Oxidative modification of methionine80 in cytochrome c by reaction with peroxides
42
0
0
全文
(2) Abstract The Met80‒heme iron bond of cytochrome c (cyt c) is cleaved by the interaction of cyt c with cardiolipin (CL) in membranes. The Met80 dissociation enhances the peroxidase activity of cyt c and triggers cyt c release from mitochondrion to the cytosol at the early stage of apoptosis. This paper demonstrates the selective oxidation of Met80 for the reaction of ferric cyt c with a peroxide, meta-chloroperbenzoic acid (mCPBA), in the presence of CL-containing liposomes by formation of a ferryl species (Compound I). After the reaction of cyt c with mCPBA in the presence of 1,2-dioloeyl-sn-glycero-3phosphocholine (DOPC) liposomes containing CL, the electrospray ionization mass spectrum of the peptide fragments, obtained by digestion of cyt c with lysyl endopeptidase, exhibited a peak at m/z=795.45; whereas, this peak was not observed for the peptide fragments obtained after the reaction in the presence of DOPC liposomes not containing CL. According to the tandem mass spectrum of the m/z=795.45 peptide fragment, Met80 was modified with a 16 Da mass increase. The purified Met80-modified cyt c exhibited a peroxidase activity more than 5-fold higher than that of the unmodified protein. Transient absorption bands around 650 nm were generated by the reactions with mCPBA for ferric wild-type cyt c in the presence of CL-containing DOPC liposomes and ferric Y67F cyt c in the absence of liposomes. The formation and decomposition rates of the 650-nm absorption species increased and decreased, respectively, by increasing the mCPBA concentration in the reaction, indicating transient formation of Compound I.. Keywords: cytochrome c; methionine oxidation; peroxidase reaction; self-modification. 2 .
(3) 1. Introduction Cytochrome c (cyt c) is an electron transfer heme protein in the respiratory chain of mitochondria. Cyt c exists in the inter-membranous space of mitochondria, and its release to the cytosol initiates apoptosis [1, 2]. Horse cyt c possesses many lysine residues on its surface and is positively charged. It also possesses two methionines, Met65 and Met80. The heme of cyt c is covalently bound to the polypeptide chain through two thioether linkages with cysteine residues, and His18 and Met80 are coordinated to the heme iron [3, 4]. Cardiolipin (CL) is a phospholipid component of the mitochondrial inner membrane [5]. Interaction of positively charged cyt c with negatively charged CL in mitochondria plays a crucial role in triggering apoptosis by mediating release of cyt c to the cytosol [6]. Cyt c exhibits a less packed tertiary conformation upon interaction with CL [7]. According to time-resolved fluorescence resonance energy transfer measurements, critical stabilizing interactions between the N- and C-termini of cyt c are broken for some of the protein conformers by interaction with CL [8]. The peroxidase activity of cyt c is low in its native form [9, 10], whereas it increases dramatically by the interaction with CL [11, 12]. This increase may be caused by the opening of the protein with Met80‒heme iron bond breaking and increase in ligand accessibility to the heme [11, 12]. The heme iron of ferric cyt c is ligated with His/Lys or His/OH− at pH 7.4 and the redox potential shifts negatively when it interacts with CL [13, 14]. It has been reported that inner membrane-bound cyt c is released to the intermembrane space by alterations in the membrane structure by the CL oxidation [15, 16]. CL has been shown to be oxidized by the peroxidase reaction of the cyt c‒CL complex [17, 18], and the CL oxidation may proceed via generation of the Tyr67 radical in cyt c [19]. In fact, docking studies of cyt c 3 .
(4) with linoleic acid, the most common fatty acid component of CL, showed that C11 of linoleic acid can bind to cyt c at a position adjacent to the heme and Tyr67 [20]. Tyr67 forms a hydrogen bond network with Met80, conserved Asn52 and Thr78, and a water molecule inside the protein, which stabilizes the Met80 coordination to the heme iron [3, 4]. Tyr67 also participates in modifying the local flexibility of a nearby polypeptide chain segment (residues 65 to 72) [21]. In the Tyr67 variants of cyt c, the Met80‒heme iron bond is disrupted and the peroxidase activity increases [22-24]. The Met80‒heme iron bond is also perturbed in ferric dimeric cyt c, which exhibits a higher peroxidase activity compared to the monomer [25, 26]. The two methionine residues of horse cyt c have been shown to be oxidized to methionine sulfoxide by a reaction with an excess amount of hypochlorous acid (HOCl), with the cyt c peroxidase activity increasing through the oxidation [27]. It has been reported that an increase in intra-mitochondrial reactive oxygen species oxidizes cyt c at Met80 [28], whereas Tyr67 of cyt c is highly oxidized by the reaction of native cyt c with H2O2 and the cyt c peroxidase activity increases by the oxidation [29]. For methionine oxidation, an enzyme, methionine-S-sulfoxide reductase A (MsrA), may reduce the methionine sulfoxide in cyt c to its unmodified methionine [30]. We have previously shown that Met80 of cyt c is oxidized site-specifically by the formation of an oxy and subsequent Compound I-like species, when Met80 dissociates from the heme iron [31]. However, the detailed reaction mechanism of Met80 oxidation with peroxides for cyt c is yet to be revealed. In this study, we show that Met80 of ferric cyt c is selectively oxidized in the presence of CL-containing liposomes by dissociation of Met80 from the heme iron and formation of a ferryl species (Compound I).. 4 .
(5) 2. Materials and methods 2.1. Preparation of monomeric, dimeric, and Met80-modified horse cyt c Ferric horse cyt c was prepared by dissolving 100 mg of horse cyt c (Sigma Aldrich) in 10 mL of 50 mM potassium phosphate buffer, pH 7.0. The cyt c solution was filtrated (Millipore; pore size 0.45 μm) and purified by gel chromatography (Hiload 26/60 Superdex 75, GE healthcare) using a fast flow liquid chromatography (FPLC) system (BioLogic DuoFlow 10, Bio-Rad) with the same buffer at 4 °C. After an addition of potassium ferricyanide (Wako, Japan) (10 equivalents to the heme) to the cyt c solution, potassium ferricyanide was removed from the protein solution by a DE52 (Whatman, UK) column with the same buffer. Ethanol was added up to 60% (v/v) to horse cyt c (Sigma Aldrich) in 50 mM potassium phosphate buffer, pH 7.0, producing cyt c precipitates [25]. The obtained precipitate was lyophilized and dissolved in the same buffer, resulting in the formation of cyt c oligomers, and dimeric cyt c was purified from the mixture of various sizes of cyt c oligomers as previously described [25]. To obtain Met80-modified cyt c, oligomeric cyt c (heme concentration: 100‒150 μM) was incubated with ten equivalents of meta-chloroperbenzoic acid (mCPBA) to heme in the same buffer at 35 °C for 30 min. After removal of mCPBA from the protein solution using an Amicon Ultra ultrafiltration tube (Merck Millipore, 3000 NMWL), cyt c oligomers were dissociated to monomers by incubation at 60 °C for 30 min. Modified monomeric cyt c was purified with an ion exchange (CM-52, Whatman) column, and subsequently with gel filtration (HiLoad 26/60 Superdex 75) and ion exchange (Mono S 5/50 GL, GE Healthcare) columns using the FPLC system (BioLogic DuoFlow 10). Cyt 5 .
