Mem. School. B. O. S. T. Kinki University No. 18: 1 '"'-' 7 (2006)
Expression and Purification of the Recombinant Mature Bone Morphogenetic Protein-2 of Pinctada fucata in Escherichia coli and Comparison of the Codon Usage
Tomoyuki Miyashita1, Hirotaka Kat01 and Ryousuke Takagi1
Abstract
1
The DNA fragment encoding the mature fragment of the bone morphogenetic protein 2(BMP-2) of Pinctada fucata was cloned into the pGEX-4T-2 expression vector and the expression was investigated. However, the plasmid failed to express the recombinant GST fusion mature protein. The reason of the absence of the expression of the fusion protein may be the existence of the co dons AGA, AGG and CGG for Arg (R) that are the major codons in the N terminal region of the mature protein. These codons are used at a very low frequency in Escherichia. coli (E.
coli )genes. We excluded the DNA region that encodes the amino terminal region of the mature protein, which probably does not participate in the function of the protein, recloned the manipulated cDNA into the pGEX-4T-2 vector and reexamined the expression. The lO-kDa mature fragment of BMP-2 was highly expressed as a glutathione-S-transferase (GST) fusion protein in E. coli BL2l after IPTG induction. GST fusion proteins were purified by using Glutathione Sepharose 4B affinity chromatography. To separate GST and mature BMP-2, the fusion protein bound to the gel was treated by thrombin and the mature BMP-2 was purified by the elution. The production and purification of the mature Pinctadafucata BMP-2 are reported here.
1. Introduction
Bone morphogenetic proteins (BMPs) belong to the transforming growth factor type beta (TGP -~) supergene familyCl). The TGP-~ supergene family plays essential roles in several important processes of living things, such as cell growth, controlling morphogenesis and cell specialization(2). The BMP family consists of at least 14 members.
BMP-2/4 genes play an important role in the ectodermal dorsoventral axis establishment in vertebrates(3,4). BMPs generally have an ability to induce endochondral bone formation at ectopic sites. Especially, BMP-2, -3, -4, -6, and -7 have the action which strongly promotes either chondrogenesis or bone formation(5,6). BMP-2 is one of the main representatives. BMPs are synthesized as precursor proteins. After the translation, these proteins are proteolytically cleaved to produce the mature C-terminal proteins(7). A homodimer of two mature fragments acts as a biologically active ligand and exerts morphogenesis and bone formation via binding to two types of serine/threonine kinase receptors (type I and type II receptors) like other members of the TGP-superfamily. Both receptors are essential for signal transduction. The type I receptors activate intracellular substrate Smad proteins. R-Smads activated by type I receptors form complexes with Co-Smads, and then translocate into the nucleus. TheSmad heteromers bind to DNA directly and activate transcription of target genes via interaction with the other DNA-binding proteins(8,9, 10).
Thus, a series of flows of bone formation is induced.
Growth factors that belong to the TGP -~ superfamily are involved in the regulation of cell proliferation and differentiation in the shells of mollusks(ll, 12, 13). Hard tissue in invertebrates is equivalent to bone in vertebrates. In invertebrates, it could be expected that BMP-2 would participate not only in morphogenesis during development, but also in the formation of hard tissues, like the nacreous or prismatic layer of Pinctadafucata. Since the mature portion that corresponds to the C-terminal 149-amino acid of Pinctada fucata BMP-2 shows 66% identity to the human BMP_2(18), it is likely that BMP-2 of Pinctadafucata has the same functions as BMP-2 in vertebrates that induces the bone formation.
The previous report demonstrated that vertebrate mature BMP-2 expressed in E. coli possesses the capability of inducing osteoblast differentiation and bone formation in vitro and in vivdI4). The synthetic oligopeptide
Received 19 June 2006
This study was supported by the Project Research of the School of Biology Oriented Science and Technology No. 03-IV-13, 2004.
