Regulation of the Wnt Signaling Pathways during Cell Culture of Human Mesenchymal Stem Cells for Efficient Bone Regeneration
10
0
0
全文
(2) 38. (MSCs). MSCs can be easily obtained from bone marrow or other sites, and their pluripotent nature allows them to replicate while undifferentiated and then differentiate into lineages of mesenchymal tissue including bone, cartilage, fat and muscle3. In the field of oral and maxillofacial surgery, including dental implant surgery, autogenous bone grafts are considered to be the gold standard for the treatment of bone defects. This well-studied procedure has a good prognosis, but it requires an extra surgical procedure at the donor site, which is an additional burden on the patient. Although allografts, xenografts, and alloplasts are commercially available, these materials tend to be easily infected and induce antigenic responses. Therefore, the establishment of a novel bone regeneration technique is needed to replace the conventional bone graft method. Bone marrow MSCs have great potential for bone regeneration, and clinical applications of MSCs are under way. In previous studies, we used MSCs as isolated cells and platelet-rich plasma (PRP) as the source of growth factors and scaffolds for bone regeneration, and we succeeded in forming bone in the grafted area4. However, the clinical application of growth factors such as BMP-25 and fibroblast growth factor-26 has revealed some problems, including limitations to the amount of bone that can be regenerated. Therefore, new methods for bone regeneration should be investigated. Wnt is a large family of secreted signaling molecules that play a number of important roles during animal development processes7. The canonical Wnt signaling pathway is the Wnt-β-catenin pathway, which regulates cytoplasmic β-catenin, leading to the downstream transcription of Wnt target genes8. Reya et al. first revealed the relationship between stem cells and Wnt signaling; they showed that the canonical Wnt pathway was involved in the self-renewal of haematopoietic stem cells9. Moreover, the canonical Wnt pathway has been implicated in bone formation. Since the osteoporosis-pseudoglioma syndrome has an inactivating mutation in the Wnt co-receptor LRP5 (low-density lipoprotein receptor-related protein 5)10,11, the canonical pathway is considered to be. Oral Science International Vol. 7, No. 2. the key regulator of osteogenesis. In the absence of Wnt signals, β-catenin is phosphorylated by GSK-3β (glycogen synthase kinase-3β) and then degraded by proteosomes. In the presence of Wnt signals, Wnt receptors, which are frizzled homologs, activate the phosphoprotein Dishevelled, and the ability of GSK-3β to phosphorylate β-catenin is then inhibited. Unphosphorylated βcatenin is stable and accumulates in the cytosol and nucleus. In the nucleus, β-catenin binds to T-cell factor (TCF)/lymphocyte enhancer-binding factor (LEF) to form a functional transcription factor, which mediates the transactivation of target genes8. Wnt signals play an important role in the proliferation and differentiation of human MSCs (hMSCs). Wnt 3a, a canonical Wnt member, induces the proliferation of MSCs and β-catenin nuclearization, and inhibits the osteogenic differentiation of MSCs. A low concentration of the Wnt mimic lithium, which inhibits phosphorylation of β-catenin by GSK-3β, also stimulates hMSC proliferation12-14. On the other hand, sFRP-3 (secreted Frizzled-related protein-3), which is a secreted Wnt inhibitor, is upregulated during the osteogenesis of MSCs12. However, these effects of Wnt signaling have not been incorporated into the conventional methods of hMSC cell culture. The regulation of Wnt signals has the potential to accelerate the proliferation and osteogenic differentiation of cultured hMSCs. This acceleration would be clinically beneficial, because it would shorten the treatment time, improve the treatment efficiency, and reduce the running cost of cell culture. In the present study, we investigated the effects of LiCl and sFRP-3 on two aspects of the behavior of hMSCs in vitro, proliferation and differentiation. We hope to establish a more efficient culture process for hMSCs, which would contribute to the future of regenerative medicine of bone. Materials and Methods Cell culture We used commercially available primary marrow-derived hMSCs (Lonza, Walkersville, MD). hMSCs were cultured at 37℃ in a 5% CO2 atmo-.
