌 を 促 進 し ,12-0-tetra-decanoyl phorbol-13-acetate(TPA)もhCG分 泌 をTPA添 加 か ら2時 間 で 約2倍 に 促 進 す る こ と
を,ま た,Benvenisteら8)は 絨 毛 癌 細 胞JEG-3に お い てPMAがhCG分 泌 を 軽 度 促 進 す る こ と を 報 告 して い る。 しか し,こ れ らの 報 告 は い ず れ もhCGの 分 泌 レベ ル に 及 ぼ す 影 響 を論 じた もの で あ り,hCGのsubunitレ ベ ル でcholera toxinな ら び にPMA添 加 の 影 響 を 絨 毛 癌 細 胞 内 レベ ル へ の 作 用 を 含 め て 検 討 した 報 告 は 本 論 文 が は じ め て で あ る。 一 方,Harmanら4)は,trophoblastに
お い てPMAがC-kinaseを 介 してhPL分 泌 を 促 進 す る こ と を,広 田 ら5)は下 垂 体 前 葉 細 胞 に お い てTPAがC-kinaseを 介 してLH分 泌 を 促 進 す る こ と を ,ま たPocotteら20)は 副 腎 髄 質 のchro-maffin細 胞 に お い てPMAがcatecholamineの 分 泌 を 促 進 す る こ と を そ れ ぞ れ 報 告 して い る。 さ
ら に,Ritvosら22)23)はBeWo細 胞 に お い てPMAに よ りcholera toxinのhCG分 泌 促 進 作 用 が 増 強 さ れ る こ と を,ま た,Byusら1)はReuber H35 hepatoma cellに お い てPMAがA-kinase活 性 を 増 強 さ せ る こ と を 報 告 して い る 。
他 方,広 田 ら5)は下 垂 体 前 葉 細 胞 培 養 系 に お い てTPA添 加 群 にCa2+ionophore A23187を 加 え る とLH分 泌 がTPA単 独 添 加 群 の 約2倍 に 増 加 す る こ と を 報 告 し,他 に も,Ca2+-calmodulin系 と DG-C-kinase系 が 協 調 的 に 作 用 す る例 と して,血 小 板 の セ ロ トニ ン放 出 反 応9)や肝 のglycogen分 解3)な ど が 知 られ て い る。 しか し,今 回 の 著 者 らの 成 績 で はBeWo細 胞 で のhCG(α,β)生 成 と 分 泌 レベ ル に及 ぼ すCa2+ionophore A23187とPMAの 協 調 的 促 進 効 果 は 観 察 され な か っ た。 V結 語 hCG(α,β)生 成,分 泌 制 御 機 構 に お け る 細 胞 内 刺 激 伝 達 系 の 役 割 を解 明 す る た め,絨 毛 癌 細 胞(BeWo)培 養 系 に 細 胞 内 刺 激 伝 達
系(cAMP-A-kinase系,DG-C-kinase系,Ca2+-ca-lmodulin系)そ れ ぞ れ の 賦 活 物 質 で あ るcholera toxin,phorbol 12-myristate 13-acetate(PMA), Ca2+ionophore A23187を 添 加 し,培 養 液 内 な ら び に 培 養 細 胞 内hCGα,hCGβ,hCGレ ベ ル に 及 ぼ す 影 響 を 経 時 的 に検 討 し,以 下 の 結 論 を 得 た 。
1)絨 毛 癌 細 胞(BeWo)で は,cholera toxinに よ っ て 賦 活 され るcAMP-A-kinaseが,ま ず 細 胞 内hCGα レベ ル を 著 し く増 加 させ た あ と,細 胞 内hCGβ レベ ル を,次 い で 細 胞 内hCGレ ベ ル を そ れ ぞ れ 大 き く増 加 させ,同 時 に そ の 分 泌 レベ ル を 細 胞 内 レベ ル の 変 化 と 同 一 の 時 間 的 推 移 で 著 明 に促 進 させ る こ と よ り,cAMP-A-kinaseはhCG(α,β)分 泌 機 構 の み な らずhCG(α,β) 生 成 機 構 の 中 で も 主 動 的 役 割 を 担 い,絨 毛 癌 細 胞 で のhCG(α,β)生 成 と 分 泌 は 連 動 関 係 に あ る
こ と が 認 め られ た 。
2)cholera toxin添 加 に 伴 う絨 毛 癌 細 胞(BeWo)内hCG(α,β)レ ベ ル と そ の 分 泌 レベ ル の 増 加 は,DG-C-kinase系 の 賦 活 剤 で あ るPMAの 同 時 添 加 に よ っ て い ず れ も一 層 顕 著 と な る こ と よ
り,絨 毛 癌 細 胞(BeWo)のhCG(α,β)生 成,分 泌 制 御 機 構 の 中 で,DG-C-kinase系 はcAMP -A-kinase系 と協 調 的 に 作 用 し促 進 的 に働 く こ と が 推 察 さ れ た 。
