博 士 学 位 論 文
ヒト型可溶性トロンボモジュリンの HMGB1 誘起アロディニア
に対する抑制作用の分子メカニズムに関する研究
平 成 3 0 年 3 月近 畿 大 学 大 学 院
薬 学 研 究 科 薬 科 学 専 攻
辻
田
隆
一
目次
緒論 ... 1 第 1 章 ヒト型可溶性トロンボモジュリンの HMGB1 誘起アロディニア抑制効果発現 におけるトロンビンの役割 ... 3 緒言 ... 3 I. 実験方法 ... 5 II. 実験結果 ... 8 III. 考察 ... 16 第 2 章 ヒト型可溶性トロンボモジュリンの HMGB1 誘起アロディニア抑制効果発現 における lectin 様ドメイン及び EGF 様ドメインの必要性 ... 18 緒言 ... 18 I. 実験方法 ... 19 II. 実験結果 ... 21 III. 考察 ... 27 総括 ... 29 引用文献 ... 31 略語一覧 ... 35 主論文 ... 36 謝辞1
緒論
痛みは、生体にとって外界からの侵害刺激や傷害を受けた組織部位を知る警戒信号 として重要である。末梢組織傷害や炎症による疼痛が過剰になった場合は、 Non-Steroidal Anti-Inflammatory Drugs(NSAIDs)や麻薬性鎮痛薬が奏功する。一方、疼 痛の中でも、神経系の傷害より慢性的な痛みを起こす末梢神経障害性疼痛は、警戒信 号としての意義は薄く、患者の quality of life(QOL)を著しく損なうほか1)、それに伴 う社会的な経済的損失も大きい。日本では、古来より「痛み」を耐えることが美徳と されてきたが、近年は痛みを積極的にコントロールする緩和療法の重要性が広く認知 されている。末梢神経障害性疼痛は、NSAIDs や麻薬性鎮痛薬による治療に抵抗性を 示すことが多いため、抗うつ薬や抗てんかん薬などが治療に応用されているほか、電 位依存性カルシウムチャネルのα2δ サブユニットに結合するプレガバリンなどが使用 されている。しかし、既存の治療薬では、十分な効果が得られないことや重篤な副作 用が見られることも多く、新たな標的分子に作用する新薬の開発が望まれている。
このよう状況の中、米国において 2001 年に「the Decade of Pain Control and Research」 が宣言され 2)、様々なタイプの痛み受容体や疼痛伝達に関与するチャネルをターゲッ
トにした新薬の開発が加速化している。さらに、敗血症の原因となる病原体由来分子 の pathogen-associated molecular patterns(PAMPs)や、生体由来の damage-associated molecular patterns(DAMPs)が3)、樹状細胞や上皮細胞あるいは免疫細胞に発現してい る pattern recognition receptors(PRRs)を介し、炎症やアポトーシスのみならず、「痛 み」の発現に深く関与することから4-6)、新しい創薬ターゲットとして注目されている 7-13)。
DAMPs の一つである核内蛋白質の high mobility group box 1(HMGB1)は、215 残 基のアミノ酸より構成された分子量約 30 kDa の非ヒストン DNA 結合蛋白質で、核中 において DNA の安定化に関与しているほか、p53 や NF-κB などの転写因子を制御し ている。HMGB1 のアミノ酸配列は、哺乳類間で高く保たれており約 99%の相同性が ある5)。その分子内には 3 つのシステイン残基(C23、C45 及び C106)を有しており、
2
する。一方、病原性細菌や物理的な傷害により核外に放出された HMGB1 は、周囲の 環境により還元型の at-HMGB1、C23 と C45 がジスルフィド結合した disulfide-HMGB1 (ds-HMGB1)、及び全てのシステインが酸化型の oxidized-HMGB1(ox-HMGB1)の 3 形態をとる。細胞外の at-HMGB1 は主に receptor for advanced glycation endproducts (RAGE)を、また ds-HMGB1 は主に Toll-like receptor 4(TLR4)を活性化すること で、局所炎症刺激による組織修復や組織因子の発現誘導による止血機構の活性化に寄 与するほか14)、炎症反応や痛みの情報伝達を促進する4-6)。尚、ox-HMGB1 の生理活 性は見出されていない15)。そこで、本研究では特に HMGB1 不活性化作用を有するこ とが知られているヒト型可溶性トロンボモジュリン(TMα)を用いる疼痛治療に焦 点をあて、HMGB1 依存性疼痛に対する抑制効果の分子メカニズムを解析した。 第 1 章では、立体構造の異なる 2 つの HMGB1、すなわち at-HMGB1 と ds-HMGB1 のトロンビンによる分解に対して、TMαがどのように影響するかを in vitro で検証し、 さらに、マウスを用いて HMGB1 依存性の機械的アロディニアに対する TMαの抑制 作用にトロンビンが関与するか否かを検討した。第 2 章では、トロンボモジュリンの 5 つのドメイン(D1~D5)のうち、TMαを構成する D1~D3 ドメインの役割を解析 した。すなわち、ピキア酵母により、D1、D2 及び D123 を作製し、at-HMGB1 と ds-HMGB1 のトロンビンによる分解に対する影響を TMαと比較検討し、さらに、in vivo 試験において、HMGB1 依存性アロディニアに対する効果を評価した。
3
第 1 章 ヒト型可溶性トロンボモジュリンの HMGB1 誘起
アロディニア抑制効果発現におけるトロンビンの役割
緒言
核内蛋白質の一つである HMGB1 は、核内において DNA の安定化や転写調節を担 う重要な働きを有する。一方、細菌感染や外傷などにより細胞外に放出された HMGB1 は、DAMPs(damage-associated molecular pattern molecules)として自然免疫、獲得免疫、 または炎症反応の誘導に関与している3)。HMGB1 は、核から細胞質へ移行あるいは細胞外へ放出された後、周囲の酸化還元状態に依存して at-HMGB1 や ds-HMGB1 の構 造を取る14)。前者は RAGE(receptor for advanced glycation endproducts)を、後者は TLR
(Toll-like receptor)4 を主に介して炎症反応を促進する。興味あることに、マウスや ラットにおいて、at-HMGB1 や ds-HMGB1 の足底内投与によって機械的アロディニア が誘起される6,9)ほか、cyclophosphamide 誘起膀胱炎モデル16)や cerulein 誘起急性膵炎 モデル17)における内臓痛の発症に内因性のHMGB1 が関与することが報告されている 6)。 血管内皮細胞上に発現している糖蛋白質のトロンボモジュリンは、D1~D5 の 5 つ のドメインからなり、D2 に結合したトロンビンが血液中の protein C を activated protein C(APC)に変換し、補酵素の protein S の働きを受け凝固系カスケードにネガティブ フィードバックをかけることで抗凝固作用を示す 18)。最近、トロンボモジュリンが、 D1 で HMGB1 を吸着し、D2 に結合したトロンビンによる分解を促進することが報告 されている19)。トロンボモジュリンの D1~D3 からなる遺伝子組換え蛋白質を CHO 細胞で作製したヒト型可溶性トロンボモジュリン(TMα)は、トロンボモジュリン の生理活性の多くを保持しており、日本では汎発性血管内凝固症(DIC:disseminated intravascular coagulation)治療薬として臨床応用されている。そこで本章では、TMαが、 at-HMGB1 及び ds-HMGB1 のトロンビンによる分解に影響を及ぼすかを in vitro で調 べ、さらに、マウスにおいて at-HMGB1 あるいは ds-HMGB1 を足底内投与すること で誘起されるアロディニアと、内因性 HMGB1 が関与するリポポリサッカライド
4
(LPS:lipopolysaccharide)の足底内投与によるアロディニア9)に対し、TMαの抑制作
5
I. 実験方法