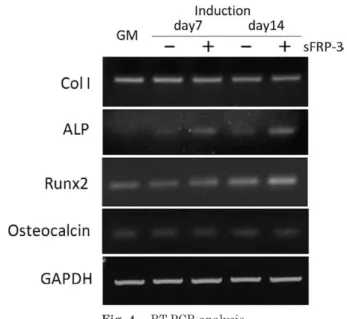
(3) November, 2010. Wnt and MSCs Culture for Efficient Bone Regeneration. sphere. MSCGM, which consists of MSCBM (basal medium) and SingleQuotsⓇ (Mesenchymal Cell Growth Supplement, L-glutamine and penicillin/ streptomycin; Lonza), was used as the growth medium, according to the manufacturer's protocol. We used hMSC Differentiation BulletKit-Osteogenic (Lonza) as the osteogenic induction medium. This medium contains dexamethasone, ascorbate, Mesenchymal Cell Growth Supplement, L-glutamine, penicillin/streptomycin, and β-glycerophosphate. The medium was refreshed every three days. We used a 400 mM stock solution of lithium chloride (LiCl) (Sigma, St. Louis, MO) diluted in MSCGM. Recombinant human sFRP-3 (R&D Systems, Minneapolis, MN) was used at a concentration of 10 ng/ml in the osteogenic induction medium. For growth rate analysis, 30,000 cells were plated onto a 35-mm tissue culture dish. Dishes were then trypsinized on days 3, 6, 9 and 12, and triplicate cell counts were performed visually with the use of a hemacytometer. The proliferation rate of subconfluent cultures was assessed by bromodeoxyuridine (BrdU) incorporation for 24 h using a Zymed BrdU staining kit (Invitrogen, Carlsbad, CA). Immunofluorescent antibody staining Cells grown on cover slips were fixed with PBS containing 4% paraformaldehyde for 15 min and then rendered permeable with PBS containing 0.1% Triton X-100 for 3 min at 4℃. After blocking with 2% bovine serum albuminutes in PBS for 30 min, the cells were incubated with anti-β-catenin mouse monoclonal antibody (BD Biosciences, San Jose, CA) diluted 1 : 500 in blocking solution at 4℃ overnight. Then, the cells were washed and incubated with a 1 : 150 dilution of goat anti-mouse secondary antibody conjugated to rhodamine (Leinco Technologies, St. Louis, MO). To visualize the bound antibody, we washed the cells and incubated them with 60 ng/ml of DAPI (Roche, Indianapolis, IN) diluted in ethanol. Alizarin red staining To detect the calcium depositions of hMSCs, the. 39. induced cells were washed with PBS and fixed with 95% ethanol for 10 min. Then, the cell cultures were washed five times with distilled water and stained with a 2% solution (pH 6.4) of Alizarin Red S (Sigma) for 5 min. PBS was then added for 15 min to reduce the intensity of staining. RNA isolation and reverse transcriptionpolymerase chain reaction (RT-PCR) On days 7 and 14 after osteogenic induction, cells were removed from the culture dish with scrapers, and total RNA was prepared from the cells using an RNeasy Mini Kit (Qiagen, Valencia, CA). To analyze the expression level of mRNA encoding type I collagen, alkaline phosphatase, Runx-2, osteocalcin and GAPDH, total RNA (10 ng) was subjected to RT-PCR analysis with the Titan One Tube RT-PCR System (Roche Molecular Biochemicals, Mannheim, Germany). Specific primers are listed in Table 1. RT-PCR was performed with 25 amplification cycles, according to the manufacturer's protocol. The reaction products were analyzed on a 2% agarose gel and stained with ethidium bromide. Quantitative real-time reverse transcription-polymerase chain reaction (real-time RTPCR) RNA (100 ng) was subjected to reverse transcription using TaqMan EZ RT-PCR Core Reagents (Applied Biosystems, Foster City, CA), according to the manufacturer's instructions. Real-time RTPCR was performed in an ABI PRISM 7000 Sequence Detector System (Applied Biosystems) with the specific primers and probes listed in Table 1. The GAPDH primer and probe (TaqMan GAPDH detection reagents) were purchased from Perkin-Elmer and Applied Biosystems. Cycling conditions were 2 min at 50℃, 30 min at 60℃, 5 min at 95℃ and 50 cycles for 20 sec at 95℃, 1 min at 60℃. Relative mRNA expression levels were normalized to GAPDH expression, and the data from sFRP-3-treated samples were also normalized with the data from the samples without sFRP-3 treatment..