(6) c modification was confirmed by a matrix-assisted laser desorption ionization-time of flight (MALDI-TOF) mass measurement with an Autoflex II mass spectrometer (Bruker Daltonics) using sinapinic acid as a matrix in linear mode.. 2.2. Preparation of Y67F horse cyt c Point mutation of Try67 to Phe was performed by PCR-based in vitro mutagenesis of the horse cyt c expression plasmid (containing the heme lyase gene) using forward and reverse primers (Eurofins genomics) (Table S1 in Suppl. mat.). Plasmid DNAs were prepared using the PrimeSTAR max DNA polymerase (Takara Bio). DNA sequencing was carried out with the BigDye Terminator v3.1 cycle sequencing kit (Applied Biosystems, Inc., Foster City, CA) and an ABI PRISM 310 genetic analyzer sequencing system (Applied Biosystems, Inc.). The obtained plasmid was introduced into the competent cells of E. coli Rosetta2 (DE3) pLysS (Novagen). Ferric Y67F horse cyt c was purified with a similar procedure reported for wild-type human cyt c [32]. Purification of Y67F cyt c without an N-acetyl group was confirmed by a MALDI-TOF mass spectrum measured under the same conditions described above.. 2.3. Preparation of liposomes Liposomes were prepared from 1,2-dioloeyl-sn-glycero-3-phosphocholine (DOPC, Avanti Polar Lipids, Inc.) with and without bovine heart CL (Avanti Polar Lipids). DOPC (6 mM) was dissolved in 1 mL CH2Cl2 with and without CL (6 mM) at room temperature. After evaporation of the DOPC solution with an evaporator, 1 mL of 25 mM HEPES buffer, pH 7.4, was added to the precipitate at room temperature. The solution was shaken with a vortex for 1−2 min at room temperature, and ultrasonicated for 1−2 min at 25 °C. 6 .
(7) The ultrasonicated solution was frozen with liquid nitrogen and incubated at 50 °C until the sample was melted. The shaking, ultrasonic, freezing, and melting were repeated three times. Liposomes were added to the protein solutions at room temperature.. 2.4. Mass measurements Ferric horse cyt c (0.3 mM) was incubated with liposomes (3 mM DOPC with and without 3 mM CL) in the presence of mCPBA (0.4 mM) in 25 mM HEPES buffer, pH 7.4, for 10 min at room temperature. For the reaction of cyt c with mCPBA in the presence of CL without DOPC, CL was treated in the same way as for obtaining DOPC liposomes. For comparison of the reactivity of monomeric and dimeric cyt c with peroxides, cyt c (heme concentration: 0.3 mM) was incubated with mCPBA or H2O2 (0.4 mM) under the same conditions. After the reaction of cyt c with mCPBA or H2O2, the buffer was exchanged with 50 mM ammonium carbonate buffer, pH 7.8, using the Amicon Ultra ultrafiltration tube (Merck Millipore, 3000 NMWL) to remove excess mCPBA from the solution. The obtained cyt c (0.3 mM) was digested by incubation with 0.05 μg/μL lysyl endopeptidase at 37 °C for 12 h. The digested samples were diluted 20 times with 0.05% formic acid and 50% acetonitrile aqueous solution. Electrospray ionization mass (ESI-MS) spectra of the digested samples were measured with an AccuTOF mass spectrometer (JEOL). Liquid chromatography-tandem mass (LC-MS/MS) analyses were performed with a LCMS-8030 triple quadrupole mass spectrometer using a Nexera UHPLC system (Shimadzu). Chromatographic separations of LC-MS/MS analyses were carried out using an ODS column (Shim-pack XR-ODS II, 150 mm × 2.0 mm, 2.2 μm, Shimadzu) at 40 °C. MALDI-TOF mass spectra of cyt c were obtained with the Autoflex II mass spectrometer (Bruker Daltonics) using sinapinic acid 7 .
(8) as a matrix in linear mode.. 2.5. Peroxidase activity measurements Peroxidase activities of Met80-modified and ferric unmodified horse cyt c were measured by mixing cyt c (2 μM) in 50 mM potassium phosphate buffer, pH 7.0, containing 100 μM 2,2'-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) with 0–100 mM H2O2 at 25 °C using a RSP-601 stopped-flow apparatus (Unisoku). The reaction mixture contained 1 μM cyt c, 50 μM ABTS, and 0–50 mM H2O2. Formation of ABTS cation radicals was monitored at 730 nm. The rate constant was obtained as the maximum of the first derivative of the product formation rate. The extinction coefficient of ferric Met80-modified cyt c was determined to be (1.19 ± 0.03) × 105 M−1cm−1 at 404.5 nm by the pyridine hemochrome method [33], and the concentration of ferric Met80modified cyt c in solution was calculated using the extinction coefficient.. 2.6. Stopped-flow measurements Ferric horse cyt c (80‒100 μM) in 50 mM potassium phosphate buffer, pH 7.0, was prepared containing DOPC (0.2 mM) with or without CL (0.2 mM) at room temperature. Solution of mCPBA (1‒5 mM) was prepared with the same buffer. Each solution was filtered through the 0.45 μm filter (Millipore) before use. The absorption changes of cyt c were monitored after mixing cyt c and mCPBA solutions at a 1:1 ratio at 25 °C using the RSP-601 stopped-flow apparatus (Unisoku). Absorbance changes at 650 nm for the reaction of Y67F cyt c (50 μM) with mCPBA (0.5‒5 mM) in 50 mM potassium phosphate buffer, pH 7.0, at 35 °C was monitored with the RSP-601 stopped-flow apparatus (Unisoku). The absorbance time courses were least-square fitted with sequential reaction 8 .
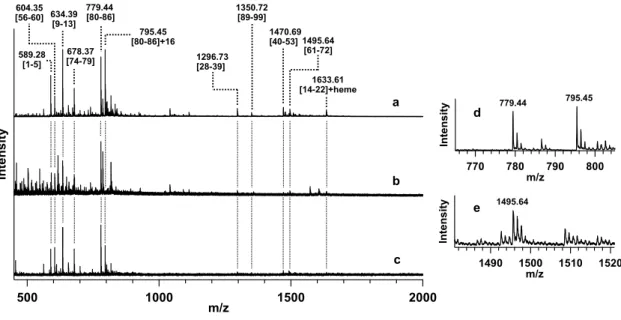
(9) equations. Transient absorption spectra and time-dependent 650-nm absorbance changes were averaged for at least 5 and 20 measurements, respectively.. 2.7. Circular dichroism measurements Circular dichroism (CD) spectra of ferric Met80-modified and unmodified horse cyt c (8 μM) were measured with a J-725 CD spectropolarimeter (Jasco) using a 0.1-cm pathlength quartz cell at 20 °C. Protein solutions in 50 mM potassium phosphate buffer, pH 7.0, were filtered through the 0.45 μm filter (Millipore) before measurements.. 3. Results and discussion 3.1. Met80 modification of cyt c by reaction with mCPBA in the presence of CL-containing DOPC liposomes mCPBA has been used as an oxidant to investigate the heterolytic cleavage of the oxygen−oxygen bond on peroxide binding to the heme iron in mutant myoglobins [34, 35]. Since cyt c decomposes rapidly by the reaction with excess hydrogen peroxide (H2O2) [9], and mCPBA is more reactive than H2O2, the reaction of cyt c with a semiequivalent amount of mCPBA to cyt c was studied. However, the absorption spectrum of horse cyt c did not change significantly by incubation with mCPBA for 60 min (see Fig. S1). ESI-MS measurements were performed for the peptide fragments obtained by cyt c digestion with lysyl endopeptidase after the reactions of ferric horse cyt c with mCPBA in the presence and absence of DOPC liposomes for 10 min at room temperature. The ESI-MS spectra exhibited mass peaks for all the expected fragments with masses more than 500 Da: m/z = 589.28, 604.35, 634.39, 678.37, 779.44, 1296.73, 1350.72, 1470.69, 1495.64, and 1633.61 (calculated monoisotopic masses (Mm) for protonated fragments: 9 .
