Introduction
Collembola, which is abundant in soils worldwide, can be found in a diverse range of other habitats including macro-fungal sporocarps (Mateos et al., 1996; Sawahata et al., 2000; Greenslade et al., 2002; Nakamori and Suzuki, 2005; Weiner et al., 2019). In the Ryukyu Islands of Japan, studies on Col-lembola have primarily consisted of species descriptions and faunal surveys (e.g., Yosii, 1964, 1965; Takara and Azuma, 1972; Uchida, 1965; Suma, 2003, 2004; Suma and Karasawa, 2005; Hasegawa et al., 2015). While many studies have been conducted in the mainland Japan (Tsuneda and Arita, 1982; Sawahata et al., 2000; Nakamori and Suzuki, 2005; Sawahata and Narimatsu, 2007; Nakamori et al., 2009; Nakano et al.,
2017), no studies have been conducted on Collembola associ-ated with fungal sporocarps in this region.
Here, we report two species of Ceratophysella associated with fungal sporocarps: the first Japanese record of C. ligula-dorsi (Lee, 1974), which was obtained from macrofungal spo-rocarps on three of the Ryuku Islands: Okinawa, Ishigaki, and Iriomote Islands, and description of C. mediolobata Nakamo-ri, sp. nov., which was obtained from sporocarps on northern Honshu Island, Japan. We determined the sequences of partial regions of the mitochondrial cytochrome c oxidase subunit 1 (COI) and 16S ribosomal RNA (16S) genes of both C. ligula-dorsi and C. mediolobata sp. nov. for DNA barcoding.
A new species of the genus Ceratophysella (Collembola: Hypogastruridae)
and the first record of Ceratophysella liguladorsi from Japan
Taizo Nakamori
1, Kei Ichisawa
2, Hoang Nguyen-Duc Pham
3and Yoshie Terashima
41Graduate School of Environmental and Information Sciences, Yokohama National University, 79-7 Tokiwadai,
Hodoga-ya, Yokohama, Kanagawa, 240-8501 Japan
2Tottori Prefectural Museum, 2-124 Higashimachi, Tottori, Tottori, 680-0011 Japan 3Laboratory of Mycology, Agricultural Hi-tech Park of Ho Chi Minh City, Vietnam
4Center of Molecular Biosciences, Tropical Biosphere Research Center, University of the Ryukyus, Senbaru 1, Nishihara,
Okinawa, 903-0213 Japan
Corresponding author: Taizo Nakamori (nakamori-taizo-gc@ynu.ac.jp) Received 24 August 2018; Accepted 18 May 2019
http://zoobank.org/810DB66C-4D85-4229-ACC7-AAA75EADE16E
Abstract Ceratophysella mediolobata Nakamori, sp. nov., obtained from fungal sporocarps in Akita, northern Japan, is described herein. Ceratophysella liguladorsi (Lee, 1974) has been obtained from fungal sporocarps in the Ryukyus, southern Japan, as the first record from Japan. These species and Ceratophysella tergilobata (Cassagnau, 1954) are similar to each other in having a central projection on abdominal segment V, but C. tergilobata can be distinguished from the other two species by having one sublobal hair on the maxillary outer lobe (two sublobal hairs in the other species), and C. mediolobata sp. nov. can be distinguished from C. liguladorsi by the absence of third setae in the posterior-row on abdominal tergum IV (present in C. liguladorsi). Partial regions of the mito-chondrial cytochorome c oxidase subunit 1 and 16S ribosomal RNA genes of C. liguladorsi and C. mediolobata sp. nov. were sequenced for DNA barcoding.
Abbreviations. a, m, and p-seta(e) ̶ seta(e) of anterior-, middle-, and posterior-row on terga; Abd. I–IV ̶ abdominal segments I–VI; Ant. I–IV ̶ antennal segments I–VI; PAO ̶ postantennal organ; Th. I–III ̶ thoracic segments I–III
Materials and Methods
Collembola were collected from macrofungal sporocarps on Okinawa, Ishigaki, and Iriomote Islands, southern Japan, and fixed in 99% ethanol. For comparison purposes, Col-lembola were also collected from macrofungal sporocarps in Akita Prefecture, northern Japan.