(4) 40. Oral Science International Vol. 7, No. 2 Table 1 Specific primers and probes used in RT-PCR and real-time RT-PCR Primer. Sequence. Genbank ID. Col I. (F)GTGACCGGGGCGAAGCTG (R)ACCGACCTCACCACGTTCAC (Probe) TGGTCCTGCTGGTCCTGCTGGTCC. NM000089. ALP. (F)AGAAAGCCAGGGGCACGAG (R)GGGAGTGCTTGTATCTCGGTTTG (Probe)CCTGGACCTCGTTGACACCTGGAAGAGC. NM000478. Runx2. (F)GCATGTCCCTCGGTATGTCC (R)GGTCCACTCTGGCTTTGGG (Probe)ACCACTCACTACCACAACCTACCTGCCAC. NM004348. OC. (F)AGAGTCCAGCAAAGGTGCAG (R)CCCAGCCATTGATACAGGTAGC (Probe)CTTCACTACCTCGCTGCCCTCCTGCT. NM000711. Col I : Type I collagen, ALP : alkaline phosphatase, OC : osteocalcin. Statistical analysis All experiments were performed at least three times. The paired Student's t test was used to analyze the data, and p < 0.05 was considered to be statistically significant. Results 1. Effect of LiCl and sFRP-3 on hMSC proliferation To investigate the effect of Wnt signaling activation by LiCl on hMSC proliferation, we compared the proliferation of LiCl-treated cells to the proliferation of a control GM group, which was cultured in MSCGM only. We analyzed cell number to assess proliferation and performed BrdU assays to assess DNA synthesis. Compared to the GM group, the hMSCs cultured with 4 mM LiCl had an increased cell number, but the proliferation of the hMSCs cultured with 20 mM or 40 mM LiCl was strongly inhibited (Fig. 1A). Compared to the GM group, the cells cultured with 4 mM LiCl had a statistically significant increase in the number of BrdU-positive cells ; however, culture with 20 mM or 40 mM LiCl yielded a statistically significant decrease in the number of BrdU-positive cells (Fig. 1B). Results from the BrdU assay were consistent with those from the growth rate analysis. We also investigated the effect of sFRP-3, a Wnt inhibitor, on hMSCs proliferation. hMSCs treated. with 10 ng/ml sFRP-3 had a statistically significant reduction in proliferation as compared to the control GM group (Fig. 1A, B). 2. Effect of LiCl on β-catenin localization in hMSCs Immunofluorescent antibody staining against cellular β-catenin was performed to detect the localization of β-catenin in hMSCs (Fig. 2). In the control group, β-catenin was localized to the cytoplasm and rarely found in the nucleus (Fig. 2A-C). In the 4 mM LiCl group, β-catenin was localized to both the cytoplasm and nucleus (Fig. 2D-F); interestingly, intense reactions to β-catenin were seen around the nucleus. In the 40 mM LiCl group, β-catenin was found both in the cytoplasm and nucleus (Fig. 2G-I). The intense reaction around the nucleus seen in the 4 mM group was not prominent, and the nuclear β-catenin was markedly increased in the 40 mM group compared to that in the 4 mM group. 3. Effect of sFRP-3 on the osteogenic differentiation of hMSCs Alizarin red staining was performed on hMSCs that were cultured for 7 or 14 days in osteogenic induction medium with or without 10 ng/ml sFRP-3 (Fig. 3A, B). On day 7, more calcification was seen in the sFRP-3-treated hMSCs than in the hMSCs without sFRP-3. This tendency was.