(10) 589.28, 604.35, 634.39, 678.38, 779.45, 1296.72, 1350.73, 1470.69, 1495.70, and 1633.61, respectively) (Figs. 1 and S2). The mass of the peak at m/z = 779.44 corresponded well to that of the digested peptide with an amino acid sequence of MIFAGIK (Met80−Lys86). A relatively intense additional mass peak was observed at m/z = 795.45, which corresponded well to the mass with an addition of 16 Da to that of the Met80−Lys86 peptide (Fig. 1, spectra a and d), suggesting that an oxygen atom was attached to the peptide. No other intense peak was observed in the mass spectrum with an increase of 16 Da from the masses of expected peptide fragments, including the m/z = 1495.64 fragment, which contained a methionine residue and Tyr67 (Fig. 1, spectrum e). The m/z = 795.45 peak was not observed in the ESI-MS spectrum of the cyt c peptide fragments obtained by the digestion after the reaction in the presence of DOPC liposomes not containing CL (Fig. 1, spectrum b). The peak was observed in the spectrum of the fragments obtained by the digestion after the reaction in the presence of CL without DOPC (Fig. 1, spectrum c); however, the oxidation at Met80−Lys86 for cyt c by the reaction with mCPBA was more prominent in the presence of CL-containing DOPC liposomes compared to that in the presence of CL liposomes, suggesting that CLcontaining DOPC liposomes facilitate the oxidation of cyt c at Met80−Lys86. Additionally, the expected fragments with masses more than 500 Da and an extra fragment with that of m/z = 795.44 were detected also by the LC-MS analysis of the horse cyt c peptide fragments obtained by digestion after the reaction with mCPBA in the presence of CL-containing DOPC liposomes (Fig. S3). These results indicate that the Met80−Lys86 region of cyt c was selectively oxidized by mCPBA in the presence of CL. To identify the modified amino acid in the peptide fragment with m/z = 795.45 obtained by the reaction of cyt c with mCPBA in the presence of CL-containing DOPC 10 .
(11) liposomes, a LC-MS/MS measurement was performed. Mass peaks corresponding to [IFAGIK]+ (m/z: 648.5, Mm: 648.4), [FAGIK]+ (m/z: 535.2, Mm: 535.3), [AGIK]+ (m/z: 388.4, Mm: 388.3), [GIK]+ (m/z: 317.0, Mm: 317.2), [IK]+ (m/z: 260.2, Mm: 260.2), and [K]+ (m/z: 147.0, Mm: 147.1) were observed in the MS/MS spectrum (Fig. 2). These results show that the amino acid sequence of the modified peptide fragment contained the IFAGIK sequence. Furthermore, an extra peak at m/z = 120.0, equivalent to the mass of a methionine sulfoxide immonium ion (Mm: 120.0), was observed in the mass spectrum, indicating that the methionine of the Met80–Lys86 peptide was modified. Both Met65 and Met80 in horse cyt c have been reported to be oxidized by highly-reactive HOCl [27]; thus, the selective oxidation of Met80 by mCPBA may be performed by a specific reaction. Recently, it has been reported that H2O2 oxidizes Tyr67 and lysine residues before Met80 [29]. Met80 and Tyr67 are both positioned close to the heme, and both amino acids have an opportunity to be oxidized by the peroxidase reaction at the heme site. Since the relative positions of the amino acids to the heme would have large effects on deciding which amino acid is modified, the structure of cyt c is important on determining the amino acid being modified. However, the difference in the nature of the peroxide may also determine the modified amino acid, and further investigations are necessary to clarify the differences.. 3.2 Met80 modification of dimeric cyt c by reaction with mCPBA Met80 is reported to dissociate from the heme iron in cyt c by interaction with CL [8]. To elucidate the effect of the Met80 dissociation on its oxidative modification, the reaction of ferric dimeric horse cyt c with mCPBA was investigated, since the Met80‒ heme iron bond is perturbed significantly in dimeric cyt c [25]. The ESI-MS spectrum of 11 .
(12) the peptide fragments obtained by digestion of the proteins with lysyl endopeptidase after the reaction of dimeric cyt c with mCPBA in the absence of CL also exhibited peaks for all the expected fragments with masses more than 500 Da: m/z = 589.27, 604.34, 634.39, 678.37, 779.43, 1296.71, 1350.71, 1470.66, 1495.66, and 1633.59 (Fig. 3, spectrum a). The peak at m/z = 795.44, which corresponded well to the mass with an addition of 16 Da to the Met80−Lys86 peptide mass, was also observed in the spectrum (Fig. 3, spectra a and e). No other intense peak was observed in the mass spectrum with an increase of 16 Da from the masses of expected peptide fragments, including the Met-containing m/z = 1495.64 fragment (Fig. 3, spectrum f); the ESI-MS mass spectrum was similar to that of the peptide fragments obtained by cyt c digestion after the reaction of monomeric cyt c with mCPBA in the presence of CL-containing DOPC liposomes (Fig. 1, spectrum a and Fig. 3, spectrum a). The peak at m/z = 795.44 was also observed in the mass spectrum of the peptide fragments obtained by digestion of monomeric cyt c after the reaction with mCPBA, but with much lower intensity (Fig. 3, spectrum b). A small amount of Met80 oxidation in monomeric cyt c was presumably due to a small amount of Met80 dissociation from the heme iron during the reaction with a relatively strong oxidant mCPBA. For the reaction with H2O2, the ESI-MS spectrum of peptide fragments obtained by digestion of cyt c with lysyl endopeptidase after the reaction of dimeric cyt c in the absence of CL exhibited peaks for all the expected fragments with masses more than 500 Da (Fig. 3, spectrum c). Additionally, the peak at m/z = 795.44, which corresponded to the mass with an addition of 16 Da to the Met80−Lys86 peptide mass, was also observed in the ESI-MS spectrum (Fig. 3, spectrum c), whereas the intensity of this peak decreased in the spectrum of peptide fragments obtained by the reaction of monomeric cyt c with H2O2 in the absence of CL (Fig. 3, spectrum d). These results demonstrate that the 12 .