全ての動物実験は、近畿大学実験動物委員会により承認され、米国国立衛生研究所 による実験動物の管理と使用のためのガイドラインに従って実施した。実験動物は、 3-4 週齢のddY系雄性マウス(紀和実験動物、和歌山)を使用した。マウスは室温22-24℃、 12 時間毎の明暗サイクル(7 時から 19 時まで点灯)下で飼育し、実験の開始まで飼料 (MF、オリエンタル酵母)及び水を自由に摂取させた。 実験に使用した試薬は以下のとおりである。 Bovine thymus-derived HMGB1(bt-HMGB1)は、シノテスト(東京、日本)から購 入した。遺伝子組換え all-thiol HMGB1(at-HMGB1)及び disulfide HMGB1 (ds-HMGB1) は、HMGB1Biotech(Milan, Italy)から購入した。Thrombomodulin alfa[TMα; Chinese hamster ovary(CHO)細胞で作製された遺伝子組換えヒト型可溶性トロンボモジュリ ン]は、旭化成ファーマ株式会社(東京、日本)から提供された。ヒト・トロンビン、 アルガトロバン(AT)及びリポポリサッカライド(LPS; Escherichia coli Serotype O26:B6) は、Sigma-Aldrich(St. Louis, MO, USA)から入手した。bt-HMGB1、at-HMGB1、 ds-HMGB1 及び LPS は、生理食塩水に溶解し、10 μL の容量でマウスの足底内に投与 した。TMαは、0.002% Tween 80 を含む生理食塩水に溶解し、各 HMGB1 または LPS 投与の 1 時間前に 10 μL の容量でマウスの足底内に投与した。AT は、TMα投与の 30 分前に腹腔内投与した。トロンビンは、1 U/paw の用量で TMαと同時に足底内に投与 した。 3. HMGB1 及び LPS 誘起マウスアロディニアモデルの作製 HMGB1 及びLPS 誘起マウスアロディニアモデルの作製は、既報に従い実施した6,9)。 すなわち、マウスの足底内に 100 ng/paw の bt-HMGB1、100 ng/paw の at-HMGB1、10 ng/paw の ds-HMGB1 または 100 ng/paw の LPS を投与することにより機械刺激に対す るアロディニアを発症させた。1. 実験動物
6
4. von Frey filaments によるアロディニアの評価
マウスを実験台から約 25cm 浮かせたメッシュの上に置き、透明のプラスチックボ ックス(10 cm×10 cm×10 cm)を被せ、8 種類のフィラメント(0.008, 0.02, 0.04, 0.07, 0.16, 0.4, 0.6 及び 1.0g)で右足底部皮膚表面を刺激した。フィラメントでの刺激は、強 度の弱いものから順に行い、up-down 法20)に従って侵害受容閾値を測定した。各フィ ラメントでの刺激は、5-10 秒以上の間隔をあけて 10 回行った。’wind up’ 現象や脱感 作を防ぐため、同一部位への繰返し刺激は避けた。評価ポイントは、Pre 値、HMGB1 または LPS 投与 0.5, 1, 1.5, 2, 3, 4, 5 時間後とした。さらに、HMGB1 または LPS 投与後 1.5 から 3 時間の time-threshold 曲線より AUC を算出し被験薬の効果を評価した。 5. Western blot 法 200~400 nM の at-HMGB1 または ds-HMGB1 と、2 U/mL のトロンビンを含む 40 μL の反応溶液(2 mM CaCl2、0.1 M NaCl、50 mM Tris-HCl、pH8.0)を、400 nM の TMα
存在下または非存在下、37℃で 15 分または 2 時間インキュベートした。反応終了後、 2%の sodium dodecyl surfate(SDS)、10% glycerol、2-mercatoethanol 及び bromophenol blue を加え、95-100℃で 5 分間煮沸処理した。サンプル中の蛋白質は 12.5%の SDS-polyacrylamide gel(和光純薬工業、大阪)を用いて 2 時間の電気泳動により分離 し、分離させた蛋白質を polyvinylidene difluoride(PVDF)膜 (Immobilon-P; Millipore Corporation, Billerica, MA, USA)へ転写し、blocking buffer(5% skim milk、137 mM NaCl、 0.1% Tween 20、20 mM Tris-HCl、pH 7.6)で処理した後、HMGB1 の C 末を認識する anti-HMGB1 polyclonal rabbit antibody(Abcam, Cambridge, UK)を 10,000 倍に希釈して 4℃で一晩反応させた。洗浄後、二次抗体として HRP-conjugated anti-rabbit IgG 抗(Cell Signaling Technology, Danver, MA, USA)を blocking buffered で 1,000 倍希釈し室温で 90 分間反応させ chemiluminescence(ナカライテスク、京都)で発光させ、Image Quant 400 (GE healthcare, Buckinghamshire, UK)でバンドを検出した。これをパーソナルコンピ ューターに取り込んだ後、ImageJ を用いてバンドの濃さを数値化した。
7
6. 統計処理
得られた実験値は、平均値 ± 標準誤差で示した。統計学的有意性の判定には、正 規分布は Tukey’s 検定、非正規分布のデータは Kruskal-Wallis H 検定を用い、P < 0.05 で有意差ありとした。
8
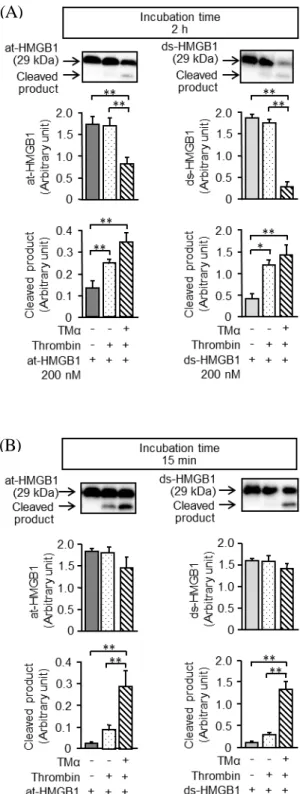
II. 実験結果
Fig. 1A は、200 nM の at-HMGB1 または ds-HMGB1 に 2 U/mL のトロンビンを加え、 37℃、2 時間のインキュベ―ション後に Western blot を実施した結果である。29 kDa に 認められる at-HMGB1 及び ds-HMGB1 のバンドは、トロンビン単独では減少しなかっ たが、400 nM の TMα存在下ではトロンビンによって at-HMB1 及び ds-HMGB1 のバ ンドが有意に減少した。一方、at-HMB1 及び ds-HMGB1 の cleaved product のバンドは、 トロンビンにより有意に増加し、TMα存在下ではこの増加効果がさらに増強された。 Fig. 1B は、400 nM の at-HMGB1 または ds-HMGB1 に 2 U/mL のトロンビンを加え、 37℃、15 分間のインキュベ―ション後に Western blot を実施した結果である。 at-HMGB1 及び ds-HMGB1 のバンドは、いずれの条件下でも明らかには減少しなかっ たが、cleaved product のバンドは、TMα存在下、トロンビンによって有意に増加した。 1. トロンビンによる HMGB1 分解に対するTMα の促進効果
9
Fig. 1 Effects of TMα on thrombin-induced cleavage of at-HMGB1 or ds-HMGB1. In the
absence or presence of TMα at 400 nM, at-HMGB1 or ds-HMGB1 at 200 nM (A) or 400 nM (B) was incubated with thrombin at 2 U/mL for 2 h (A) or 15 min (B). The HMGB1 in distinct redox forms and its cleaved product were analyzed by Western blotting. The top panels show typical blotting photographs, and the protein levels were quantified by densitometry. Data show the mean ± S.E.M. from 5 distinct experiments. *P<0.05, **P<0.01.