(5) November, 2010. Wnt and MSCs Culture for Efficient Bone Regeneration. 41. Fig. 1 Effect of LiCl and sFRP-3 on hMSC proliferation.. (A) Growth rate analysis. Proliferation of hMSCs was accelerated with the addition of 4 mM LiCl (4 mM group) compared with the GM group. In the 20 mM and 40 mM groups, hMSC proliferation was strongly inhibited. Addition of sFRP-3 (GM/ sFRP-3 group) suppressed the proliferation of hMSCs. (B) BrdU-positive cells were statistically increased in the 4 mM group but statistically decreased in the 20 mM and 40 mM groups compared with the GM group. In the GM/sFRP-3 group, reduced cell proliferation of hMSCs was shown. Error bars represent S.D. from three experiments. The asterisks indicate significant difference from the control GM group (p < 0.05).. Fig. 2 Immunocytochemistry for β-catenin.. (A-C) hMSCs cultured in basal medium (MSCGM). (D-F) hMSCs cultured with 4 mM LiCl. Cytoplasmic and nuclear β-catenin (arrow). (G-I) hMSCs cultured with 40 mM LiCl. Intense localization of βcatenin in the nucleus (arrow). Scale bar = 200 μm. more prominent on day 14, and the calcification increased rapidly in the sFRP-3-treated hMSCs. Gene expression of type I collagen, alkaline. phosphatase, Runx2 and osteocalcin was also analyzed on days 7 and 14. RT-PCR analysis showed high expression levels of the alkaline phosphatase.
(6) 42. Oral Science International Vol. 7, No. 2. Fig. 3 Alizarin red staining of sFRP-3-treated hMSCs in osteogenic differentiation.. (A) During osteogenic induction of hMSCs, more calcification occurred in the hMSCs treated with sFRP-3 than in the non-treated cells on days 7 and 14. (B) Phase-contrast microscopic view ( × 40).. gene in the sFRP-3-treated hMSCs on days 7 and 14. On days 7 and 14, Type I collagen and Runx2 gene expression was seen regardless of sFRP-3 treatment, while osteocalcin gene expression was relatively weak (Fig. 4). The expression level of alkaline phosphatase mRNA on day 7 was increased in sFRP-3-treated hMSCs compared to hMSCs cultured without sFRP-3 (Fig. 5A). On day 14, the expression level of alkaline phosphatase mRNA tended to be higher in the sFRP-3treated hMSCs compared to the untreated cells (Fig. 5B).. Discussion Bone regeneration with hMSCs will provide minimally invasive techniques, by eliminating the need for surgery at the donor site. Furthermore, this technique does not create any ethical problems, and the patient's safety is guaranteed by the use of their own autogenous cells. These advantages will contribute to increased quality of life for the patient. However, there are problems associated with hMSC-based regenerative medicine. The cultivation of hMSCs is expensive and timeconsuming. Moreover, there are differences in cell.
(7) November, 2010. Wnt and MSCs Culture for Efficient Bone Regeneration. proliferation among patients. Although Wnt signaling is involved in regulating the proliferation and differentiation potential. Fig. 4 RT-PCR analysis. Gene expression of type I collagen (Col I), alkaline phosphatase (ALP), Runx2 and osteocalcin was analyzed on days 7 and 14 after osteogenic induction. RT-PCR analysis showed a high expression level of the ALP gene in the sFRP3-treated hMSCs both on days 7 and 14. There were no obvious differences in the expression levels of the other genes.. 43. of hMSCs12,13, the application of Wnt signaling to optimize culture conditions for hMSCs has not previously been addressed. In the present study, we investigated how to more efficiently cultivate hMSCs for clinical use by utilizing the characteristics of Wnt signaling, which regulates the proliferation of hMSCs and negatively regulates the differentiation of hMSCs. Boland et al. revealed that Wnt 3a exposure inhibits hMSC osteogenic and adipogenic differentiation and promotes cell proliferation12. It was also reported that a high concentration of LiCl promoted the alkaline phosphatase activity and changed the cell appearance, while a low concentration of LiCl decreased the alkaline phosphatase activity and promoted the proliferation of hMSCs13,14. In the present study, we investigated the effect of LiCl on the proliferation of hMSCs by examining their cell number and BrdU incorporation. While the addition of 4 mM LiCl promoted the proliferation of hMSCs compared with hMSCs cultured in MSCGM only, high concentrations of LiCl (20 or 40 mM) inhibited the proliferation of. Fig. 5 Real-time RT-PCR analysis.. (A) The ALP expression level in sFRP-3-treated hMSCs was increased on day 7 compared to that in untreated hMSCs. (B) On day 14, there was a tendency for the ALP expression level to be increased in sFRP-3-treated hMSCs compared to untreated hMSCs. Relative mRNA expression levels were normalized to GAPDH expression. The data from sFRP-3-treated samples were also normalized with the data from the samples without sFRP-3 treatment. Error bars represent the S.D. from three experiments. The asterisks indicate significant differences from the control GM group (p < 0.05)..