To conduct morphological and molecular analyses of a single specimen, DNA was extracted prior to morphological examinations, as described in Aoyama et al. (2015). In brief, a dissected antenna or the entire body of a specimen was boiled in buffer. For morphological studies, specimens were mounted between cover slips with lactic acid, propylene glycol, or An-dre and Hoyer s fluid and examined under an optical micro-scope. If necessary, specimens were cleared in 10% potassium hydroxide aqueous solution for several minutes before mount-ing.
Since C. liguladorsi, C. mediolobata sp. nov., and C. ter-gilobata (Cassagnau, 1954) are similar to each other, we also examined the morphology of some of the voucher specimens reported under the name C. tergilobata from Hokkaido Island (Suma, 1985, 2017), Okinawa Island (Suma and Karasawa, 2005), and Ishigaki and Iriomote Islands (Suma, 2003), Japan. DNA sequences of partial regions of the COI and 16S genes of representative specimens of C. liguladorsi and C. mediolobata sp. nov. were determined by direct Sanger se -quencing as described in Potapov et al. (2017). Sequences of both genes were aligned using MUSCLE (Edgar, 2004; https://www.ebi.ac.uk/Tools/msa/muscle/), and then 16S se-quences were cleaned using Gblocks 0.91b (Castresana, 2000; http://molevol.cmima.csic.es/castresana/GBLOCKS_server. html). Proportions of different sites between two sequences (p-distances) were calculated using MEGA7 (Kumar et al., 2016) for both genes.
Representative specimens of C. liguladorsi and types of C. mediolobata sp. nov. were deposited in the Tottori Prefectural Museum under numbers TRPM-AAr-0000762– TRPM-AAr-0000769 and TRPM-AAr-0000772–TRPM-AAr-0000779. Sequences obtained through this research were submitted to GenBank under accession numbers LC406886– LC406901 (Table 1).
Taxonomic Account
Ceratophysella liguladorsi (Lee, 1974)
(Figs. 1a–1d)
Syn. Hypogastrura (Ceratophysella) tergilobata: Suma, 2003; Suma and Karasawa, 2005
Hypogastrura (Ceratophysella) tergilobata: Yoshii and Suhardjono, 1989
Materials examined
Voucher Materials: two specimens, Okinawa Island (26.7398˚N, 128.2360˚E), 14 November 2013; two speci-mens, Ishigaki Island (24.3752˚N, 124.1571˚E), 4 Septem-ber 2014; and two specimens, Iriomote Island (24.4003˚N, 123.7969˚E), 2 September 2014. Kept in Tottori Prefectural Museum (voucher numbers TRPM-AAr-0000762–TRPM-AAr-0000767).
Material examined: More than 10 individuals from each voucher material locality.
Description
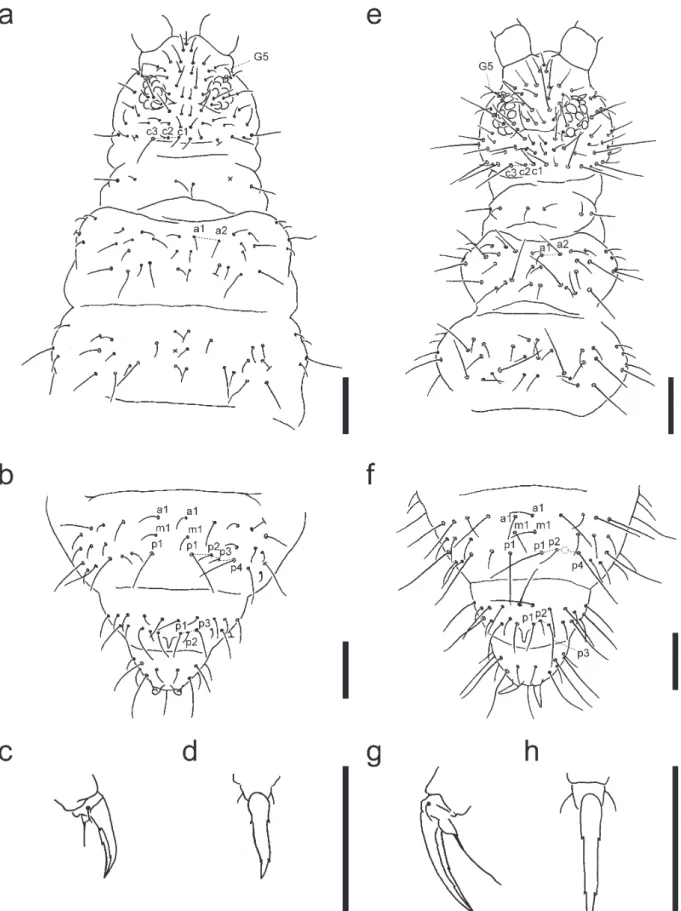
Body length up to 1.5 mm. Color light brown to gray. Tegumentary granulation fine; Abd. V with ca. 10 tegumentary granules between p1-setae. Ant. I and II with 7 and 13 setae, re-spectively. Ant. III organ with two sensilla guarded by a raised cuticle and two guard sensilla. Ant. III microsensilla present. Ant. IV with a trilobed apical antennal bulb and eight poorly dif-ferentiated blunt sensilla; ventral file with numerous (ca. 40–70) peg-like setae surrounding a central seta. Eyes 8 + 8. PAO with four lobes and an accessory tubercle. Labrum with 5, 5, and 4 setae. Maxillary outer lobe with two sublobal hairs. Maxillary head armata-type (Fjellberg, 