(A)
10
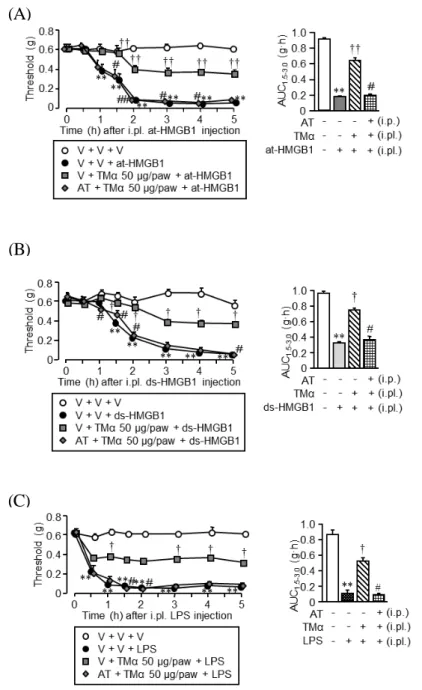
Fig. 2 は、ウシ胸腺から精製した bt-HMGB1(100 ng/paw)、遺伝子組換えにより作 製した at-HMGB(100 ng/paw)及び ds-HMGB1(10 ng/paw)、ならびに LPS(100 ng/paw) をマウスの足底内に投与することで誘起したアロディニアに対する TMαの抑制効果 を調べた結果である。各刺激剤で誘起された閾値低下は、いずれも 5 時間以上継続し ていたが、LPS は、3 種類の HMGB1 に比べて、より早期にアロディニアを誘起する ことが判明した。各種 HMGB1 あるいは LPS 投与 1 時間前に TMαを 0.5~50 μg/paw の用量で足底内投与したところ、用量依存的な抗アロディニア効果が認められた。し かしながら、TMαは 50 μg/paw でも各アロディニアを完全には抑制することができな かった。 2. 各種 HMGB1 及び LPS 誘起アロディニアに対する TMαの用量依存的な抑制効果
11
Fig. 2 Dose-dependent effects of i.pl. administration of TMα on the mechanical allodynia
induced by i.pl. injection of bovine thymus-derived HMGB1 (bt-HMGB1), at-HMGB1, ds-HMGB1 or LPS in mice. TMα at 0.5, 5 or 50 μg/paw or vehicle was administered i.pl. 1 h before i.pl. injection of bt-HMGB1 at 100 ng/paw, at-HMGB1 at 100 ng/paw, ds-HMGB1 at 10 ng/paw, LPS at 100 ng/paw or vehicle. Left panels show the time course of the nociceptive threshold, and right panels indicate the AUC between 1.5 and 3 h after i.pl. bt-HMGB1, at-HMGB1, ds-HMGB1 or LPS. Data show the mean ± S.E.M. for 5-6 mice. V, vehicle. **P<0.01 vs. V + V; †P<0.05, ††P<0.01 vs. V + bt-HMGB1, at-HMGB1, ds-HMGB1 or LPS.
(A)
(B)
(C)
12
3. HMGB1 誘起アロディニアに対する TMαの抑制効果と内在性トロンビンの役割
Fig. 3 は、at-HMGB1、ds-HMGB1 または LPS により誘起されるアロディニアに対す る TMαの抑制効果に内因性トロンビンが関与するか否かを調べた結果である。100 ng/paw の at-HMGB1、10 ng/paw の ds-HMGB1 及び 100 ng/paw の LPS によるアロディ ニア誘起は、50 μg/paw の TMαにより部分的にではあるが有意に抑制された。この TMαの抗アロディニア作用は、トロンビン直接阻害薬の AT 10 mg/kg(i.p.)を前投与 することで完全に消失した。
13
Fig. 3 Argatroban (AT), a thrombin inhibitor, abolishes the suppressive effect of i.pl. TMα on
the mechanical allodynia induced by i.pl. injection of at-HMGB1, ds-HMGB1 or LPS in mice. TMα at 50 μg/paw or vehicle was administered i.pl. 1 h before i.pl. injection of at-HMGB1 at 100 ng/paw, ds-HMGB1 at 10 ng/paw or LPS at 100 ng/paw. AT at 10 mg/kg or vehicle was administered i.p. 0.5 h before i.pl. TMα. Left panels show the time course of the nociceptive threshold, and right panels indicate the AUC between 1.5 and 3 h after i.pl. bt-HMGB1, at-HMGB1, ds-HMGB1 or LPS. Data show the mean ± S.E.M. for 5-8 mice. V, vehicle. **P<0.01 vs. V + V + V; †P<0.05, ††P<0.01 vs. V + V + at-HMGB1, ds-HMGB1 or LPS; #P<0.05 vs. V + TMα + at-HMGB1, ds-HMGB1 or LPS.
(A)
(B)
14 4. HMGB1 誘起アロディニアに対する TMαの抑制効果とトロンビンの併用効果 Fig. 4 は、at-HMGB1、ds-HMGB1 及び LPS 誘起アロディニアに対する TMαの部分 的な抑制効果が、トロンビンの局所投与により影響するか否かを調べた結果である。 先と同様に、at-HMGB1、ds-HMGB1 及び LPS 誘起アロディニアに対し、50 μg/paw の TMαは、部分的な抗アロディニア効果を示したが、少量のトロンビン(1 U/paw)を 足底内に前投与すると、この抗アロディニア効果は著しく増強され、アロディニアが ほぼ完全に消失した。
15
Fig. 4 Thrombin enhances the suppressive effect of i.pl. TMα on the mechanical allodynia
induced by i.pl. injection of at-HMGB1, ds-HMGB1 or LPS in mice. TMα at 50 μg/paw and thrombin at 1 U/paw was administered i.pl. 1 h before i.pl. at-HMGB1 at 100 μg/paw, ds-HMGB1 at 10 μg/paw or LPS at 100 ng/paw. Left panels show the time course of the nociceptive threshold, and right panels indicate the AUC between 1.5 and 3 h after i.pl. bt-HMGB1, at-HMGB1, ds-HMGB1 or LPS. Data show the mean ± S.E.M. for 9-11 (A, C) or 5 (B) mice. V, vehicle. *P<0.05, **P<0.01 vs. V + V + V; †P<0.05, ††P<0.01 vs. V + V + at-HMGB1, ds-HMGB1 or LPS; #P<0.05 vs. V + TMα + at-HMGB1, ds-HMGB1 or LPS.