(8) 44. hMSCs. Immunocytochemistry against nuclear βcatenin indicated that Wnt signaling was activated in a dose-dependent manner. A high concentration (40 mM) of LiCl caused a strong reaction against nuclear β-catenin but inhibited proliferation. The BrdU assay supports this dose-dependent effect of LiCl and is consistent with the data on population doubling of LiCl-treated hMSCs reported by de Bore et al.13 High concentrations of LiCl inhibit hMSC proliferation and cause cytoskeletal rearrangement. Our preliminary studies also showed the rearrangement of actin filaments and alterations in the shape of hMSCs from spindle to cuboidal, but the dose-dependent inhibition of hMSC proliferation by Wnt 3a conditioned medium has not been reported until now. As the inhibition of GSK3β kinase activity by LiCl is thought to regulate the cytoskeletal Tau protein15,16, de Boer et al. inferred that the dosedependent inhibition of hMSC proliferation by LiCl might occur in a Wnt-independent manner13. In previous studies, Wnt 3a-conditioned culture medium was collected from a murine L cell line that was stably transfected with Wnt 3a and used to activate Wnt signaling in vitro. However, many problems such as safety for using the product of the other animal must be overcome before Wnt 3a can be used clinically. LiCl, however, has been used clinically for decades as an effective drug for bipolar disorder17. When activating Wnt signaling in hMSC culture for clinical use, there are fewer problems associated with LiCl than with Wnt 3a-conditioned culture medium. Moreover, LiCl is not an expensive drug ; thus, it may contribute not only to shortening the culture time but also to reducing the cost of culturing hMSCs. The effects of LiCl on hMSCs are reversible16, and LiCl does not affect the pluripotency of hMSCs13. Our preliminary study also showed that hMSCs treated with 4 mM LiCl could differentiate into an osteogenic lineage (data not shown). Among the clinical applications of osteogenic differentiated hMSCs, the effects of Wnt signaling have not been considered. Wnt signaling inhibits the osteogenic differentiation of hMSCs in vivo12. If we establish a new method of cell culture that reduces the negative effect of Wnt signaling,. Oral Science International Vol. 7, No. 2. the efficient osteogenic induction of hMSCs can be achieved. In the present study, we used the secreted Wnt inhibitor sFRP-3 to downregulate Wnt signaling and investigated how the osteogenic differentiation of hMSCs could be regulated. We found that sFRP-3 suppressed hMSC proliferation, increased early calcification and increased the expression level of the alkaline phosphatase gene 7 days after the initiation of osteogenic induction. On day 14, the calcification of sFRP-3treated hMSCs was more prominent, and the alkaline phosphatase gene expression was more increased compared to that of untreated hMSCs. The promotion of the osteogenic differentiation of hMSCs with sFRP-3 implies that Wnt signaling somewhat inhibits the osteogenic induction under conventional culture methods, but inefficiently. sFRP-3 is a member of the family of soluble proteins that are structurally related to the Wnt receptors, Frizzled proteins. sFRPs are recognized as decoy receptors that bind to Wnt proteins to prevent signal activation18. sFRP-1 causes loss of bone mass in adult mice by inhibiting Wnt-βcatenin signaling in osteoblasts. sFRP-1-/- mice exhibited an increased trabecular bone mineral density, volume and mineral apposition rate compared with sFRP-1+/+ controls19. Oshima et al. reported that human multiple myeloma cell lines constitutively produced sFRP-2 at the protein level and that bone formation was suppressed at least in part through the inhibition of Wnt signaling via the secretion of sFRP-220. Thus, the sFRP families are related to bone formation and act competitively with Wnt signaling. However, contrary to these previous reports, the present study showed that the inhibition of Wnt signaling by sFRP-3 promoted the osteogenic differentiation of hMSCs. There are some differences in the effects of Wnt signaling among different cellular backgrounds and cell origins. For example, Wnt signaling activates alkaline phosphatase in C3H/10T1/2 cells from mouse embryos 21, but Wnt signaling inactivates alkaline phosphatase in hMSCs12,13. Thus, the effects of Wnt signaling on cell proliferation and differentiation are cell-type specific, and little is known about the responsible mechanisms. However, Boland et al. showed that.