(13) Met80−Lys86 region of cyt c is selectively oxidized for the reaction of dimeric cyt c even in the absence of CL, supporting the idea that the Met80 dissociation from the heme iron is important for selective Met80 oxidation. In the tandem mass spectrum of the protonated ion of the peptide fragment with m/z = 795.44 obtained by the digestion after the reaction of ferric dimeric cyt c with mCPBA, mass peaks corresponding to [IFAGIK]+ (m/z: 648.3, Mm: 648.4), [FAGIK]+ (m/z: 535.3, Mm: 535.3), [AGIK]+ (m/z: 388.2, Mm: 388.3), [GIK]+ (m/z: 317.1, Mm: 317.2), [IK]+ (m/z: 260.1, Mm: 260.2), [K]+ (m/z: 147.1, Mm: 147.1), and methionine sulfoxide immonium ion (m/z: 120.1, Mm: 120.0) were observed (Fig. S4). These results indicate that Met80 of cyt c was oxidized by the reaction of the dimer with mCPBA. It has been reported that the interaction of cyt c with CL leads to destabilization of the heme–Met80 bond and increases its peroxidase activity [14]. Destabilization of the heme–Met80 bond of cyt c allows binding of external ligands such as carbon monoxide and nitric oxide to the ferrous and ferric heme irons, respectively [36, 37]. Replacement of Met80 with alanine in cyt c creates a binding site for external ligands to the heme iron [38-40]. In dimeric cyt c, Met80 coordination to the heme iron is perturbed [25]. Therefore, the destabilization of the Met80‒heme iron bond may allow mCPBA and H2O2 to bind to the heme and selectively oxidize Met80. These results indicate that the destabilization of the Met80‒heme iron bond of cyt c increases the peroxidase activity, which is related to apoptosis [11, 12].. 3.3 Peroxidase activity of Met80-modified cyt c To investigate the effect of site-specific Met80 oxidation on cyt c peroxidase activity in detail, we modified horse cyt c by the reaction of cyt c oligomers with mCPBA and 13 .
(14) measured the peroxidase activity of the purified Met80-modified cyt c. An intense peak was observed at m/z = 12377 in the MALDI-TOF mass spectrum of Met80-modified cyt c, whereas a single peak was observed at m/z = 12360 for that of unmodified cyt c (calculated molecular weight of positively charged cyt c: 12360) (Fig. S5), showing that modification of cyt c and purification of the modified protein were successful. The oxidation rate of ABTS resulting from the peroxidase activity of Met80-modified and unmodified cyt c followed the equation for a bimolecular reaction:. v = k1[cyt c][H2O2]. (1). = kobs[cyt c]. (2). where v and kobs represent the observed peroxidase reaction rate and the rate constant, respectively, where kobs = k1[H2O2]. The k1 values for Met80-modified and unmodified cyt c were obtained from the slopes of the plots as 94 ± 2 and 18 ± 1 M-1s-1, respectively (Fig. 4); the peroxidase activity of Met80-modified cyt c being more than 5-fold higher than that of unmodified cyt c (Fig. 4). The intensity of the negative band at 208 nm in the CD spectra of Met80-modified cyt c decreased slightly compared to that of the unmodified protein, indicating that the secondary structures of cyt c decreased only slightly by Met80 modification and the protein mostly maintained its secondary structures (Fig. S6). It has been reported that the peroxidase activity of cyt c increases by the rupture of the Met80–heme iron bond [41, 42]. Therefore, the higher activity for Met80-modified cyt c is apparently due to the dissociation of Met80 from the heme iron allowing H2O2 to bind to the heme easier, whereas the Met80–heme iron bond of unmodified cyt c may also be perturbed a little by the addition of H2O2. It has been reported that the peroxidase 14 .
(15) activity of cyt c increases by oxidation of cyt c with HOCl [27], or by carboxymethylation at Met65 and Met80 [42]. The apoptotic activity of cyt c may increase due to the decrease in Met80 coordination to the heme iron by the oxidative modification of Met80.. 3.4 Kinetic measurements of the reaction of cyt c with mCPBA Stopped-flow analyses were performed to elucidate the reaction mechanism of cyt c with mCPBA in the presence of CL-containing DOPC liposomes in potassium phosphate buffer (Fig. 5). We used a phosphate buffer, since Met80 of cyt c has been shown to be oxidized efficiently with H2O2 in the potassium phosphate buffer [43]. The absorption of ferric cyt c around 530 nm decreased, whereas that around 650 nm increased by the reaction with mCPBA in the presence of CL-containing DOPC liposomes (Fig. 5A). In the presence of DOPC liposome without CL, the absorption around 530 nm decreased but that around 650 nm did not change (Fig. 5B). The decrease in the absorption at 530 nm implies decomposition of the protein, making it difficult to further study the Met80 oxidation with the wild-type protein. Tyr67 forms a hydrogen bond network with Met80 and also Asn52 and Thr78, stabilizing the Met80‒heme iron bond [3, 4]. The protein entatic contribution to the strength of the Met80‒heme iron bond of cyt c is ~4 kcal/mol, and the bond can be broken by interrupting the hydrogen bond network connected to Met80 [44]. To investigate the kinetics of the reaction of Met80-dissociated cyt c with mCPBA, we used Y67F horse cyt c, in which the hydrogen bond network including Met80 is modified [21]. The absorption spectrum of ferric Y67F cyt c exhibits an absorption band around 695 nm related to the Met80‒heme iron coordination, although slightly red shifted for 5 nm, indicating Met80 coordination to the heme iron (Fig. S7). In addition, the mass of Y67F cyt c increased 16 15 .
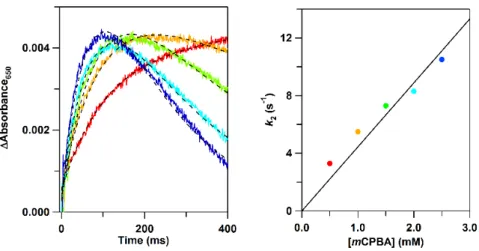
(16) from m/z = 12302 (calculated molecular weight of positively charged Y67F cyt c: 12302) to m/z = 12318 by the reaction with mCPBA ([cyt c]:[mCPBA] = 1:2) in the absence of liposomes, indicating oxidative modification of Y67F cyt c by the reaction (Fig. S8). By the stopped-flow analysis for the reaction of ferric Y67F cyt c with mCPBA in the absence of liposomes, an increase and a decrease in the absorption around 650 and 530 nm, respectively, were also observed (Fig. 5C). A small absorption increase was also observed at about 433 nm by stopped-flow monitoring the absorption changes in the Soret region for the reaction of ferric Y67F cyt c with mCPBA (Fig. S9). Although the Y67F cyt c concentration had to be decreased to one-tenth of that of the experiments monitoring the absorption at 650 nm, due to the strong Soret absorption, the relative absorption change at 433 nm was larger than that at 650 nm, taking into account the cyt c concentration (Figs. 5C and S9). Time courses were measured for the reaction of Y67F cyt c with mCPBA, since the absorption changes were detected in the absence of liposomes. The concentration of mCPBA was varied to verify the formation of the 650-nm absorption species by the reaction of Y67F cyt c with mCPBA (Fig. 6A). The absorption at 650 nm increased, and subsequently decreased for all the conditions measured. The 650-nm absorption band intensity increased faster, and also it subsequently decreased faster, for higher mCPBA concentrations. The 650-nm absorbance changes were least-square fitted with the following sequential reaction equation,. ∆Abs =. {exp(−k2t) – exp(−k3t)} + b. (3). where t represents the reaction time, k2 and k3 represent the rate constants for the initial 16 .