(A)
(B)
16
III. 考察
本章では、初めに、TMαがトロンビンによる at-HMGB1 及び ds-HMGB1 の分解を いずれも促進することを実証した。トロンビンは、HMGB1 の N 末から 10 番目の Arginine と 11 番目の Glycine の間を切断することが報告されているが19)、今回の検討 により、N 末端周辺の立体構造が異なる at-HMGB1 と ds-HMGB1 が何れもトロンビン による切断を受け、TMα存在下では、この反応がより効率的に進むことが明らかと なった。 at-HMGB1、ds-HMGB1 及び LPS 誘起アロディニアに対して、TMαは用量依存的な 抑制効果を示し、50 μg/paw の TMαの効果は 1 U/paw のトロンビンの併用投与により 増強された。In vitro では高濃度のトロンビン単独で HMGB1 を分解することができる が、in vivo では今回用いた用量のトロンビン単独では HMGB1 誘起アロディニアを抑 制しなかった。これは、足底皮下局所では内因性トロンボモジュリンの発現量が十分 ではないか、内因性アンチトロンビンによってトロンビンが失活したためではないか と考えられる。HMGB1 は TMαの D1(lectin-like domain)に結合し、D2(EGF-like domain) に結合したトロンビンにより効率的に分解されると推察されるが、D1 による HMGB1 の吸着のみによっても HMGB1 の作用を抑制できる可能性も考えられる。しかしなが ら、TMαの抗アロディニア効果がトロンビン直接阻害薬の AT により完全に消失した ことから、TMαによる HMGB1 の不活性化にはトロンビンの存在が必須であること が判明した。このことより、TMαの D1 と HMGB1 の結合はあまり強固ではなく、in vivo では at-HMGB1 と RAGE あるいは ds-HMGB1 と TLR4 の結合を阻害することが できなかったのではないかと推察される(Fig. 5)。LPS によるアロディニアは、LPS 投与 30 分後から有意な閾値の低下を示し、アロデ ィニアの発現に 1~1.5 時間を要する HMGB1 とは大きく異なっていた。これは、LPS 投与初期のアロディニアには、内因性の HMGB1 の関与は少なく、LPS による TLR4 への直接的で効率的な活性化が関与するものと推察される21)。
17
Fig. 5 Scheme for the thrombin-dependent suppression by TMα of the allodynia caused by
at-HMGB1 and ds-HMGB1. In the absence of thrombin, TMα interacts with at-HMGB1 and ds-HMGB1 in a D1 domain-dependent manner, but does not inhibit their pronociceptive activity through RAGE and TLR4, respectively. In the presence of thrombin, TMα sequesters at-HMGB1 and ds-HMGB1 by the D1 domain, and promotes their degradation by thrombin binding to the D2 domain, exhibiting antinociceptive activity.
18
第 2 章 ヒト型可溶性トロンボモジュリンの HMGB1 誘起
アロディニア抑制効果発現における lectin 様ドメイン及び
EGF 様ドメインの必要性
緒言
トロンボモジュリンは、血管内皮細胞に特異的に発現する糖蛋白質で、トロンビン 依存的に血液中の protein C を APC に変換する22)。この APC が凝固カスケードの Va及び VIIIa 凝固因子を阻害することにより抗凝固作用を示す18)。トロンボモジュリン は、細胞外のレクチン様ドメイン(D1)、上皮成長因子(EGF)様ドメイン(D2)及 びセリン/スレオニンに富むドメイン(D3)と、膜貫通ドメイン(D4)及び細胞内ド メイン(D5)により構成される。D2 はトロンビンと結合しプロテイン C の活性化に 寄与する。D1 ドメインは、炎症反応を増悪させる DAMPs の一つである HMGB1 を吸 着することが知られている。D1 への HMGB1 吸着と D2 に結合したトロンビンによる HMGB1 の分解によって、HMGB1 の活性は効率的に抑制される23)。TMαはトロンボ モジュリンの細胞外ドメイン(D1~D3)を遺伝子工学的な手法により CHO 細胞で産 生させた糖蛋白質であり HMGB1 を吸着・分解することが報告されている19)。TMα は、2008 年より本邦において DIC 治療薬として臨床応用されている24,25)。 本章では、TMαによる HMBG1 不活化の作用機序及び抗アロディニア作用を調べ るため、トロンボモジュリンの D1、D2 及び D123 をピキア酵母により構築し26)、in vitro におけるトロンビンによる at-HMGB1 及び ds-HMGB1 の分解作用と、in vivo における at-HMGB1、ds-HMGB1 及び LPS 誘起アロディニアに対する各ドメインの効果を検討 した。
19
I. 実験方法
全ての動物実験は、近畿大学実験動物委員会により承認され、米国国立衛生研究所 による実験動物の管理と使用のためのガイドラインに従って実施した。実験動物は、 3-4 週齢の ddY 系雄性マウス(紀和実験動物、和歌山)を使用した。マウスの飼育は 第 1 章と同様に行った。 トロンボモジュリンの D123(aa1-498)、D1(aa1-155)及び D2(aa224-462)は、 ピキア酵母 [Pichia pastoris strain SMD1163 (his-4pep4 prd1) ] より遺伝子工学的に産生 されたものを旭化成ファーマ株式会社(東京、日本)から提供された 26)。TMαを含 むその他の試薬は第 1 章と同様に入手した。TMα、D123、D1 及び D2 は、0.002% Tween 80 を含む生理食塩水に溶解し、HMGB1 または LPS 投与の 1 時間前に 10 μL の容量で マウスの足底内に投与した。また、10 mg/kg の AT 及び 10 mg/kg の TMαは、LPS 投 与のそれぞれ 1.5 時間及び 1 時間前に腹腔内投与した。各 HMGB1 及び LPS 誘起マウ スアロディニアモデルの作製は第 1 章と同様に行った。 マウスにおける HMGB1 及び LPS 誘起アロディニアの評価は第 1 章と同様に von Frey 法により実施した。 400 nM の at-HMGB1 または ds-HMGB1 は、2 U/mL のトロンビンを含む 40 μL の反 応溶液(2 mM CaCl2、0.1 M NaCl、50 mM Tris–HCl、pH8.0)で 37℃、15 分間インキュベートした後、各 HMGB1 と cleaved products を Western blot により検出した。TMα の効果は、400 nM の TMα存在下または非存在下で比較検討した。その他の条件は第 1 章と同様に行った。
1. 実験動物
2. 使用薬物と投与スケジュール
3. von Frey filaments によるアロディニアの評価
20
統計処理は第 1 章と同様の検定方法を用いて行い、P < 0.05 を有意差ありとした。
21
II. 実験結果
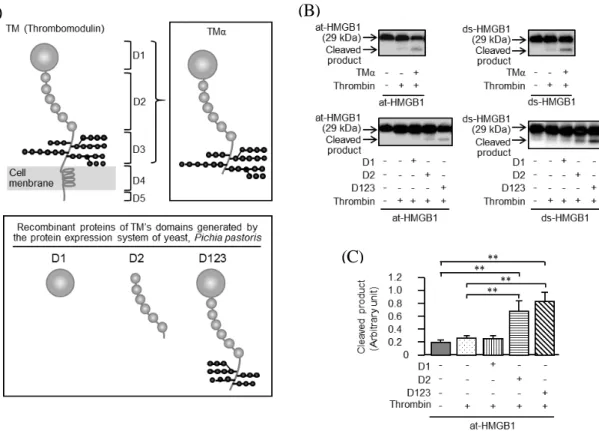
Fig. 6A にトロンボモジュリン、TMα、D1、D2 及び D123 の構造の模式図を示す。 トロンボモジュリンは、575 のアミノ酸で構成された膜一回貫通蛋白質で、5 つのドメ インから構成されている。TMαは、トロンボモジュリンの細胞外ドメインのみを CHO 細胞により産生させた498のアミノ酸で構成された糖蛋白質で、糖鎖構造はヒト型TM と同様である。一方、ピキア酵母により産生させた D1(aa1-155)、D2(aa224-462) 及び D123(aa1-498)のうち、D123 の糖鎖はトロンボモジュリンとは N 型糖鎖構造が 異なるため27)、CHO 細胞で産生された TMα(D3 は O-Gly site rich)とは異なる生理活性を有する可能性も否定できない28)。