(9) November, 2010. Wnt and MSCs Culture for Efficient Bone Regeneration. the gene expression of alkaline phosphatase and bone sialoprotein was downregulated in hMSCs osteogenically induced with Wnt 3a but upregulated in hMSCs osteogenically induced with Wnt 3a and sFRP-312. This result indicates that Wnt signaling is a negative regulator of osteogenic differentiation of hMSCs and that sFRP-3 activates osteogenic differentiation, which is consistent with the results of the present study. Although the effects of Wnt signaling are cell specific, the targets are the same. The targets are the osteogenicrelated genes such as alkaline phosphatase and bone sialoprotein. Gaur et al. demonstrated by mutational analysis that a functional TCF regulatory element responsive to canonical Wnt signaling resides in the prompter of the Runx2 gene; they suggested that the canonical Wnt signaling pathway directly regulated Runx2 and that it was a target of β-catenin/TCF1 for the stimulation of bone formation22. In the present study, the gene expression of alkaline phosphatase was upregulated on day 7, and the gene expression of Runx2 remained relatively high on day 14. However, sFRP-3 treatment did not affect the gene expression of osteocalcin, while more calcification was seen in sFRP-3 treated cells. The discrepant results between calcification and the gene expression of osteocalcin in sFRP-3 treated hMSCs might arise from the stage of calcification. The gene expression of osteocalcin gene is known to be upregulated at the late stage of osteoblast developmental sequence23. Our study focused on the early stage of osteogenic differentiation of hMSCs and the gene expression of osteocalcin gene did not seem to be upregulated at such an early stage. However, the gene expression of Runx2 gene was relatively high on day 14 and Runx2 is known as a positive regulator that can upregulate the expression of osteocalcin24, so the gene expression of osteocalcin is expected to be upregulated at the late stage of osteogenic differentiation of sFRP-3 treated hMSCs. From this point of view, our results indicate that Wnt signaling can affect the early stage of osteogenic differentiation of hMSCs. Careful investigations are required when we consider animal studies, because the effects of Wnt signaling may be cell-type specific. Recently,. 45. it has been reported that Wnt 5a, which is thought to be part of the non-canonical Wnt pathways, regulates cell proliferation and osteogenic differentiation and affects the canonical Wnt pathway25. Potential cross-talk between the canonical and non-canonical Wnt signaling pathways was also reported26. Future studies should clarify the complexities of Wnt signaling in hMSCs. The present study revealed the possibilities of efficient cell culture of hMSCs for regenerative bone medicine by regulating Wnt signaling. In other words, regulation of Wnt signaling promoted cell proliferation without the use of any growth factors and accelerated the osteogenic differentiation of hMSCs. These results will contribute to future regenerative bone medicine by decreasing the time and expense of cell culture and by producing better quality regenerated bone. Acknowledgement This work was supported in part by a Grant-in-Aid for Young Scientists (B) (No. 1979154) from the Ministry of Education, Culture, Sports, Science, and Technology of Japan. Reference 1. Langer R., and Vacanti J.P. : Tissue Engineering. Science 260:920-926, 1993. 