つくばリポジトリ SR 7 5545
全文
図
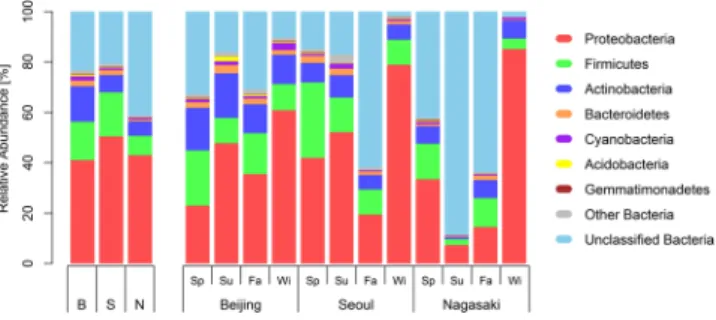

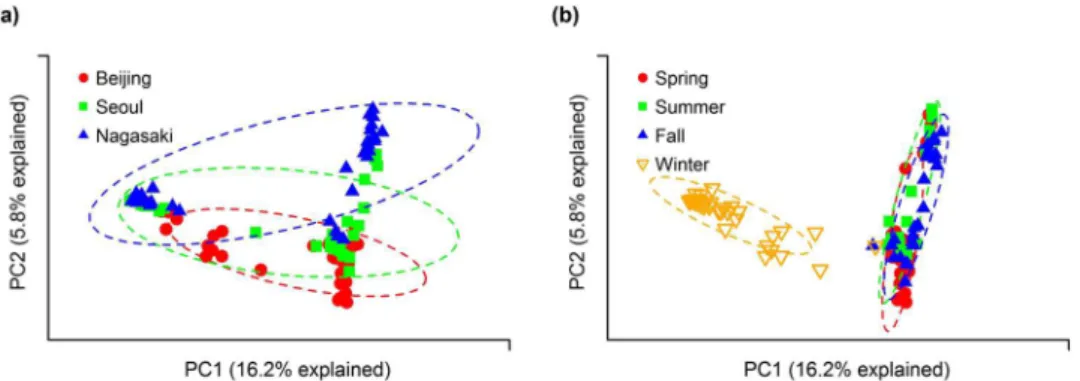
関連したドキュメント
— Holonomic and semi-holonomic geometries modelled on a homogeneous space G/P are introduced as reductions of the holonomic or semi-holonomic frame bundles respectively satisfying
Department of Mathematics, Beijing Jiaotong University, Beijing, P. Several sets of games have been considered earlier to better understand the behaviour of mis`ere games. We
The set of families K that we shall consider includes the family of real or imaginary quadratic fields, that of real biquadratic fields, the full cyclotomic fields, their maximal
Based on the sieving conditions in Theorem 5, together with BTa(n) and BCa(n) that were provided by Boyer, the sieving process is modified herein by applying the concept of
More general problem of evaluation of higher derivatives of Bessel and Macdonald functions of arbitrary order has been solved by Brychkov in [7].. However, much more
) Late 80’s ⇠ : Kesten (’86) anomalous behavior of RW on the critical perco. cluster ) Di↵usions / analysis on fractals (Fractals are “ideal” disordered media).. )
Here we shall supply proofs for the estimates of some relevant arithmetic functions that are well-known in the number field case but not necessarily so in our function field case..
Seventy young mathematicians from developing countries (their names are below) traveled to Beijing, China, with their travel paid by the International Mathematical Union and