1984) with lamella 1 pointed and not feathered. Claw with an internal tooth and two lateral teeth (Fig. 1c, 1d). Empodium with a lamella and filament reaching half the length of the claw (Fig. 1c). Ventral tube laterally with 4 + 4 setae. Tenaculum with 4 + 4 teeth. Dens with seven setae on posterior side. Mucro curved, with lateral lamella. Abd. V with a central projection covered with fine granules. Anal spines long, pale yellow in color, situated on basal papillae. Body setae slightly serrated and pointed at the apex. Sensory setae long and weakly differentiated from body setae. G5-setae on head mod-erately long: longer than short setae c2 but shorter than macro-setae c3. Th. I with 3 + 3 macro-setae. Th. II with a2-macro-setae slightly longer than a1-setae (Fig. 1a). One pair of lateral microsensilla present on Th. II (not illustrated). Abd. IV with p1-setae about
2.5 times as long as p2-setae, and p3-setae present (Fig. 1b). Distance between m1-setae subequal or slightly longer than that between a1-setae on Abd. IV. Abd. V with p2-setae situated on the line between p1- and p3-setae (Fig. 1b).
Remarks
Morphological characteristics were generally consistent with the original description of Korean specimens by Lee (1974), except that Japanese specimens with two lateral teeth (one lateral tooth in original description) and G5-setae mod-erately long (G5-setae macrochaeta in original description). Ceratophysella liguladorsi differs from C. mediolobata sp. nov. and C. tergilobata in the relative length of the a2-setae (a2-setae only slightly longer than a1-setae) on Th. II and the presence of p3-setae on Abd. IV.
Ceratophysella liguladorsi has sometimes been reported under the name C. tergilobata. Voucher specimens recorded under the name C. tergilobata from Okinawa Island (Suma and Karasawa, 2005) and Ishigaki and Iriomote Islands (Suma, 2003) were morphologically identical to specimens of C. ligu-ladorsi described in the present study. Specimens recorded as C. tergilobata from Java, Indonesia (Yoshii and Suhardjono, 1989) can be identified as C. liguladorsi by the relative length of a2-setae on Th. II (Fig. 1G of cited article). Therefore, the updated distribution of C. liguladorsi is as follows: Ko-rea (Lee, 1974), China (Weiner et al., 2019), southern Japan (Suma 2003; Suma and Karasawa, 2005; present study), and Indonesia (Yoshii and Suhardjono, 1989). By considering these occurrences of C. liguladorsi in Asia and the
Mediter-ranean distribution of C. tergilobata (Thibaud et al., 2004), the species listed as C. tergilobata in Thailand (Jantarit et al., 2016) seems likely to instead be C. liguladorsi.
Distribution and ecology
Known from Korea, China, southern Japan, and Indone-sia. It can be abundant on macrofungal sporocarps (Weiner et al., 2019; present study).
Ceratophysella mediolobata Nakamori, sp. nov.
(Figs. 1e–1h and 2)
Syn. Hypogastrura (Ceratophysella) tergilobata: Suma, 1985
Ceratophysella liguladorsi: Babenko, 1994 Ceratophysella tergilobata: Suma, 2017
Materials examined
Holotype: A female, Noshiro, Akita, Japan (40.0807˚N, 140.2526˚E), 24 September 2016. Kept in Tottori Prefectural Museum (voucher number TRPM-AAr-0000768).