3)Ca2+ionophore A23187ひ こよ っ て 賦 活 さ れ るCa2+-calmodulin系 は 絨 毛 癌 細 胞(BeWo)で の hCGα 分 泌 に促 進 的 に 関 与 す る が,そ の 生 成 制 御 機 構 へ の 関 与 は 希 薄 で あ っ た 。
謝 辞
本 研 究 の 一 部 は 文 部 省 科 学 研 究 費 補 助 金(一 般 研 究C-61570796,C-0267074)に よ り援 助 され た 。
文 献
1)
Byus,
C.V.,
Trevillyan,
J.M.,
Cavit, L. J. and Fletcher,
W.H.:
Activation
of cyclic
adenosine
3' 5' - monophosphate
- dependent
protein
kinase
in H35 hepatoma
and Chinese
hamster
ovary cells by a phorbol ester tumor promoter.
Cancer Res., 43 3321-3326,
1983.
2)
Futamura,
K.,
Maruo,
T. and Mochizuki,
M.: Differential
effects of dibutyryl
cyclic
AMP
and epidermal
growth
factor
on the synthesis
and secretion
of human
chorionic
gonadotropin
and its subunits
by trophoblastic
and non - trophoblastic
cells. Acta Obstet.
Gynecol.
Jpn.,
39: 1641-1648,
1987.
3)
Garrison,
J. C.,
Johnsen,
D.E.
and
Campanile,
C. P.:
Evidence
for the role of phosphorylase
kinase,
protein
kinase
C, and
other
Ca2+- sensitive
protein
kinases
in the response
of hepatocytes
to angiotensin
II and
680 絨 毛癌 細 胞(BeWo)のhCG(α,β)生分 成,
泌制 御 機 構 に おけ る細胞 内刺 激伝 達 系 の 役割(西 野,他3名)
vasopressin.
J. Biol. Chem.,
259: 3283-3292,
1984.
4)
Harman,
I., Zeitler,
P.,
Ganong,
B., Bell, R.M.
and Handwerger,
S.:
Sn-1,2-diacylglycerols
and phorbol esters
stimulate
the synthesis
and release of human
placental
lactogen
from placental
cells: a role
forproteinkinaseC.Endocrinology,119:1239-1244,1986. 5) 広 田 憲 二,青 野 敏 博,
池 上 博 雅,門 脇 浩 三,谷 澤 修:下 垂 体gonadotropin分 泌 とproteinkinaseC.ホ ル モ ソ と 臨 床,
34: 365-369, 1986.
6) Holmgren, J.: Actions of cholera toxin and the prevention
and treatment
of cholera.
Nature,
292: 413-417,
1981.
7) Hussa,
R.O.:
Biosynthesis of human chorionic gonadotropin. Endocrine Rev., 1: 268-294, 1980.
8) Ilekis, J. and Benveniste, R.: Effects of epidermal growth factor, phorbol myristate
acetate,
and arachidonic acid on chorionic gonadotropin
secretion by cultured human
choriocarcinoma cells. Endocrinology, 116: 2400-2409,1985. 9) 川 原 康 洋,津 田
孝,高 井 義 美:diacylglycerol, Ca2+とcyclic AMP.最 新 医 学,40:296-303,1985.
10) Kikkawa, U., Takai, Y., Miyake, R. and Nishizuka, Y.: Protein kinase C as a
possible receptor
protein
of tumor-promoting
phorbol esters. J. Biol. Chem.,
258:
11442-11445, 1983.