(17) and subsequent reactions, respectively, and a and b represent constants. The rate constant k2 increased linearly as the mCPBA concentration was increased (Fig 6B), supporting the transient formation of the 650-nm species by the reaction of cyt c with mCPBA.. 3.5 Mechanism for the reaction of cyt c with mCPBA A peroxidase Compound I-like intermediate, a ferryl porphyrin π-cation radical species, has been reported to form by the reaction of cyt c with H2O2 [45]. Formation of Compound I has also been hypothesized for the reaction of Y67R cyt c with H2O2 by monitoring a small absorption change at 434 nm [24]. We have previously shown that by interaction of cyt c with CL-containing DOPC liposomes, the dissociated Met80 is selectively oxidized to methionine sulfoxide with O2, through a formation of oxy and Compound I-like species in the presence of reducing agents [31]. In carboxymethylated cyt c, Met80 was carboxymethylated and the Met80−heme iron bond was cleaved [42]. Thus, carboxymethylated cyt c reacted with excess H2O2, generating a Compound I-like absorption band at 628 nm. This band was stable for more than 30 min [42]. In dimeric cyt c, the Met80‒heme iron band is perturbed, and absorption bands have been observed at 595 and 650 nm by the reaction with mCPBA, indicating formation of a Compound-I like species [26]. The absorption band around 650 nm, characteristic of Compound I, has also been detected in other heme proteins, such as HRP (~622 and ~650 nm) [46, 47], and a dye-decolorizing peroxidase (613 and 648 nm) [48]. Compound I of cyt c may have been produced by the reaction with mCPBA when cyt c interacts with CL in liposomes. In Y67F cyt c, the Met80‒heme iron band is perturbed and mCPBA may bind to the heme iron and generate Compound I more efficiently than the wild-type protein (Fig. 5C). Oxidative modification of a Trp residue in the heme vicinity by a Compound I-like species 17 .
(18) has been reported for the F43W/H64L mutant of myoglobin by the reaction of its ferric form with mCPBA [49]. The oxy complex of a cyt c mutant with Val83 and Glu84 residues deleted was converted to a Compound I-like species by incubation with DTT in air, where Met80 of the cyt c mutant was oxidized to methionine sulfoxide [31]. These results support the hypothesis that Met80 can be oxidized by a Compound I-like species. Considering these results, we propose the following mechanism for the oxidative modification of Met80 by the reaction of cyt c with peroxides in the presence of CLcontaining liposomes (Fig. 7). By interaction of ferric cyt c with CL, Met80 dissociates from the heme iron, allowing a peroxide, mCPBA or H2O2, to bind to the oxidized heme. For the case of ferrous cyt c interacting with CL, cyt c may be oxidized when Met80 dissociates from the heme owing to the low redox potential of the 5-coordinate heme. After the peroxide binding, Compound I forms and selectively oxidizes the dissociated Met80 near the heme. The present results indicate that Met80 of cyt c is oxidized by peroxides in the presence of CL-containing liposomes. The peroxidase activity of cyt c increases by the oxidation of Met80 [27] or carboxymethylations at Met65 and Met80 [42]. The coordination of Met80 to the heme may become weaker by the oxidative modification of Met80, increasing the peroxidase activity of cyt c. It has been reported that the oxidation of CL by the cyt c peroxidase reaction plays an important role in apoptosis by enhancing the membrane permeability [17]. Since the peroxidase activity increased for Met80-modified cyt c, the Met80 oxidation may increase the apoptotic activity of cyt c. 4. Conclusion According to mass and kinetic measurements, Met80 of cyt c was selectively modified 18 .
(19) by the reaction of ferric cyt c with mCPBA in the presence of CL. Met80 was also selectively modified by the reaction of dimeric cyt c, in which the Met80‒heme iron band is perturbed, with peroxides even in the absence of CL. These results demonstrate that Met80 of cyt c is selectively oxidized by peroxides when Met80 dissociates from the heme iron. Met80-modified cyt c was purified successfully, and its peroxidase activity was shown to increase more than 5-fold compared to that of unmodified cyt c. An absorption band at 650 nm was transiently observed by the reaction of ferric cyt c with mCPBA in the presence of CL-containing DOPC liposomes, but not in the presence of DOPC liposomes not containing CL. These results indicate that Met80 of cyt c is oxidatively modified by peroxides through the formation of a Compound I-like species, whereas the peroxidase activity of the Met-modified cyt c increases compared to that of the native protein, which may increase the apoptotic activity of cyt c.. Table of Abbreviations ABTS,. 2,2'-azino-bis(3-ethylbenzothiazoline-6-sulfonic. acid;. Mm,. calculated. monoisotopic mass; CL, cardiolipin; CD, circular dichroism; cyt c, cytochrome c; DOPC, 1,2-dioloeyl-sn-glycero-3-phosphocholine; ESI-MS, electrospray ionization mass; LCMS/MS, liquid chromatography-tandem mass; MALDI-TOF, matrix-assisted laser desorption ionization-time of flight; mCPBA, meta-chloroperbenzoic acid; MsrA, methionine-S-sulfoxide reductase A.. Acknowledgments We thank Prof. Leigh McDowell, Nara Institute of Science and Technology, for his advice on manuscript preparation. We are also grateful to Ms. Mika Yamamura, Ms. Yuriko 19 .
(20) Nishiyama, and Ms. Yoshiko Nishikawa, Nara Institute of Science and Technology, for ESI-MS and MALDI-TOF mass measurements, and Mr. Jun Watanabe and Ms. Keiko Matsumoto, Shimadzu Cooperation, for LC-MS/MS measurements. This work was partially supported by Grants-in-Aid for Scientific Research (Category B, No. JP26288080, S.H.; Innovative Areas, No. JP16H00839, S.H.) from JSPS.. Appendix A. Supplementary data Supplementary data to this article can be found online at http://xxxxx.. References [1] D. Spierings, G. McStay, M. Saleh, C. Bender, J. Chipuk, U. Maurer, D.R. Green, Science 310 (2005) 66–67. [2] P. Li, D. Nijhawan, I. Budihardjo, S.M. Srinivasula, M. Ahmad, E.S. Alnemri, X. Wang, Cell 91 (1997) 479–489. [3] R.E. Dickerson, T. Takano, D. Eisenberg, O.B. Kallai, L. Samson, A. Cooper, E. Margoliash, J. Biol. Chem. 246 (1971) 1511–1535. [4] G.W. Bushnell, G.V. Louie, G.D. Brayer, J. Mol. Biol. 214 (1990) 585–595. [5] G. Daum, Biochim. Biophys. Acta 822 (1985) 1–42. [6] V.E. Kagan, V.A. Tyurin, J.F. Jiang, Y.Y. Tyurina, V.B. Ritov, A.A. Amoscato, A.N. Osipov, N.A. Belikova, A.A. Kapralov, V. Kini, I.I. Vlasova, Q. Zhao, M.M. Zou, P. Di, D.A. Svistunenko, I.V. Kurnikov, G.G. Borisenko, Nat. Chem. Biol. 1 (2005) 223–232. [7] F. Sinibaldi, L. Fiorucci, A. Patriarca, R. Lauceri, T. Ferri, M. Coletta, R. Santucci, Biochemistry 47 (2008) 6928–6935. 20 .