Fig. 6B は、400 nM の at-HMGB1 または ds-HMGB1 に 2 U/mL のトロンビンを加え 37℃、15 分間のインキュベ―ション後、Western blot を実施したものである。Cleaved product のバンドは、トロンビン単独ではほとんど変化しなかったが、TMα存在下で トロンビンを作用させることで著しく増加し、第 1 章の結果が再現された。ピキア酵 母により産生した D123 は、TMαと同様、at-HMGB1 と ds-HMGB1 の cleaved product の産生を増加させたが、D1 にはそのような作用は全く認められなかった。Fig. 6C は、 Fig. 6B で at-HMGB1 の Western blot に用いたゲルを densitometry により定量化した結 果である。トロンビンは、単独あるいは D1 存在下では効果を示さなかったが、D123 では cleaved product の産生を有意に増加させた。興味あることに、トロンビンは、D2 の存在下でも cleaved product の産生を僅かに増加させた。
1. トロンビンによる HMGB1 分解に及ぼす酵母産生各種トロンボモジュリン
22
Fig. 6 Effects of TMα and yeast-generated recombinant proteins of human TM’s D1, D2 and
D123 on thrombin-dependent cleavage of at-HMGB1 or ds-HMGB1 in vitro. (A) Structure of TM, TMα and yeast-generated human TM’s D123, D1 and D2. The glycosylation states in the D3 regions of yeast-generated D123 and TMα are considered structurally different. (B) Western blot analysis of the effects of TMα and TM’s D1, D2 and D123 on thrombin-dependent cleavage of HMGB1 thrombin-cleaved products of HMGB1 in vitro. In the absence or presence of TMα or TM’s D1, D2 and D123, at 400 nM, at-HMGB1 or ds-HMGB1 at 400 nM was incubated with 2 U/mL thrombin for 15 min at 37ºC in vitro. The protein levels of at-HMGB1 or ds-HMGB1 and their primary cleaved products were assessed by Western blotting. (C) The amount of the thrombin-cleaved product of at-HMGB1 after 15-min incubation in the absence and presence of TM’s D1, D2 or D123. Western blot data was quantified by densitometry. Data show the means ± S.E.M. for 5 experiments. *P < 0.05, **P <0.01.
(A) (B)
23
2. マウスにおけるHMGB1誘起アロディニアに対するTMαと酵母産生各種トロンボ
モジュリンドメインの効果
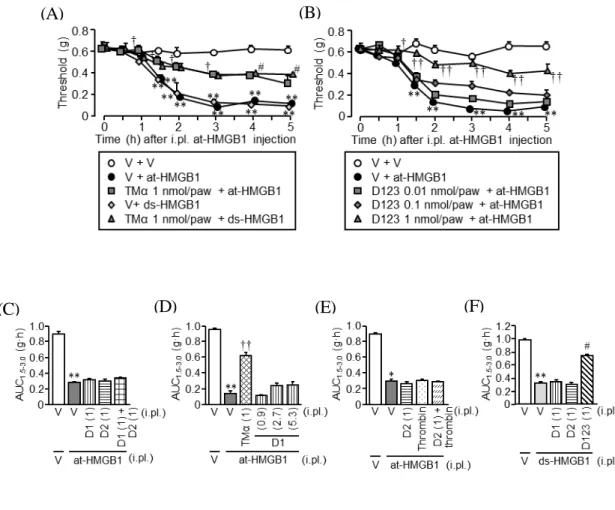
Fig. 7A は、at-HMGB1(100 ng/paw)及び ds-HMGB1(10 ng/paw)誘起アロディニ アに対する TMα の抑制効果を調べた結果である。TMα(1 nmol/paw)は、at-HMGB1 及び ds-HMGB1 誘起アロディニアに対し、ほぼ同等に有意な抑制作用を示した。
Fig. 7B は、ピキア酵母により作製した D123 の at-HMGB1(100 ng/paw)誘起アロデ ィニアに対する効果を調べた結果である。D123 は、0.01、0.1 及び 1 nmol/paw でアロ ディニアを用量依存的に抑制し、1 nmol/paw では同用量の TMα とほぼ同等の抗アロ ディニア効果を示した。
Fig. 7C-F は、at-HMGB1(100 ng/paw)及び ds-HMGB1(10 ng/paw)誘起アロディ ニアに対する各トロンボモジュリンドメインの作用を AUC で示した結果である。D1、 D2 及び D1+D2 を、TMα の有効量と同じ 1 nmol/paw で足底内投与したところ、 at-HMGB1 誘起アロディニアは全く抑制されなかった(Fig. 7C)。また、D1 の投与量 を 2.7 及び 5.3 nmol/paw の髙濃度まで上げて検討したが、抗アロディニア効果は認め られなかった(Fig. 7D)。さらに、in vitro でトロンビンによる HMGB1 分解を促進し たD2 が、in vivo で無効であった原因がトロンビン不足である可能性を検討するため、 トロンビンと D2 の併用投与も行ったが、抗アロディニア効果は認められなかった(Fig. 7E)。同様に、ds-HMGB1(10 ng/paw)誘起アロディニアに対して、1 nmol/paw の D1、D2 及び D123 が抑制効果を示すか否かを調べた結果、D1 及び D2 は無効であっ たが、D123 は TMα と同様の抗アロディニア作用を示した(Fig. 7F)。
24
Fig. 7 Effects of TMα and yeast-generated recombinant proteins of human TM’s D1, D2 and
D123 on the HMGB1-induced mechanical allodynia in vivo. (A, B) Effects of i.pl. TMα (A) and TM’s D123 (B) on the mechanical allodynia induced by i.pl. at-HMGB1 or ds-HMGB1 in mice. The mice received i.pl. administration of TMα at 1 nmol/paw (A) and TM’s D123 at 0.01, 0.1 or 1 nmol/paw (B), 1 h before at-HMGB1 at 100 ng/paw (A, B) or ds-HMGB1 at 10 ng/paw (A). (C-F) Effects of i.pl. TM’s D1, D2 or D123 on the mechanical allodynia induced by i.pl. HMGB1 in mice. The AUC1.5-3.0 was calculated from the time-threshold curve between 1.5 h and 3.0 h after i.pl. HMGB1. The mice received i.pl. administration of TMα and TM’s D1, D2, D123 or combination of D1 and D2, 1 h after i.pl. at-HMGB1 (C-E) or ds-HMGB1 (F). Thrombin at 1 U/paw was co-administered i.pl. with TM’s D2. Figures in parentheses indicate the dose (nmol/paw) of recombinant proteins (C-F). Data show the means ± S.E.M. for 5 (A-C, E-F) or 5-8 (D) mice. *P < 0.05, **P < 0.01 vs. Vehicle (V) + V; †P < 0.05, ††P < 0.01 vs. V + at-HMGB1 or ds-HMGB1.