Paratypes: Nine specimens, same locality and date as holotype. Kept in Tottori Prefectural Museum (voucher num-bers TRPM-AAr-0000769, TRPM-AAr-0000772–TRPM-AAr-0000779).
Description
Body length up to 1.9 mm. Color light brown to gray. Table 1. Voucher sample information and GenBank accession numbers of sequence data used in molecular analyses.
Voucher sample information GenBank accession number
Taxa Locality Individual ID Museum ID COI gene 16S gene
Ceratophysella liguladorsi Okinawa Island OH11 TRPM-AAr-0000762 LC406894 LC406886
OH12 TRPM-AAr-0000763 LC406895 LC406887
Ishigaki Island OH01 TRPM-AAr-0000764 LC406896 LC406888
OH02 TRPM-AAr-0000765 LC406897 LC406889
Iriomote Island OH05 TRPM-AAr-0000766 LC406898 LC406890
OH06 TRPM-AAr-0000767 LC406899 LC406891
Ceratophysella mediolobata sp. nov. Akita FH35 TRPM-AAr-0000768 LC406900 LC406892
Fig. 1. Ceratophysella liguladorsi from Ishigaki Island (a–d) and Ceratophysella mediolobata sp. nov. (e–h). Head, Th. I and II (a, e); Abd. IV–VI (b, f); hind claw (c, d, g, h). Bar, 0.1 mm.
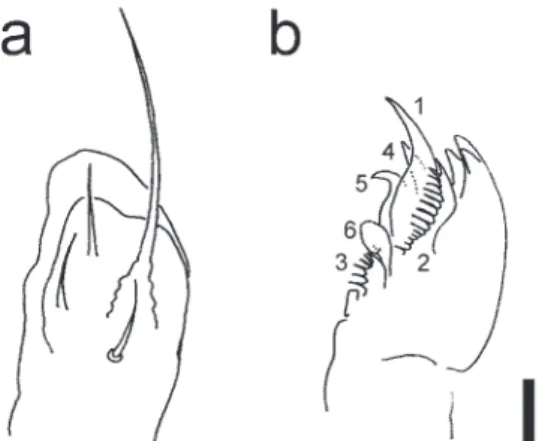
Tegumentary granulation fine; Abd. V with ca. 10 tegumen-tary granules between p1-setae. Ant. I and II with 7 and 13 setae, respectively. Ant. III organ with two sensilla guarded by a raised cuticle and two guard sensilla. Ant. III microsensilla present. Ant. IV with a trilobed apical antennal bulb and eight poorly differentiated blunt sensilla; ventral file with numerous (ca. 40–60) peg-like setae surrounding a central seta. Eyes 8 + 8. PAO with four lobes and an accessory tubercle. Labrum with 5, 5, and 4 setae. Maxillary outer lobe with two sublobal hairs (Fig. 2a). Maxillary head armata-type (Fjellberg, 1984) with lamella 1 pointed and not feathered (Fig. 2b). Claw with an internal tooth and two lateral teeth (Fig. 1h). Empodium with a lamella and filament reaching tip of the claw (Fig. 1g). Ventral tube laterally with 4 + 4 setae. Tenaculum with 4 + 4 teeth. Dens with seven setae on posterior side. Mucro curved, with lateral lamella. Abd V with a central projection covered with fine granules. Anal spines long, pale yellow in color, situ-ated on basal papillae. Body setae slightly serrsitu-ated and point-ed at the apex. Sensory setae long. G5-setae on head moder-ately long: longer than short setae c2 but shorter than macro-setae c3. Th. I with 3 + 3 macro-setae. Th. II with a2-seta 2.5 times as long as a1-setae (Fig. 1e). One pair of lateral microsensilla present on Th. II (not illustrated). Abd. IV with p1-setae about 2.5 times as long as setae p2-setae, and p3-setae absent (Fig. 1f). Distance between m1-setae subequal or slightly longer than that between a1-setae on Abd. IV. Abd. V with p2-setae situated anterior to p1- and p3-setae (Fig. 1f).