1. Department of Genetic Engineering, Kinki University, Kinokawa, Wakayama 649-6493, Japan
NSVNSKlPKACCVPTELSAI derived from the mature human BMP-2 bound to a BMP-2- specific receptor and elevated the mRNA synthesis of both genes of alkaline phosphatase and osteocalcin in the murine multipotent mesenchymal cell line C3H10T1I2(l5). Moreover, the same oligopeptide also induced ectopic bone formation when conjugated to a covalently cross-linked alginate gel(l6). To examine whether the Pinctadafucata BMP-2 expressed in E. coli induces mesenchymal stem cells to be osteogenic cell lines, we performed the production and purification of the mature Pinctadafucata BMP-2.
2. Materials and Methods 2.1 Bacterial strains and plasmids
The host E.coli strain BL21 (DE3) pLysS (recD::Tn10, F, hsdS, gal, OmpT) was purchased from Stratagene.
The plasmid pGEX4T-2 was purchased from Pharmacia Inc. (Dorval, Quebec, Canada).
2.2 Plasmid constructions
The Pincda fucata BMP-2 gene encoding the mature region was cut out with HincH and Not! from the cDNA cloned into pBluescript SKH(+) vector(I7, 18) (FigJ). The fragment was ligated into the HindI-Not! site of pGEX4 T-2(Novagene).
2.3 SDS-PAGE analysis of the expression of mature BMP-2 in E. coli
pGEX4T-2 carrying the BMP-2 gene encoding the mature region was introduced into E. coli BL21 (DE3) pLysS.
Cells were grown at 37°C in 3 ml Luria broth medium containing ampicillin (100 ~g/ml) to an A600 of about 0.5.
Then, isopropyl-D-thiogalactoside (lPTG) was added to a final concentration of 1 mM, and the cultures were incubated at 25°C. After induction, the 200 ~l of sample was collected five times in six hours and cells were precipitated by centrifugation at 12 000 x g for 2 min. The pellet was dissolved in 10 ~l 1 x SDS sample buffer (50 mM Tris-HCI, pH 6.8, 2% SDS, 0.1 M DTT, 10% glycerol, 0.1 % bromophenol blue). Dissolved samples were heated at 100°C for 3 min, and electrophoresed on 12% SDS-polyacrylamide gel (SDS-PAGE). SDS-PAGE was carried out as described by Laemmli(l9).
2.4 Expression and purification of mature BMP-2 in E. coli
A single colony ofBL21 containing pGEX4T-2 plasmid was grown overnight at 30°C in LB medium containing 1 00 ~g / ml ampicillin. Cultures were then diluted (1: 10 ) in fresh 50 ml LB medium and grown at 25°C to an A600 of approximately 0.5. Then, isopropyl-D-thiogalactoside (lPTG) was added to a final concentration of 1 mM, and the cultures were incubated at 25°C. The cultures were grown for an additional 4 hr. The culture was collected and centrifuged at 12 000 x g for 10 min, and the pellet was dissolved in 20 ml of sonication buffer [50 mM Tris-HCI (pH 8.0), 50 mM NaCI, 1 mM EDTA,l mM DTT]. The cell suspension was sonicated for 20 seconds at O°C in a sonicator (Ultrasonic Generator U0300FB). This processing was repeated six times. After that, Triton X-100 was added to the sonicated sample to be 1 % in the final concentration. The cell extracts were centrifuged at 12000 x g for 40 min and about 20 ml of the supernatant were saved:
Affinity purification was basically performed according to the instructions as described for GST purification column (Pharmacia, USA). The glutathione sepharose was washed with PBS (Phosphate Buffered Saline) and equilibrated with PBS-0.5% Triton X-100 (PBST). The 20 ml of the supernatant of the cell extract was mixed with 1 ml of the glutathione sepharose and incubated at 4°C for 1 hr with gentle rotation. After that, the mixture was centrifuged at 6000 x g for 5 min at 4°C. The supernatant was wasted. The pellet of the glutathione sepharose was washed 5 times with 20 ml of the PBST at 6000 x g for 30 min at 4°C. To elute the fusion protein, 1 ml of the glutathione buffer [50 mM Tris-HCI (PH 9.6), 1 mM DTT, 10 mM reduced glutathione] was added to the glutathione sepharose and incubated for 10 min at room temperature. The supernatant containing the fusion protein was recovered by centrifugation at 6000 x g for 5 min at 4°C.