(A) (B)
25
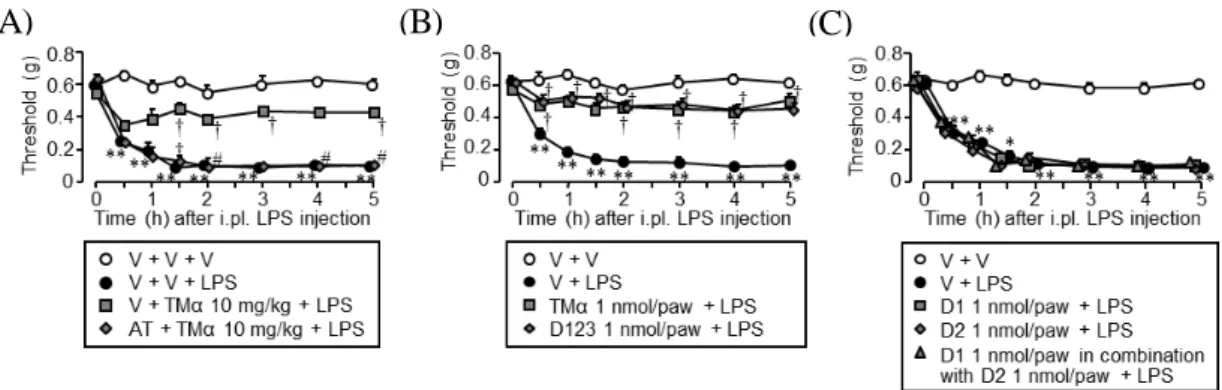
Fig. 8 は、LPS 足底内投与誘起アロディニアに対する TMα及び各種トロンボモジュ リンドメインの効果を検討した結果である。LPS 投与 1 時間前に TMαを 10 mg/kg の 用量で腹腔内投与したところ LPS 誘起アロディニアが有意に抑制された。この TMα の腹腔内投与による抗アロディニア効果は、トロンビン直接阻害剤の AT(10 mg/kg、 i.p.)を LPS 投与 1.5 時間前に投与することで完全に消失した(Fig. 8A)。また、LPS 誘起アロディニアに対し、1 nmol/paw の TMα及び D123 の足底内投与による抗アロ ディニア効果は同等であった(Fig. 8B)。一方、LPS 誘起アロディニアに対し、D1、 D2 及び D1+D2 の足底内投与は、全く抑制作用を示さなかった(Fig. 8C)。
Fig. 8 Effects of TMα and yeast-generated recombinant proteins of human TM’s D1, D2 and
D123 on the LPS-induced “endogenous HMGB1-dependent” mechanical allodynia in vivo. (A) Endogenous thrombin-dependent preventive effect of systemic (i.p.) TMα on the LPS-induced mechanical allodynia in mice. Argatroban (AT), a direct thrombin inhibitor, at 10 mg/kg and TMα at 10 mg/kg were administered i.p. 1.5 h and 1 h, respectively, before i.pl. LPS at 100 ng/paw. (B) TM’s D123 mimics the TMα-induced suppression of LPS-induced mechanical allodynia in mice. The mice received i.pl. administration of TMα or TM’s D123 at 1 nmol/paw, 1 h before i.pl. LPS. (C) The lack of effects of TM’s D1, D2 or their combination on the LPS-induced mechanical allodynia in mice. The mice received i.pl. administration of TM’s D1, D2 or their combination, each at 1 nmol/paw, 1 h before i.pl. LPS. Data show the means ± S.E.M. for 5 mice. *P <0.05, **P <0.01 vs. Vehicle (V) + V (B, C) or V + V +V (A); †P < 0.05, vs. V + LPS (B, C) or V+ V + LPS (A); #P < 0.05 vs. V + TMα + LPS (A).
3. マウスにおいて LPS により誘起される内在性 HMGB1 依存性アロディニアに
対する TMα及び酵母産生各種トロンボモジュリンドメインの効果
26
Fig. 9 A working hypothesis for the mechanism of the effects of TM’s D123, D1 and D2 on the
HMGB1-induced allodynia. (A) The allodynia caused by at-HMGB1 and ds-HMGB1 is mediated by RAGE and TLR4, respectively. (B) TM’s D123 sequesters at-HMGB1 and ds-HMGB1 with the D1 region and promotes their degradation by thrombin (Thr) binding to the D2 region, thereby suppressing the allodynia caused by at-HMGB1 or ds-HMGB1. (C) TM’s D1 does not inhibit the mechanical allodynia caused by at-HMGB1 and ds-HMGB1 in vivo, although it may bind to at-HMGB1 and ds-HMGB1 in vitro. (D) TM’s D2 is not able to block the allodynia caused by at-HMGB1 and ds-HMGB1 in vivo, although it is capable of facilitating the thrombin-induced degradation of at-HMGB1 and ds-HMGB1 in vitro.
27
III. 考察
血管内皮上に特異的に発現するトロンボモジュリンは、抗凝固作用と抗炎症作用を 併せ持つ多機能蛋白質である23)。TM は 5 つのドメイン(D1~D5)から構成されてお り、抗凝固作用及び抗炎症作用の発現には細胞外ドメイン、すなわち D1 から D3 が重 要な働きを示す。遺伝子組換え型可溶性トロンボモジュリン(TMα)は、トロンボ モジュリンの細胞外ドメイン(aa1 - 498)のみを CHO 細胞で産生させた糖蛋白質であ り、DIC 治療薬として臨床応用されている24,25)。しかしながら、様々な疼痛モデル動 物で認められる TMαの鎮痛効果が6,7,9,11,16,17)、どの様な作用機序によるものかは、こ れまでに明らかにされていない。 TM の D1 ドメインは LPS や HMGB1 を吸着し、D2 ドメインはトロンビンまたはプ ロテイン C を結合することが既に知られており 23)、伊藤らは、トロンビンによる HMGB1 の分解反応がTMαにより促進されることを報告している19)。しかしながら、 TMαの HMGB1 誘起アロディニアに対する抑制作用と各ドメインの関与については 明確となっていない。本章では、蛋白質の分泌生産能力に優れているピキア酵母によ り、トロンボモジュリンの D1、D2 及び D123 を産生させ、これら各ドメインが at-HMGB1 と ds-HMGB1 のトロンビン分解に及ぼす影響を調べ、さらに at-HMGB1、 ds-HMGB1 あるいはLPS の足底内投与により誘起されるアロディニアに対する作用を 解析した。 In vitro の検討において、今回用いた濃度のトロンビンは単独では at-HMGB1 を分解 しなかったが、D2 あるいは D123 の存在下ではトロンビンによって at-HMGB1の cleaved product の産生が明らかに増加した。この結果は、蛋白質分解酵素であるトロン ビンが D2 と複合体を形成することにより効率的に at-HMGB1 を分解した可能性を示 唆している。一方、in vivo 試験において、at-HMGB1 及び ds-HMGB1 によるアロディ ニアは、D1、D2 及び D1+D2 では抑制されず、さらに D2 とトロンビンを併用投与し ても抑制効果を確認できなかった。これらのことから、in vitro では D2 はトロンビン と結合し HMGB1 の分解を促進することができるが、RAGE、TLR4 を含む多様な HMGB1 結合蛋白質が存在する生体内では、D1 によって HMGB1 吸着する過程が必須28 で、これによって D123 の D2 部分に結合したトロンビンによる HMGB1 分解が効率 的に進むのではないかと考えられる(Fig. 9)。 TMαにおいて糖鎖修飾を受ける D3 部分は、TMαの水溶性と血中の安定性に関与 していることは既に報告されている28)。ピキア酵母は、蛋白質を効率的に産生するこ とから、遺伝子組換え蛋白質の研究に応用されているが、哺乳類と酵母では、蛋白質 の糖鎖修飾が異なるため27)、同じアミノ酸配列を有する TMαと D123 が同様の生理 活性を示すか否かの検討は重要である。Western blot 法により検討したトロンビンによ る HMGB1 分解反応に対して、等モルの TMαと D123 がほぼ同等の促進効果を示す ことが確認できた。さらに、in vivo において、at-HMGB1 誘起アロディニアに対して、 1 nmol/paw の D123 及び TMαの足底内投与は、ほぼ同等の抑制効果を示した(Fig. 7A, B)。さらに、LPS 誘起アロディニアに対しても TMαと D123 はほぼ同様の抑制効果 を示した(Fig. 8B)。 今回の検討により CHO 細胞により産生させた TMα(aa1 - 498)とピキア酵母によ り産生させた D123(aa1 - 498 + シグナルペプチド)は、等モルにおいて同等の抗アロ ディニア作用を示した。本章で検討した局所(足底内)投与の条件下において、発現 細胞の違いによる TMαと D123 の抗アロディニア作用に違いのないことが明確とな った。一方、D3 は、その長さと糖鎖修飾の違いによりトロンボモジュリン細胞外ドメ インの水溶性と血中安定性に深く関与することが報告されている28)。TMαと D123 の 生物学的同等性を検証するためには、全身投与による抗アロディニア作用を比較・検 討することが今後の課題である。さらに、抗アロディニア作用における D3 の関与を 明らかにするため、D12 による検討が必要である。