(21) [8] J. Hanske, J.R. Toffey, A.M. Morenz, A.J. Bonilla, K.H. Schiavoni, E.V. Pletneva, Proc. Natl. Acad. Sci. U.S.A. 109 (2012) 125–130. [9] R. Vazquez-Duhalt, J. Mol. Catal. B: Enzym. 7 (1999) 241–249. [10] R.E. Diederix, M. Ubbink, G.W. Canters, Eur. J. Biochem. 268 (2001) 4207–4216. [11] Y.A. Vladimirov, E.V. Proskurnina, D.Y. Izmailov, A.A. Novikov, A.V. Brusnichkin, A.N. Osipov, V.E. Kagan, Biochemistry (Moscow) 71 (2006) 998– 1005. [12] N.A. Belikova, Y.A. Vladimirov, A.N. Osipov, A.A. Kapralov, V.A. Tyurin, M.V. Potapovich, L.V. Basova, J. Peterson, I.V. Kurnikov, V.E. Kagan, Biochemistry 45 (2006) 4998–5009. [13] J.M. Bradley, G. Silkstone, M.T. Wilson, M.R. Cheesman, J.N. Butt, J. Am. Chem. Soc. 133 (2011) 19676–19679. [14] L.V. Basova, I.V. Kurnikov, L. Wang, V.B. Ritov, N.A. Belikova, I.I. Vlasova, A.A. Pacheco, D.E. Winnica, J. Peterson, H. Bayir, D.H. Waldeck, V.E. Kagan, Biochemistry 46 (2007) 3423–3434. [15] M. Ott, J.D. Robertson, V. Gogvadze, B. Zhivotovsky, S. Orrenius, Proc. Natl. Acad. Sci. USA 99 (2002) 1259–1263. [16] S.L. Iverson, S. Orrenius, Arch. Biochem. Biophys. 423 (2004) 37–46. [17] Y.Y. Tyurina, V. Kini, V.A. Tyurin, I.I. Vlasova, J. Jiang, A.A. Kapralov, N.A. Belikova, J.C. Yalowich, I.V. Kurnikov, V.E. Kagan, Mol. Pharmacol. 70 (2006) 706–717. [18] V.E. Kagan, G.G. Borisenko, Y.Y. Tyurina, V.A. Tyurin, J. Jiang, A.I. Potapovich, V. Kini, A.A. Amoscato, Y. Fujii, Free Radic. Biol. Med. 37 (2004) 1963–1985. [19] A.A. Kapralov, N. Yanamala, Y.Y. Tyurina, L. Castro, A. Samhan-Arias, Y.A. 21 .
(22) Vladimirov, A. Maeda, A.A. Weitz, J. Peterson, D. Mylnikov, V. Demicheli, V. Tortora, J. Klein-Seetharaman, R. Radi, V.E. Kagan, Biochim. Biophys. Acta 1808 (2011) 2147–2155. [20] L.J. McClelland, H.B. Steele, F.G. Whitby, T.C. Mou, D. Holley, J.B. Ross, S.R. Sprang, B.E. Bowler, J. Am. Chem. Soc. 138 (2016) 16770–16778. [21] A.M. Berghuis, J.G. Guillemette, M. Smith, G.D. Brayer, J. Mol. Biol. 235 (1994) 1326–1341. [22] T. Ying, Z.H. Wang, Y.W. Lin, J. Xie, X. Tan, Z.X. Huang, Chem. Commun. (2009) 4512–4514. [23] W. Lan, Z. Wang, Z. Yang, T. Ying, X. Zhang, X. Tan, M. Liu, C. Cao, Z.-X. Huang, PLoS One 9 (2014) e107305. [24] L. Tognaccini, C. Ciaccio, V. D'Oria, M. Cervelli, B.D. Howes, M. Coletta, P. Mariottini, G. Smulevich, L. Fiorucci, J. Inorg. Biochem. 155 (2016) 56–66. [25] S. Hirota, Y. Hattori, S. Nagao, M. Taketa, H. Komori, H. Kamikubo, Z. Wang, I. Takahashi, S. Negi, Y. Sugiura, M. Kataoka, Y. Higuchi, Proc. Natl. Acad. Sci. U.S.A. 107 (2010) 12854–12859. [26] Z. Wang, T. Matsuo, S. Nagao, S. Hirota, Org. Biomol. Chem. 9 (2011) 4766–4769. [27] Y.R. Chen, L.J. Deterding, B.E. Sturgeon, K.B. Tomer, R.P. Mason, J. Biol. Chem. 277 (2002) 29781–29791. [28] H.S. Aluri, D.C. Simpson, J.C. Allegood, Y. Hu, K. Szczepanek, S. Gronert, Q. Chen, E.J. Lesnefsky, Biochim. Biophys. Acta 1840 (2014) 3199–3207. [29] V. Yin, G.S. Shaw, L. Konermann, J. Am. Chem. Soc. 139 (2017) 15701–15709. [30] L.A. Brennan, W. Lee, T. Cowell, F. Giblin, M. Kantorow, Mol. Vis. 15 (2009) 985–999. 22 .
(23) [31] Z. Wang, Y. Ando, A.D. Nugraheni, C. Ren, S. Nagao, S. Hirota, Mol. Biosyst. 10 (2014) 3130–3137. [32] P.P. Parui, M.S. Deshpande, S. Nagao, H. Kamikubo, H. Komori, Y. Higuchi, M. Kataoka, S. Hirota, Biochemistry 52 (2013) 8732–8744. [33] E.A. Berry, B.L. Trumpower, Anal. Biochem. 161 (1987) 1–15. [34] T. Matsui, S. Ozaki, Y. Watanabe, J. Biol. Chem. 272 (1997) 32735–327358. [35] T. Matsui, S. Ozaki, Y. Watanabe, J. Am. Chem. Soc. 121 (1999) 9952–9957. [36] S.M. Kapetanaki, G. Silkstone, I. Husu, U. Liebl, M.T. Wilson, M.H. Vos, Biochemistry 48 (2009) 1613–1619. [37] G. Silkstone, S.M. Kapetanaki, I. Husu, M.H. Vos, M.T. Wilson, Biochemistry 51 (2012) 6760–6766. [38] Y. Lu, D.R. Casimiro, K.L. Bren, J.H. Richards, H.B. Gray, Proc. Natl. Acad. Sci. U.S.A. 90 (1993) 11456–11459. [39] K.L. Bren, H.B. Gray, J. Am. Chem. Soc. 115 (1993) 10382–10383. [40] G.G. Silkstone, C.E. Cooper, D. Svistunenko, M.T. Wilson, J. Am. Chem. Soc. 127 (2005) 92–99. [41] Z.H. Wang, Y.W. Lin, F.I. Rosell, F.Y. Ni, H.J. Lu, P.Y. Yang, X.S. Tan, X.Y. Li, Z.X. Huang, A.G. Mauk, Chembiochem : a European journal of chemical biology 8 (2007) 607–609. [42] S. Prasad, N.C. Maiti, S. Mazumdar, S. Mitra, Biochim. Biophys. Acta 1596 (2002) 63–75. [43] D.A. Capdevila, S. Oviedo Rouco, F. Tomasina, V. Tortora, V. Demicheli, R. Radi, D.H. Murgida, Biochemistry 54 (2015) 7491–7504. [44] M.W. Mara, R.G. Hadt, M.E. Reinhard, T. Kroll, H. Lim, R.W. Hartsock, R. 23 .