Remarks
The new species is characterized by a central projection on Abd. V as in C. liguladorsi and C. tergilobata. In the new species, the tergum chaetotaxy is similar to that of C.
tergilo-bata, and the structures of the maxillary head and maxillary outer lobe are similar to those of C. liguladorsi. The present species, C. mediolobatasp. nov., can be distinguished from C. liguladorsi by the absence of p3-setae on Abd. IV (p3-setae present in C. liguladorsi) and by the relative length of a2-setae̶that is, 2.5 times as long as a1-setae on Th. II (a2-setae only slightly longer than a1-setae in C. liguladorsi). The present species can be distinguished from C. tergilobata by the presence of two sublobal hairs on the maxillary outer lobe (one sublobal hair in C. tergilobata after Fjellberg (1984)) and by having the pointed and non-feathered lamella 1 of the maxillary head (flattened and feathered in C. tergilobata after Fjellberg (1984)).
The new species was previously recorded under differ-ent names. Voucher specimens recorded under the name C. tergilobata from Hokkaido Island (Suma, 1985, 2017) were morphologically identical to the present species. Specimens recorded as C. liguladorsi from Sakhalin, Russia (Babenko, 1994) can be identified as C. mediolobata sp. nov. by the fol-lowing key characters: two sublobal hairs on maxillary outer lobe; pointed and non-feathered lamella 1 of maxillary head; a2-setae 2.5 times as long as a1-setae on Th. II; p3-setae absent on Abd. IV; elongated empodium. Babenko (1994) mentioned the possibility that Sakhalin specimens were a new species. However, Sakhalin specimens had been considered as a variation of C. liguladorsi, since specimens from Vladivo-stok had intermediate characters between C. liguladorsi and Sakhalin specimens (Babenko, 1994). In the present study, we considered that the differences between C. liguladorsi and Akita specimens are sufficient to separate the two species, and established C. mediolobata sp. nov. The taxonomic position of the Vladivostok population remains to be studied.
Distribution and ecology
Known from northern Japan and eastern Russia. It can be abundant on macrofungal sporocarps (Suma, 1985; present study).
Etymology
The specific name mediolobata, from the Latin words medius, middle, and lobus, a lobe, refers to the prominent me -dian projection on Abd. V.
Molecular identification
Molecular data showed that p-distances for COI and 16S genes within C. liguladorsi were 0.002–0.199 and 0.000– 0.126, while those between C. liguladorsi and C. mediolobata sp. nov. were 0.196–0.198 and 0.154–0.168, respectively. The Okinawa, Ishigaki, and Iriomote populations of C. liguladorsi all showed large p-distances with C. mediolobata sp. nov.; within C. liguladorsi, the p-distances were large between the Okinawa and Ishigaki–Iriomote populations (Table 2). Based on these results, it seems that COI and 16S sequences can be used to distinguish C. liguladorsi and C. mediolobata sp. nov. if local populations are considered. Accumulation of barcode data for local populations will improve the identification of these species through DNA barcoding.
Key to species of Ceratophysella with a central projection on Abd. V
1. Maxillary outer lobe with one sublobal hair ……… ………C. tergilobata (Cassagnau, 1954) (Western Europe) - Maxillary outer lobe with two sublobal hairs ………2 2. Abd. IV with p3-setae present……… ………C. liguladorsi (Lee, 1974) (Eastern Asia) - Abd. IV with p3-setae absent………
…………C. mediolobata Nakamori, sp. nov. (Northeastern Asia)
Acknowledgments
We would like to thank Iriomote Station, Tropical Biosphere Research Center and Yona Field, Subtropical Field Science Center, University of the Ryukyus and TOHOKU Regional For-est Office for their support in conducting field surveys. We also thank Mr. Suma for giving us the opportunity to examine his voucher specimens and Ms. Chika Sumida (Yokohama National University, Japan) for supporting the sequence analyses. The study was supported in part by the Collaborative Research of Tropical Biosphere Research Center, University of the Ryukyus, by MEXT/JSPS KAKENHI (Nos. 25281029 and 18H04009), and by JSPS and RFBR under the Japan–Russia Research Coop-erative Program (No. 16-54-50068 in Russian side).
Table 2. Ranges of p-distances for COI and 16S genes of Ceratophysella liguladorsi and Ceratophysella mediolobata sp. nov.