29
総括
本研究では、HMGB1 誘起アロディニアに対する TMαの抑制メカニズムを検討し た。 第 1 章では、disulfide 結合の有無による立体構造の異なる 2 つの HMGB1、すなわち at-HMGB1 と ds-HMGB1 がトロンビンにより分解され、この反応が TMαにより増強 されることを検証した。さらに、at-HMGB1 、ds-HMGB1 あるいは LPS の足底内投与 で発症するアロディニアに対し、TMαは用量依存的に抑制効果を示すことが示唆さ れた。TMαの抗アロディニア作用は、トロンビンの直接阻害剤である AT により完全 に消失し、単独ではマウスに影響のない低用量のトロンビン(1 U/paw)を投与するこ とにより抗アロディニア作用が増強されることが明確となった。これらの結果から、 TMαによる種々の HMGB1 及び LPS 誘起アロディニア抑制作用は、トロンビンに依 存することが示唆された。 第 2 章では、トロンボモジュリンの各ドメインの機能に注目し、TMαの抗アロディ ニア作用の機序についてさらなる検討を行った。トロンボモジュリンは D1 から D5 の 5 つのドメインより構成され、TMαは細胞外の D1 から D3 ドメインからなる。ピ キア酵母により、D1、D2 及び D123 を作製し、トロンビンによる at-HMGB1 と ds-HMGB1 の分解反応を in vitro 試験により検討した。D1 は at-HMGB1 と ds-HMGB1 のトロンビンによる分解を促進しなかったが、D2 及び D123 は両 HMGB1 のトロンビ ン分解を促進した。尚、統計学的な差はないものの、D2 よりも D123 の方が、より強 く HMGB1 分解を促進する傾向が認められた。マウスを用いた in vivo 試験において、 TMαとピキア酵母で作製した D123 は、at-HMGB1 あるいは ds-HMGB1 により誘起さ れたアロディニアをほぼ同等に抑制したが、D1 及び D2 は抗アロディニア作用を全く 示さなかった。さらに、LPS 誘起アロディニアに対しても D123 は TMαとは同等の 抗アロディニア作用を示し、D1 及び D2 に抗アロディニア作用は認められなかった。 このことより、D2 は in vitro ではトロンビンによる HMGB1 分解を促進することがで きるが、in vivo での抗アロディニア作用発現には少なくとも連続した D12 の構造が必 要であると推察された。このことは、第 1 章において、TMαを足底内投与した場合 の HMGB1 誘起アロディニアに対する抑制作用が内因性トロンビンに依存するとの結30 果と一致する。第 2 章においても、腹腔内投与した TMαの LPS 誘起アロディニア抑 制作用がトロンビンの直接阻害剤である AT により完全に消失することを確認できた。 以上より、in vivo における HMGB1 依存性アロディニアに対する TMαの抑制効果発 現には、D1 で HMGB1 を吸着し、この HMGB1 を D2 に結合したトロンビンで分解す る必要があることが示唆された。 今回の検討により、HMGB1誘起アロディニアに対するTMαの抑制作用発現には、 トロンビンに依存した HMGB1 の分解が必須であることが判明した。また、in vivo に おいて、TMαがトロンビン依存性に HMGB1 を不活性化するには、D2 のみならず、 D2 に結合した D1 の存在が必須であると推察された。今後、糖鎖構造をもつ D3 の役 割を解明し、D12 の抗アロディ二ア作用を検討する必要がある。 これまでに、膀胱炎に伴う膀胱痛、急性膵炎に伴う膵臓痛及び抗がん剤(paclitaxel, vincristine)誘発末梢神経障害性疼痛(CIPN: chemotherapy-induced peripheral neuropathy) などに内因性 HMGB1 が関与することが、抗 HMGB1 中和抗体用いた研究により明ら かにされており、これらのモデルでは TMαも抗 HMGB1 中和抗体と同等の効果を示
す11,16,17)。現在、ヒトへの応用として、抗がん剤の oxaliplatin 誘発 CIPN に対するトロ
ンボモジュリン製剤の発症抑制効果が臨床治験中である。今回の結果が、TMαの様々 な疼痛モデルや臨床における有効性を裏付ける一助になることを期待している。
31
引用文献
1) Schaefer C, Sadosky A, Mann R, Daniel S, Parsons B, Tuchman M, Anschel A, Stacey BR, Nalamachu S, Nieshoff E. Pain severity and the economic burden of neuropathic pain in the United States: BEAT Neuropathic Pain Observational Study. Clinicoecon
Outcomes Res, 6, 483-496 (2014).
2) Hamdy RC. The decade of pain control and research. South Med J, 94, 753-754 (2001). 3) Venereau E, De Leo F, Mezzapelle R, Careccia G, Musco G, Bianchi ME. HMGB1 as
biomarker and drug target. Pharmacol Res, 111, 534-544 (2016).
4) Agalave NM, Svensson CI. Extracellular high-mobility group box 1 protein (HMGB1) as a mediator of persistent pain. Mol Med, 20, 569-578 (2015).
5) Wan W, Cao L, Khanabdali R, Kalionis B, Tai X, Xia S. The Emerging Role of
HMGB1 in Neuropathic Pain: A Potential Therapeutic Target for Neuroinflammation. J
Immunol Res, 2016, 6430423 (2016).
6) Yamasoba D, Tsubota M, Domoto R, Sekiguchi F, Nishikawa H, Liu K, Nishibori M, Ishikura H, Yamamoto T, Taga A, Kawabata A. Peripheral HMGB1-induced
hyperalgesia in mice: Redox state-dependent distinct roles of RAGE and TLR4. J
Pharmacol Sci, 130, 139-142 (2016).
7) Shibasaki M, Sasaki M, Miura M, Mizukoshi K, Ueno H, Hashimoto S, Tanaka Y, Amaya F. Induction of high mobility group box-1 in dorsal root ganglion contributes to pain hypersensitivity after peripheral nerve injury. Pain, 149, 514-521 (2010).
8) Feldman P, Due MR, Ripsch MS, Khanna R, White FA. The persistent release of HMGB1 contributes to tactile hyperalgesia in a rodent model of neuropathic pain. J
Neuroinflammation, 9, 180 (2012).