2. Takahashi K., Tanabe K., Ohnuki M., Narita M., Ichisaka T., Tomoda K. and Yamanaka S. : Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 131:861-872, 2007. 3. Deans R.J., and Moseley A.B. : Mesenchymal stem cells : biology and potential clinical uses. Exp Hematol. 28:. 875-884, 2000. 4. Yamada Y., Ueda M., Hibi H., and Nagasaka T. : Translational research for injectable tissue-engineered bone regeneration using mesenchymal stem cells and plateletrich plasma : from basic research to clinical case study. Cell Transplant 13:343-355, 2004. 5. Herford A.S., and Boyne P.J. : Reconstruction of mandibular defects with bone morphogenic protein-2 (rhBMP-2). J Oral Maxillofac Surg 66:616-624, 2008. 6. Kitamura M., Nakashima K., Kowashi Y., Fujii T., Shimauchi H., Sasano T., Furuuchi T., Fukuda M., Noguchi T., Shibutani T., Iwayama Y., Takashiba S., Kurihara H., Ninomiya M., Kido J., Nagata T., Hamachi T., Maeda K., Hara Y., Izumi Y., Hirofuji T., Imai E., Omae M., Watanuki M., and Murakami S. : Periodontal tissue regeneration using fibroblast growth factor-2 : randomized controlled.
(10) 46. Oral Science International Vol. 7, No. 2 phase II clinical trial. PLoS ONE 3:e2611, 2008.. 7. Dale T.C. : Signal transduction by Wnt family of ligands. Biochem J 329(Pt2):209-223, 1998.. 17. Freeman M.P., and Freeman S.A. : Lithium : Clinical considerations in internal medicine. Am J Med. 119:478-. 481, 2006.. 8. Akiyama T. : Wnt/β-catenin signaling. Cytokine Growth. 18. Westendorf J.J., Kahler R.A., and Schroeder T.M. : Wnt signaling in osteoblasts and bone diseases. Gene. Factor Rev 11:273-282, 2000. 9. Reya T., Duncan A.W., Ailles L., Domen J., Scherer D.C.,. 341:. 19-39, 2004.. Willert K., Hintz L., Nusse R., and Weissman I.L. : A role. 19. Bodine P.V., Zhao W., Kharode Y.P., Bex F.J., Lambert A.J.,. for Wnt signaling in self-renewal of haematopoietic stem. Goad M.B., Gaur T., Stein G.S., Lian J.B., and Komm B. S. : The Wnt antagonist secreted frizzled-related protein-1. cells. Nature 423:409-414, 2003. 10. Boyden L.M., Mao J., Belsky J., Mitzner L., Farhi A., Mitnick M.A., Wu D., Insigna K., and Lifton R.P. : High bone density due to a mutation in LDL-receptor-related protein. is a negative regulator of trabecular bone formation in adult mice. Mol Endocrinol 18:1222-1237, 2004. 20. Oshima T., Abe M., Asano J., Hara T., Kitazoe K., Sekimoto E., Tanaka Y., Shibata H., Hashimoto T., Ozaki S.,. 5. N Engl J Med 346:1513-1521, 2002. 11. Little L.D., Carulli J.P., del Mastro R.G., Dupuis J.,. Kido S., Inoue D., and Matsumoto T. : Myeloma cells sup-. Osborne M., Folz C., Manning S.P., Swain P.M., Zhao S.C.,. press bone formation by secreting a soluble Wnt inhibitor,. Eustace B., Lappe M.M., Spitzer L., Zweier S., Braunsch-. sFRP-2. Blood 106:3160-3165, 2005.. weiger K., Benchekroun Y., Hu X., Adair R., Chee L.,. 21. Bain G., Müller T., Wang X., and Papkoff J. : Activated. FitzGerald M.G., Tulig C., Caruso A., Tzellas N., Bawa A.,. beta-catenin induces osteoblast differentiation of. Franklin B., McGuire S., Nogues X., Gong G., Allen K.M.,. C3H10T1/2 cells and participates in BMP-2 mediated sig-. Anisowicz A., Morales A.J., Lomedico P.T., Recker S.M.,. nal transduction. Biochem Biophys Res Commun. Van Eerdewegh P., Recker R.R., and Johnson M.L. : A. 84-91, 2003.. 