Thrombin treatment was performed as follows:Fractions containing purified fusion protein were pooled and dialyzed for sonication buffer to remove reduced glutathione. One ml of the glutathione sepharose equilibrated with sonication buffer was added to the dialyzed sample and incubated at 4°C for 2 hr with gentle rotation. After centrifugation at 6000 x g for 5 min at 4°C, the supernatant was wasted. The glutathione sepharose was washed 3
3
times with 20 ml of the thrombin buffer [50 mM Tris-HCl (pH 8.0),150 mM NaCl, 2.5 mM CaCh, 5 mM MgCh, 1 mM DTT] at 6000 x g for 30 min at 4°C. After that, 1 ml of the thrombin buffer containing thrombin(lO units/ml) was added to the glutathione sepharose and incubated for overnight at 4°C with gentle rotation. The glutathione sepharose adsorbed glutathione-S-transferase and released mature BMP-2 were separated by centrifugation at 6000 x g for 30 min at 4°C. The supernatnant containing the mature BMP-2 was concentrated with Vivapore 5 (VIVA SCIENCE).
3o Results and Discussion
It has been certified that the bone morphogenetic protein induces the bone formation in various experimental systems in in vivo and in vitro. The cDNAs of human BMP-2 and BMP-4 were cloned into eukaryotic expression vectors that were transfected into the murine mesenchymal progenitor cell line C3HIOT1I2. The expression of these genes induce the differentiation into the osteogenic lineage(ll). The differentiation ofC3HIOT1I2 cells was confirmed by the appearance of the marker genes typical or specific for osteogenesis such as alkaline phosphatase, osteopontin, osteoca1cin, and osteonectin. It has been demonstrated that recombinant mature human BMP-2 produced in E. eoli is biologically active for the function that it induces the formation of bone or cartilage in vitro and in vivO<20).
100 bp
TATACAGACGACAATC CGGTCGAAGAACTAAAAGAtAAACAoAoTTAoioc"GA"ioAAAoGACoGAT
287aa. Y T O O N R G
R I R
T KRI r
T I N 0 K 0 0 ,oooAAAAoGAAoGoAAA"T°xoGAoioAAAoGoGAGoGAGGoCoGoC;ioiioGAoAOAAA T AGA C GA C GAAAAAA T AA GA G Gl
307aa K R R N R K R R R R R K N R R R K N K R
!
AAAAATAAGAAGAATAGAAAAAATAATAAAACAAAAAGAAAGAAGTACACCGATGCGTGTl
327 aa K N K K N R K N N K T K R K K Y T 0 A C iflinell ~
AAAAGAAAACCATTATATGT1GACTTCAAAGCAGTGGGGTGGAATGACTGGATTTTTGCA!
347aa K R K P L Y V 0 F K A V G W NOW I F A !
CCTCCTGGATACGAAGCTTATTATTGTCATGGGTCATGTAACTGGCCGTATGACGATCAT~
367 aa P P G Y E A Y Y C H G S C N W P Y D O H
i
ATGAACGTCACAAACCATGCAATAGTGCAGGACTTAGTGAATTCTATAAACCCAGGGTCA!
387aa M N V T N H A I V Q 0 L V N S I N P G 5
!
- i
GTACCCAAACCTTGCTGTGTACCCACAGAACTTAGCTCTCTTTCATTGTTATATACCGAC!