C. liguladorsi C. mediolobata sp. nov.
Okinawa Ishigaki Iriomote Akita
COI gene Okinawa 0.002 Ishigaki 0.184–0.188 0.008 Iriomote 0.190–0.199 0.029–0.032 0.011 Akita 0.196–0.198 0.179–0.181 0.172–0.176 0.005 16S gene Okinawa 0.000 Ishigaki 0.130–0.133 0.002 Iriomote 0.126 0.009–0.012 0.000 Akita 0.154–0.156 0.164–0.168 0.156–0.159 0.002
摘 要
中森泰三1・ 一澤圭2・Hoang Nguyen-Duc Pham3・ 寺嶋芳江4
(1横浜国立大学環境情報研究院〒240-8501神奈川県横浜市保
土ヶ谷区常盤台79-7・2鳥取県立博物館〒680-0011鳥取県鳥
取市東町二丁目124番地・3Laboratory of Mycology, Agricul-tural Hi-tech Park of Ho Chi Minh City, Vietnam・4琉球大学熱
帯生物圏研究センター分子生命科学研究施設〒903-0213沖
縄県中頭郡西原町字千原1番地):Ceratophysella 属(トビム
シ目: ムラサキトビムシ科) の一新種およびCeratophysella
liguladorsiの日本初記録.
秋田県において菌類の子実体から得られたCeratophysella
me-diolobata Nakamori, sp. nov.を記載した.また,沖縄県から本 邦初記録となるCeratophysella liguladorsi (Lee, 1974)が菌類
の子実体から得られた. これらの2種およびCeratophysella tergilobata (Cassagnau, 1954)は腹部第5節に突起をもつ点で 類似するが,C. tergilobataは他の2種と上顎外片に1本の副 片毛をもつ点で区別でき(他の2種は2本),C. mediolobata sp. nov.はC. liguladorsiと腹部第4節背面の後列の第3毛を 欠く点で区別できる(C. liguladorsiは第3毛をもつ).DNA バーコードによる同定を可能にするために,C. mediolobata sp. nov.およびC. liguladorsiのミトコンドリアのチトクローム CオキシダーゼサブユニットI遺伝子および16Sリボソーム RNA遺伝子の一部分の塩基配列を決定した. キーワード: 秋田,DNAバーコーディング,剛毛式,菌類子 実体,琉球列島 References
Aoyama, H., Saitoh, S., Fujii, S., Nagahama, H., Shinzato, N., Kaneko, N., and Nakamori, T., 2015. A rapid method of non-destructive DNA extraction from individual springtails (Col-lembola). Applied Entomology and Zoology, 50: 419–425. Babenko, A. B., 1994. Genus Hypogastrura. pp. 30–195, In:
Col-lembola of Russia and Adjacent Countries: Family Hypogas-truridae (ed. Chernova, N. M.), Nauka, Moscow (in Russian). Cassagnau, P., 1954. Faune française des Collemboles (V). Col-lemboles recoltés en Corse par Miss T. Clay. The Proceedings
of the Royal Entomological Society of London (B), 23: 11–12. Castresana, J., 2000. Selection of conserved blocks from multiple
alignments for their use in phylogenetic analysis. Molecular
Biology and Evolution, 17: 540–552.
Edgar, R. C., 2004. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Research, 32: 1792–1797.
Fjellberg, A., 1984. Maxillary structures in Hypogastruridae (Col-lembola). Annales de la Société Royale Zoologique de
Bel-gique, 114: 89–99.
Greenslade, P., Simpson, J. A. and Grgurinovic, C. A., 2002. Col-lembola associated with fungal fruit-bodies in Australia.
Pe-dobiologia, 46: 345–352.
Hasegawa, M., Sasaki, T., Sato, H. and Abe, S., 2015. Effects of roads on collembolan community structure in subtropical evergreen forests on Okinawa Island, southwestern Japan.
Pedobiologia, 58: 13–21.
Jantarit, S., Bedos, A. and Deharveng, L., 2016. An annotated checklist of the Collembolan fauna of Thailand. Zootaxa, 4169: 301–360.
Kumar, S., Stecher, G. and Tamura, K., 2016. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets.
Molecular Biology and Evolution, 33: 1870–1874.