(24) Alonso-Mori, M. Chollet, J.M. Glownia, S. Nelson, D. Sokaras, K. Kunnus, K.O. Hodgson, B. Hedman, U. Bergmann, K.J. Gaffney, E.I. Solomon, Science 356 (2017) 1276–1280. [45] A. Lawrence, C.M. Jones, P. Wardman, M.J. Burkitt, J. Biol. Chem. 278 (2003) 29410–29419. [46] W.E. Blumberg, J. Peisach, B.A. Wittenberg, J.B. Wittenberg, J. Biol. Chem. 243 (1968) 1854–1862. [47] H.B. Dunford, J.S. Stillman, Coord. Chem. Rev. 19 (1976) 187–251. [48] J.N. Roberts, R. Singh, J.C. Grigg, M.E. Murphy, T.D. Bugg, L.D. Eltis, Biochemistry 50 (2011) 5108–5119. [49] I. Hara, T. Ueno, S. Ozaki, S. Itoh, K. Lee, N. Ueyama, Y. Watanabe, J. Biol. Chem. 276 (2001) 36067–36070.. 24 .
(25) 779.44 604.35 [56-60] 634.39 [80-86] [9-13] 589.28 [1-5]. 678.37 [74-79]. 1350.72 [89-99] 795.45 [80-86]+16 1296.73 [28-39]. 1470.69 [40-53] 1495.64 [61-72] 1633.61 [14-22]+heme. Intensity. Intensity. a. d. 770. 780. Intensity. b. c 500. 1000. m/z. 1500. e. 795.45. 779.44. m/z. 790. 800. 1495.64. 1490. 1500 m/z. 1510. 1520. 2000. Fig. 1. ESI-MS spectra of the peptide fragments obtained by the digestion of horse cyt c with lysyl endopeptidase after the reaction of its ferric form with mCPBA in the presence of DOPC liposomes (a) with and (b) without CL or (c) in the presence of CL without DOPC. The expanded spectra around m/z = (d) 770‒800 and (e) 1490‒1520 of spectrum (a) are also depicted. The protein solution after the reaction with mCPBA was treated with lysyl endopeptidase at 37 °C for 12 h before mass measurements. The assigned amino acid sequence numbers of the peptide fragments are depicted in brackets. Reaction conditions: cyt c, 0.3 mM; DOPC, 3 mM; CL, 3 mM; mCPBA, 0.4 mM; buffer, 25 mM HEPES buffer; pH, 7.4; temperature, 4 °C.. 25 .
(26) [M (Immonium ion)+O]+ 120.0. G. I [K]+. [IK]+. 260.2 317.0. I. F [AGIK]+. [ FAGIK]+. 388.4. 535.2. [IFAGIK]+ 648.5. Intensity. 147.0. A. [GIK]+. 200. 400 m/z. 600. Fig. 2. Tandem mass spectrum of the protonated molecular ion of the oxygenated peptide fragment with m/z = 795.45 obtained by the digestion of ferric horse cyt c after the reaction with mCPBA in the presence of CL-containing DOPC liposomes. The protein solution after the reaction with mCPBA was treated with lysyl endopeptidase at 37 °C for 12 h before the analysis.. 26 .
(27) 604.34 [56-60]. 634.39 [9-13]. 589.27 [1-5]. 678.37 [74-79]. 779.43 [80-86]. 1470.66 [40-53] 1495.66 [61-72] 1296.71 1350.71 [28-39] [89-99]. 795.44 [80-86]+16. 1633.59 [14-22]+heme. Intensity. Intensity. a. b. 779.43. e. 795.44. 770. 780. 790. 800. Intensity. m/z. c. f. 1495.66. 1490. 1500 m/z. 1510. 1520. d 500. 1000. m/z. 1500. 2000. Fig. 3. ESI-MS spectra of the peptide fragments obtained by the digestion of horse cyt c with lysyl endopeptidase after the reaction of (a,c) its ferric dimer and (b,d) monomer with (a,b) mCPBA and (c,d) H2O2. The expanded spectra around m/z = (e) 770‒800 and (f) 1490‒1520 of spectrum (a) are also depicted. The protein solution after reaction with mCPBA was treated with lysyl endopeptidase at 37 °C for 12 h before the mass measurement. The assigned amino acid sequence numbers of the peptide fragments are depicted in brackets. Reaction conditions: cyt c (heme unit), 0.3 mM; mCPBA, 0.4 mM; H2O2, 0.4 mM; buffer, 25 mM HEPES buffer; pH, 7.4; temperature, 4 °C.. 27 .
(28) Fig. 4. Dependence of peroxidase reaction (ABTS oxidation) rates on [H2O2] for ferric Met80-modified (squares, red) and unmodified cyt c (circles, black). Measurement conditions: protein concentration: cyt c concentration (heme), 1 μM; ABTS concentration, 50 μM; buffer, 50 mM potassium phosphate buffer; pH, 7.0; temperature, 25 °C.. 28 .
(29) Fig. 5. Absorption spectra of (A,B) ferric wild-type horse cyt c observed by the reaction with mCPBA in the presence of DOPC liposomes (A) with and (B) without CL and (C) ferric Y67F horse cyt c observed by the reaction with mCPBA in the absence of liposomes. Difference absorption spectra obtained by subtraction of each absorption spectrum with the spectrum at 5 ms after the mixing are shown in the bottom. Measurement conditions: sample concentrations: cyt c, (A,C) ~50 μM and (B) ~40 μM; mCPBA, 2.5 mM; DOPC, 0.1 mM; CL, 0.1 mM; buffer, 50 mM potassium phosphate buffer; pH, 7.0; temperature, 25 °C; reaction times, 5, 10, 20, 30, 50, 70, and 100 ms.. 29 .
(30) Fig. 6. (A) Time-dependent absorbance changes at 650 nm obtained for the reaction of ferric Y67F horse cyt c with mCPBA without liposomes. The least-square fitted curves according to Eq. 3 are shown as black broken lines. (B) Plot of the formation rate constant of the 650-nm absorption species against mCPBA concentrations. The reaction rate increases as the mCPBA concentration is increased. Measurement conditions: sample concentrations: cyt c, 50 μM; mCPBA, 0.5 (red), 1.0 (orange), 1.5 (green), 2.0 (light blue), and 2.5 mM (dark blue); buffer, 50 mM potassium phosphate buffer; pH, 7.0; temperature, 25 °C.. 30 .
(31) Met. Met S. + CL. S. (in liposome). FeIII. FeIII. His. His Oxidized form (Met dissociation). Oxidized form. Met mCPBA (H2O2). Met. S O FeIV. S O. +・. His Compound I. FeIII His Oxidative modification. Fig. 7. Putative mechanism for the oxidation of Met80 in cyt c by a peroxidase reaction in the presence of CL-containing liposomes.. 31 .