301:. mutation in the LDL receptor-related protein 5 gene. 22. Gaur T., Lengner C.J., Hovhannisyan H., Bhat R.A.,. results in the autosomal dominant high-bone-mass trait.. Bodine P.V., Komm B.S., Javed A., van Wijnen A.J., Stein J.L., Stein G.S., and Lian J.B. : Canonical WNT signaling. Am J Hum Genet 70:11-19, 2002. 12. Boland M., Perkins G., Hall D.J., and Tuan R.S. : Wnt 3a promotes proliferation and suppresses osteogenic differentiation of adult human mesenchymal stem cells. J Cell. promotes osteogenesis by directly stimulating Runx2 gene expression. J Biol Chem 280:33132-33140, 2005. 23. Lian J.B., and Stein G.S. : Concepts of osteoblast growth and differentiation : Basis for modulation of bone cell. Biochem 93:1210-1230, 2004. 13. de Boer J., Wang H.J., and van Blitterswijk C. : Effects of Wnt signaling on proliferation and differentiation of human mesenchymal stem cells. Tissue Eng 10:93-401,. development and tissue formation. Crit Rev Oral Biol Med 3:269-305, 1992. 24. Harada H., Tagashira S., Fujiwara M., Ogawa S., Katsumata T., Yamaguchi A., Komori T., and Nakatsuka M. :. 2004. 14. Etheridge S.L., Spencer G.J., Heath D.J., and Genever. Cbfa1 isoforms exert functional differences in osteoblast. P.G. : Expression profiling and functional analysis of Wnt. differentiation. J Biol Chem 274:6972-6978, 1999.. signaling mechanisms in mesenchymal stem cells. Stem. 25. Mikels A.J., and Nusse R. : Purified Wnt 5a protein activates or inhibits β-catenin-TCF signaling depending on. Cells 22:849-860, 2004. 15. Sperber B.R., Leight S., Goedert M., and Lee V.M. : Glycogen synthase kinase-3β phosphorylates τ protein at multiple sites in intact cells. Neurosci Lett. 197:149-153,. 1995. 16. Stambolic V., Ruel L., and Woodgett J.R. : Lithium inhibits glycogen synthase kinase-3 activity and mimics Wingless signaling in intact cells. Curr Biol 6:1664-1668, 1996.. receptor context. PLoS Biol 4:e115, 2006. 26. Baksh D., Boland G.M., and Tuan R.S. : Cross-talk between Wnt signaling pathways in human mesenchymal stem cells leads to functional antagonism during osteogenic differentiation. J Cell Biochem 2007.. 101:1109-1124,.
(11)
図



関連したドキュメント
The inclusion of the cell shedding mechanism leads to modification of the boundary conditions employed in the model of Ward and King (199910) and it will be
It is suggested by our method that most of the quadratic algebras for all St¨ ackel equivalence classes of 3D second order quantum superintegrable systems on conformally flat
Answering a question of de la Harpe and Bridson in the Kourovka Notebook, we build the explicit embeddings of the additive group of rational numbers Q in a finitely generated group
Next, we prove bounds for the dimensions of p-adic MLV-spaces in Section 3, assuming results in Section 4, and make a conjecture about a special element in the motivic Galois group
Transirico, “Second order elliptic equations in weighted Sobolev spaces on unbounded domains,” Rendiconti della Accademia Nazionale delle Scienze detta dei XL.. Memorie di
Then it follows immediately from a suitable version of “Hensel’s Lemma” [cf., e.g., the argument of [4], Lemma 2.1] that S may be obtained, as the notation suggests, as the m A
In our previous paper [Ban1], we explicitly calculated the p-adic polylogarithm sheaf on the projective line minus three points, and calculated its specializa- tions to the d-th
Our method of proof can also be used to recover the rational homotopy of L K(2) S 0 as well as the chromatic splitting conjecture at primes p > 3 [16]; we only need to use the