11) Kishimoto, A., Takai, Y., Mori, T., Kikkawa, U. and
Nishizuka, Y.:
Activation
of calcium and phospholipid - dependent protein
kinase by
diacylglycerol, its possible relation to phosphatidylinositol
turnover. J. Biol. Chem., 255:
2273-2276, 1980.
12) Maruo, T., Segal, S.T. and Koide, S.: Studies on the
apparent human chorionic gonadotropin-like
factor. Endocrinology, 104: 932-939, 1979.
13) Maruo, T., Matsuo, H., Ohtani, T., Hoshina, M. and Mochizuki, M.: Differential
modulation of chorionic gonadotropin
(CG) subunit mRNA levels and CG secretion by
progesterone in normal placenta and choriocarcinoma
cultured in vitro. Endocrinology,
119: 855-864,
1986.
14) Maruo, T., Matsuo, H., Oishi, T., Hayashi, M.,
Nishino,
R. and Mochizuki,
M.: Induction
of differentiated
trophoblast
function by
epidermal growth factor: Relation of immunohistochemically
detected cellular epidermal
growth
factor levels.
J. Clin.
Endocrinol.
Metab.,
64: 744-750,
1987.
15) Maruo, T., Hoshina, M. and Mochizuki, M.: Differential regulation of chorionic
gonadotropin subunit mRNA levels and secretion by gonadal steroids and growth factors in
normal and malignant trophoblasts.
Bellet D. and Bidart J.M. (ed.) In Structure-Function
Relationship
of Gonadotropins,
Raven Press,
New York, 1989, p.191-208.
16) McCaffrey, P.G., Friedman, B. and Rosner, M.R.
Diacylglycerol modulates binding
and phosphorylation
of the epidermal growth factor receptor.
J. Biol. Chem.,
259:
12502-12507,
1984.
17) Nishizuka, Y.: The role of protein kinase C in cell
surface signal transduction and tumour promotion. Nature, 308: 693-698,
1984.
18) Nishizuka, Y.: Studies and perspectives of protein kinase C. Science, 233: 305-312,
1986.
19) Pattilo, R.A. and Gey, G.O.:
The establishment
of a cell line of
hormone - synthetizing trophoblastic cells in vitro. Cancer Res., 28: 1231-1236, 1968.
20) Pocotte,
S.L.:
Effects of phorbol ester on catecholamine
secretion
and protein
phosphorylation in adrenal medullary cell cultures. Neurobiology, 82: 930-934, 1985.
21) Reed, P.W. and Lardy, H.A.: A23187:A divalent cation ionophore. J. Biol. Chem.,
247: 6970-6977, 1972.
22) Ritvos, O.,
Jalkanen, J., Huhtaniemi, I., Stenman,
U.,
Alfthan,
H. and Ranta,
T.:
12-0- tetradecanoyl
phorbol-13- acetate potentiates
adenosine 3', 5'-monophosphate
- mediated chorionic gonadotropin secretion
by cultured
human choriocarcinoma cells. Endocrinology, 120: 1521-1526, 1987.
23) Ritvos,
O.,
Jalkanen,
J., Pekonen,
F., Stenman,
U. and Ranta,
T.:
Epidermal
growth factor but
not insulin - like growth
factor-I
potentiates
adenosine 3', 5'-monophosphate
- mediated
chorionic gonadotropin secretion by cultured human choriocarcinoma cells. Endocrinology,
123: 859-865, 1988.
24) Sawyer, S.T. and Cohen, S. Enhancement of calcium
uptake and phosphatidylinositol
turnover
by epidermal growth factor in A-431
cells.
Biochemistry, 20: 6280-6286, 1981.
25) TerBush, D.R. and Holz, R.W.: Effects
of phorbol esters, diglyceride, and cholinergic agonists on the subcellular distribution of
protein kinase C in intact or digitonin-permeabilized
adrenal chromaffin cells. J. Biol.
Chem., 261: 17099-17106, 1986.
26) Tojo, S., Mochizuki, M. and Maruo, T.:
Chorionic gonadotropin produced from the cultivated trophoblasts. In: Moudgal N.R. (ed.)
Gonadotropins and Gonadal Function, Academic Press, New York, 1974, p.321-331.
(受 付 日:'90.10.17) (採 択 日:'90.12.26)