(32) Supplementary Data. Oxidative modification of methionine80 in cytochrome c by reaction with peroxides Ari Dwi Nugraheni, Chunguang Ren, Yorifumi Matsumoto, Satoshi Nagao, Masaru Yamanaka, Shun Hirota Graduate School of Materials Science, Nara Institute of Science and Technology, 8916-5 Takayama, Ikoma, Nara 630-0192, Japan E-mail: [email protected] Contents Table S1. Nucleotide sequences of the primers.. p. S2. Fig. S1.. Absorption spectra of ferric horse cyt c before and after incubation with mCPBA.. p. S3. Fig. S2.. Expected peptide fragments obtained by lysyl endopeptidase digestion of horse cyt c.. p. S4. Fig. S3.. HPLC chromatogram of the peptide fragments of horse cyt c obtained by digestion with lysyl endopeptidase.. p. S5. Fig. S4.. Tandem mass spectrum of the protonated molecular ion of p. S6 the oxygenated m/z = 795.44 peptide fragment obtained by digestion of horse cyt c after the reaction of its dimer with mCPBA.. Fig. S5.. MALDI-TOF mass spectra of ferric Met80-modified and unmodified horse cyt c.. p. S7. Fig. S6.. CD spectra of ferric Met80-modified and unmodified horse cyt c.. p. S8. Fig. S7.. Absorption spectra of ferric wild-type and Y67F horse cyt c.. p. S9. Fig. S8.. MALDI-TOF mass spectra of ferric modified and unmodified horse Y67F cyt c.. p. S10. Fig. S9.. Difference absorption spectrum between the spectra at 50–80 ms and 0–15 ms after mixing the Y67F horse cyt c and mCPBA solutions.. p. S11. S1.
(33) Table S1. Nucleotide sequences of the primers. Primer. Sequencea. #1-F #1-R. ATGGAGTTTTTGGAGAATCCCAAGAA CTCCAAAAACTCCATCAGTGTTTCCT. a. Underlines indicate the nucleotides for the modified amino acid.. S2.
(34) Fig. S1. Absorption spectra of ferric horse cyt c before (black) and after (red) incubation with mCPBA for 60 min. Reaction conditions: cyt c, 0.3 mM; mCPBA, 0.4 mM; buffer, 25 mM HEPES buffer; pH, 7.4; temperature, room temperature.. S3.
(35) GDVEK [1-5]+acetyl 589.28 HK. GK. K. IFVQK [9-13] 634.39. CAQCHTVEK [14-22]+heme 1633.61. TGPNLHGLFGRK. TGQAPGFTYTDANK. [28-39] 1296.72. [40-53] 1470.69. EETLMEYLENPK. K. [61-72] 1495.70 TEREDLIAYLK. YIPGTK. MIFAGIK. [74-79] 678.38. [80-86] 779.45. GGK. NK. GITWK [56-60] 604.35. K. K. ATNE. [89-99] 1350.73 Fig. S2. Expected peptide fragments obtained by lysyl endopeptidase digestion of horse cyt c. The number inside the bracket represents the amino acid sequence number of the peptide, and that under the bracket represents the monoisotopic mass of its protonated molecular ion.. S4.
(36) GDVEK+acetyl. oxidized MIFAGIK TEREDLIAYLK. CAQCHTVEK+heme. TGPNLHGLFGRK EETLMEYLENPK. Intensity. TGQAPGFTYTDANK. MIFAGIK. ATNE. IFVQK GITWK YIPGTK. 0. 5. 10. 15 20 Retention time (min). 25. 30. Fig. S3. HPLC chromatogram of the peptide fragments of horse cyt c obtained by digestion with lysyl endopeptidase after the reaction with mCPBA in the presence of CL-containing DOPC liposomes. The fragments were assigned by their masses, which were defined by MALDI-TOF mass measurements.. S5.
(37) [M (Immonium ion)+O]+ 120.1. I. A. [IK]+ [GIK]+ [AGIK]+ 260.1 317.1 388.2. F. I [FAGIK]+ [IFAGIK]+ 535.3 648.3. Intensity. [K]+ 147.1. G. 200. 400 m/z. 600. Fig. S4. Tandem mass spectrum of the protonated molecular ion of the oxygenated m/z = 795.44 peptide fragment obtained by the digestion of horse cyt c after the reaction of its dimer with mCPBA. The protein solution after reaction with mCPBA was treated with lysyl endopeptidase at 37 °C for 12 h before the mass measurement. Reaction conditions: dimeric cyt c (heme unit), 0.3 mM; mCPBA, 0.4 mM; buffer, 25 mM HEPES buffer; pH, 7.4; temperature, 4 °C.. S6.
(38) Fig. S5. MALDI-TOF mass spectra of ferric (a) Met80-modified and (b) unmodified horse cyt c.. S7.
(39) Fig. S6. CD spectra of ferric Met80-modified (red) and unmodified (black) horse cyt c. Measurement conditions: protein concentration, 8 μM; buffer, 50 mM potassium phosphate buffer; pH, 7.0; temperature, 25 °C.. S8.
(40) Fig. S7. Absorption spectra of ferric wild-type (black) and Y67F (red) horse cyt c. Measurement conditions: protein concentration, 200 μM; buffer, 50 mM potassium phosphate buffer; pH, 7.0; temperature, 25 °C.. S9.
(41) Fig. S8. MALDI-TOF mass spectra of ferric (a) modified and (b) unmodified horse Y67F cyt c. Ferric Y67F cyt c (300 μM) was reacted with mCPBA (600 μM) in 50 mM potassium phosphate buffer, pH 7.0, at 35 °C before the measurement.. S10.
(42) Fig. S9. Difference absorption spectrum obtained by subtraction of the spectrum at 50–80 ms with that at 0–15 ms after mixing the Y67F horse cyt c and mCPBA solutions. Measurement conditions: cyt c, 5 μM; mCPBA, 0.5 mM; buffer, 50 mM potassium phosphate buffer; pH, 7.0; temperature, 25 ºC.. S11.
(43)
図


![Fig. 4. Dependence of peroxidase reaction (ABTS oxidation) rates on [H 2 O 2 ] for ferric Met80-modified (squares, red) and unmodified cyt c (circles, black)](https://thumb-ap.123doks.com/thumbv2/123deta/8635048.1341384/28.892.311.563.127.365/dependence-peroxidase-reaction-oxidation-modified-squares-unmodified-circles.webp)
+7





関連したドキュメント
The C-minor partial orders determined by the clones gen- erated by a semilattice operation (and possibly the constant operations corresponding to its identity or zero elements)
In fact, the only points on H 1 (C) with two preimages on C are the two critical points of the mating. Finally, note that per our construction all deformations of C by H can
The distributed-microstructure model for the flow of single phase fluid in a partially fissured composite medium due to Douglas-Peszy´ nska- Showalter [12] is extended to a
We present sufficient conditions for the existence of solutions to Neu- mann and periodic boundary-value problems for some class of quasilinear ordinary differential equations.. We
The main problem upon which most of the geometric topology is based is that of classifying and comparing the various supplementary structures that can be imposed on a
Let C be a co-accessible category with weak limits, then the objects of the free 1 -exact completion of C are exactly the weakly representable functors from C
The commutative case is treated in chapter I, where we recall the notions of a privileged exponent of a polynomial or a power series with respect to a convenient ordering,
Then it follows immediately from a suitable version of “Hensel’s Lemma” [cf., e.g., the argument of [4], Lemma 2.1] that S may be obtained, as the notation suggests, as the m A