Lee, B.-H., 1974. Étude de la faune Coréenne des insectes collem-boles II. Description de quatre éspeces nouvelles de la famille Hypogastruridae. Nouvelle Revue d Entomologie, 4: 89–102. Mateos, E., López, R., Barranco, T., Hoyo, P. and Llimona, X.,
1996. Colémbolos (Hexapoda, Collembola) asociados con carpóforos de Basidiomicetes recolectados en el SW de Cata-luña. Revista Catalana de Micología, 19: 99–107.
Nakamori, T. and Suzuki, A., 2005. Preference of three collembo-lan species for fruit-bodies of three species of basidiomycete fungi. Pedobiologia, 49: 119–125.
Nakamori, T., Suzuki, A., Itoh, R. and Hasegawa, M., 2009. First record of Microgastrura Stach (Collembola: Hypogastruri-dae) from Japan, with notes on its DNA barcodes.
Edapholo-gia, 85: 13–17.
Nakano, M., Ochiai, A., Kamata, K. and Nakamori, T., 2017. The preference of Morulina alata (Collembola: Neanuridae) feeding on some fungal sporocarps and the effects of passage through the gut on spores. European Journal of Soil Biology, 81: 116–119.
Potapov, M., Nakamori, T., Saitoh, S., Kuznetsova, N. and Ba-benko, A., 2017. New or little-known taxa of Anurophorinae (Collembola) with anal spines from East Asia with notes on DNA barcode. Zootaxa, 4318: 312–324.
Sawahata, T. and Narimatsu, M., 2007. Abundance of Collembola (Insecta) inhabiting the hyphal mat of an ectomycorrhizal fungus, Sarcodon scabrosus, in a Pinus densiflora forest.
My-coscience, 48: 63–65.
Sawahata, T., Soma, K. and Ohmasa, M., 2000. Number and food habit of springtails on wild mushrooms of three species of Agaricales. Edaphologia, 66: 21–33.
Suma, Y., 1985. Classification and key to families or genera of springtails (Collembola) of Hokkaido (1). Sylvicola, 3: 37–48
(in Japanese; translated English title was given in the present article).
Suma, Y., 2003. On the Collembolan fauna of Okinawa, Part I.
Jezoensis, 29: 81–87 (in Japanese).
Suma, Y., 2004. On the Collembolan fauna of Okinawa, Part II.
Jezoensis, 30: 137–142 (in Japanese).
Suma, Y., 2017. Collembola of Mt. Apoi-dake I. Fauna of the zone of creeping pine and upper mixed forests. Jezoensis, 43: 135–143 (in Japanese).
Suma, Y. and Karasawa, S., 2005. Collembola fauna of Okinawa III. Particularly, Collembola of Asplenium nidus. Jezoensis, 31: 89–104 (in Japanese).
Takara, T. and Azuma, S., 1972. Fauna of the Iriomote Island, Ryukyus. II List of the insects (1) Order Protura, Collembola, Thysanura, Ephemeroptera, Odonata and Plecoptera. The
Sci-ence Bulletin of the Faculty of Agriculture, University of the Ryukyus, 19: 87–97 (in Japanese).
Thibaud, J.-M., Schulz, H.-J. and da Gama Assalino, M. M., 2004. Synopses on Palaearctic Collembola: Hypogastruridae.
Ab-handlungen und Berichte des Naturkundemuseums Görlitz,
75: 1–287.
Tsuneda, A. and Arita, T., 1982. Mycophagous activity of a col-lembolan insect, Hypogastrura reticulata Börner on shiitake bed-logs. Reports of the Tottori Mycological Institute, 20: 70–75.
Uchida, H., 1965. Collembola of the Ryukyus. Kontyu, 33: 85–96. Weiner, W. M., Xie, Z., Li, Y. and Sun, X., 2019. Ceratophysella
species from mushrooms in China (Collembola, Hypogastru-ridae). ZooKeys 822: 67–77.
Yoshii, R. and Suhardjono, Y. R., 1989. Notes on the collembolan fauna of Indonesia and its vicinities I. Miscellaneous notes, with special references to Seirini and Lepidocyrtini. Acta
Zoologica Asiae Orientalis, 1: 23–90.
Yosii, R., 1964. Some Collembola from Okinawa caves, with notes on Sinella–Coecobrya complex of Japan. Bulletin of the
Akiyoshi-dai Science Museum, 3: 25–34.
Yosii, R., 1965. On some Collembola of Japan and adjacent coun-tries. Contributions from the Biological Laboratory Kyoto