9) Tanaka J, Seki Y, Ishikura H, Tsubota M, Sekiguchi F, Yamaguchi K, Murai A, Umemura T, Kawabata A. Recombinant human soluble thrombomodulin prevents peripheral HMGB1-dependent hyperalgesia in rats. Br J Pharmacol, 170, 1233-1241 (2013).
32
10) Nakamura Y, Morioka N, Abe H, Zhang FF, Hisaoka-Nakashima K, Liu K, Nishibori M, Nakata Y. Neuropathic pain in rats with a partial sciatic nerve ligation is alleviated by intravenous injection of monoclonal antibody to high mobility group box-1. PLoS
One, 8, e73640 (2013).
11) Nishida T, Tsubota M, Kawaishi Y, Yamanishi H, Kamitani N, Sekiguchi F, Ishikura H, Liu K, Nishibori M, Kawabata A. Involvement of high mobility group box 1 in the development and maintenance of chemotherapy-induced peripheral neuropathy in rats.
Toxicology, 365, 48-58 (2016).
12) Maeda T, Ozaki M, Kobayashi Y, Kiguchi N, Kishioka S. HMGB1 as a potential therapeutic target for neuropathic pain. J Pharmacol Sci, 123, 301-305 (2013). 13) Ito T. PAMPs and DAMPs as triggers for DIC. J Intensive Care, 2, 67 (2014). 14) Crews FT, Vetreno RP. Mechanisms of neuroimmune gene induction in alcoholism.
Psychopharmacology (Berl), 233, 1543-1557 (2016).
15) Frank MG, Weber MD, Watkins LR, Maier SF. Stress-induced neuroinflammatory priming: A liability factor in the etiology of psychiatric disorders. Neurobiol Stress, 4, 62-70 (2016).
16) Tanaka J, Yamaguchi K, Ishikura H, Tsubota M, Sekiguchi F, Seki Y, Tsujiuchi T, Murai A, Umemura T, Kawabata A. Bladder pain relief by HMGB1 neutralization and soluble thrombomodulin in mice with cyclophosphamide-induced cystitis.
Neuropharmacology, 79, 112-118 (2014).
17) Irie Y, Tsubota M, Ishikura H, Sekiguchi F, Terada Y, Tsujiuchi T, Liu K, Nishibori M, Kawabata A. Macrophage-derived HMGB1 as a Pain Mediator in the Early Stage of Acute Pancreatitis in Mice: Targeting RAGE and CXCL12/CXCR4 Axis. J
Neuroimmune Pharmacol, 12, 693-707 (2017).
18) Yang L, Rezaie AR. The fourth epidermal growth factor-like domain of
thrombomodulin interacts with the basic exosite of protein C. J Biol Chem, 278, 10484-10490 (2003).
19) Ito T, Kawahara K, Okamoto K, Yamada S, Yasuda M, Imaizumi H, Nawa Y, Meng X, Shrestha B, Hashiguchi T, Maruyama I. Proteolytic cleavage of high mobility group
33
box 1 protein by thrombin-thrombomodulin complexes. Arterioscler Thromb Vasc Biol,
28, 1825-1830 (2008).
20) Chaplan SR, Bach FW, Pogrel JW, Chung JM, Yaksh TL. Quantitative assessment of tactile allodynia in the rat paw. J Neurosci Methods, 53, 55-63 (1994).
21) Ryu JK, Kim SJ, Rah SH, Kang JI, Jung HE, Lee D, Lee HK, Lee JO, Park BS, Yoon TY, Kim HM. Reconstruction of LPS Transfer Cascade Reveals Structural
Determinants within LBP, CD14, and TLR4-MD2 for Efficient LPS Recognition and Transfer. Immunity, 46, 38-50 (2017).
22) Esmon CT. Inflammation and thrombosis: the impact of inflammation on the protein C anticoagulant pathway. Haematologica, 80, 49-56 (1995).
23) Ito T, Maruyama I. Thrombomodulin: protectorate God of the vasculature in thrombosis and inflammation. J Thromb Haemost, 9 Suppl 1, 168-173 (2011).
24) Saito H, Maruyama I, Shimazaki S, Yamamoto Y, Aikawa N, Ohno R, Hirayama A, Matsuda T, Asakura H, Nakashima M, Aoki N. Efficacy and safety of recombinant human soluble thrombomodulin (ART-123) in disseminated intravascular coagulation: results of a phase III, randomized, double-blind clinical trial. J Thromb Haemost, 5, 31-41 (2007).
25) Aikawa N, Shimazaki S, Yamamoto Y, Saito H, Maruyama I, Ohno R, Hirayama A, Aoki Y, Aoki N. Thrombomodulin alfa in the treatment of infectious patients
complicated by disseminated intravascular coagulation: subanalysis from the phase 3 trial. Shock, 35, 349-354 (2011).
26) Ikezoe T, Yang J, Nishioka C, Pan B, Xu K, Furihata M, Nakamura K, Yurimoto H, Sakai Y, Honda G, Yokoyama A. The fifth epidermal growth factor-like region of thrombomodulin exerts cytoprotective function and prevents SOS in a murine model.
Bone Marrow Transplant, 52, 73-79 (2017).
27) Hamilton SR, Davidson RC, Sethuraman N, Nett JH, Jiang Y, Rios S, Bobrowicz P, Stadheim TA, Li H, Choi BK, Hopkins D, Wischnewski H, Roser J, Mitchell T, Strawbridge RR, Hoopes J, Wildt S, Gerngross TU. Humanization of yeast to produce complex terminally sialylated glycoproteins. Science, 313, 1441-1443 (2006).
34
28) Honda G, Masaki C, Zushi M, Tsuruta K, Sata M. The roles played by the D2 and D3 domains of recombinant human thrombomodulin in its function. J Biochem, 118, 1030-1036 (1995).
35
略語一覧
APC activated protein C AT argatroban
AUC area under the blood concentration-time curve CIPN chemotherapy-induced peripheral neuropathy CHO Chinese hamster ovary
DAMPs damage-associated molecular patterns DIC disseminate intravascular coagulation DNA deoxyribonucleic acid
EGF epidermal growth factor HMGB1 high mobility group box 1
at-HMGB1 all-thiol high mobility group box 1
bt-HMGB1 bovine thymus-derived high mobility group box 1 ds-HMGB1 disulfide high mobility group box 1
ox-HMGB1 oxidized high mobility group box 1 LPS lipopolysaccharide
NSAIDs non-steroidal anti-inflammatory drugs PAMPs pathogen-associated molecular patterns PRRs pattern recognition receptors
QOL quality of life
RAGE receptor for advanced glycation endproducts TLR4 Toll-like receptor 4
TM thrombomodulin TMα thrombomodulin alfa
36
主論文
本論文の内容は以下の学術雑誌に公表した。
TSUJITA, R., TSUBOTA, M., HAYASHI, Y., SAEKI, H., SEKIGUCHI, F.,
KAWABATA, A.
Role of thrombin in soluble thrombomodulin-induced suppression of peripheral HMGB1- mediated allodynia in mice.
Journal of Neuroimmune Pharmacology, in Press (2017).
https://doi.org/10.1007/s11481-017-9773-2.
HAYASHI, Y., TSUJITA, R. (co-first author), TSUBOTA, M., SAEKI, H.,
SEKIGUCHI, F., HONDA, G., KAWABATA, A.
Human soluble thrombomodulin-induced blockade of peripheral HMGB1-dependent allodynia in mice requires both the lectin-like and EGF-like domains.