407 aai:
o
V P K P C C V P T E L S S L S L L Y T 0
j
GAACATGAAGTTGTCGTCTTAAAAGTTTATCCCGATATGGTTGTTGAAGGATGCGGATGTl
: :I. .. i.~.~.~~.~.: ... ~ ... ~ ... ~ ... :., .... : ... ~ .... _~ ... ~ ... : ... ~ ... _~ ... ~_ .... ~ ... : ... :_ .... ~ ... : ... ...i
Fig. 1 The nucleotide and amino acid sequence of the Pinetada jileata BMP-2 gene corresponding to the mature region. Arg-X-X-Arg putative maturation site is enclosed with solid line and the generated mature protein is enclosed with dashed line. Numbering of amino acids starts from the first ATG of BMP-2 coding sequence. The hatched box shows the multi cloning sites ofpBluescript SK vector. The closed box shows the cDNA of Pinetadafueata BMP-2.
The open box shows the UTR (Untranslated Region). The part of the bold under line shows the array that is similar to the the array SVPKPCCVPTELSSL.
To examine the above-mentioned function about the bone morphogenetic protein-2 (BMP-2) of Pinctadafucata, the expression of the mature protein was examined in E. coli. Fig. I shows the amino acid and the DNA sequence of Pinctada fucata BMP-2 cDNN I7. 18). The Arg-X-X-Arg (R-X-X-R) sequence is a putative processing site to produce the mature protein of TGF-~ family(11. 21). The sequence R-T-K-R enclosed with solid line is corresponds to the maturation site of Pinctada./itcata BMP-2. We cloned the Accll-Notl fragment which contained the complete mature coding region into Smal-NotI site of pGEX4T-2 that was transfected into BL21 E. coli host. After IPTG induction, expression of fusion protein was analyzed with SDS-PAGE. The molecular size of the putative fusion protein is about 45 kDa. However, as shown in Fig. 2, a 25 kDa protein was accumulated after IPTG induction. The molecular size was equivalent to the glutathione-S-transferase. One of the explanations of this phenomenon is the premature translational termination originated in the existence of the codon with extremely low frequency of use.
kDa
M
12 3 4 5 6
225 150 ~ ...
100
75 ...50'"
35~
25~
15
~10~
Fig. 2 SDS-PAGE analysis of total crude protein produce by E. coli. Protein molecular size marker (lane M); 200 /-ll of aliquot of culture lysate before IPTG induction (lane I) and 200 /-ll of aliquot of culture lysate of I h (lane 2), 2h (lane 3), 3h (lane 4), 5h (lane 5) and 6 h (lane 6) after IPTG induction. The arrow indicates the recombinant GST-BMP fusion protein.
Most amino acids are encoded by more than one codon. There is an original tendency about the codon usage efficiency in each organism. That is an own bias in the usage of the 61 available amino acid codons. In each organism, there are strong positive correlation between the tRNA abundance and the codon usage efficiencies(23). Insufficient tRNA pools can lead to translational posing and premature translation termination(24). Therefore, as for over-expression of heterogeneous gene in E. coli, it is necessary to consider the differences in codon usage that impede translational efficiency.
The precise receptor-binding domain of the mature region that induces the bone formation has not yet been clarified. Recently, it has been shown that a synthetic peptide NSVNSKIPKACCYPTELSAI corresponding to residues 68-87 that exist in the C-terminal portion of the human mature BMP-2 protein can induces bone formation(IS). The synthetic peptide KIPKASSYPTELSAISTLY corresponding to residues 73-92 except that Cys78, Cys79, and Met89 are changed to Ser, Ser, and Thr, respectively, also has a similar activity<25). The synthetic peptide induced a higher alkaline phosphatase (ALP) activity than the NSVNSK1PKACCYPTELSAI corresponding to residues 68 -87 peptide. In the murine fibroblast cell line C3H lOT] 12, the 73 -92 peptide had the cell surface receptor binding capability, and elevated the level of the mRNA of osteocalcin gene(26). Therefore, a common part of
5
two arrays that is KIPKACCYPTELSAI will have the function. As the array that is similar to the array SVPKPCCYPTELSSL exists also in the C-terminal portion of Pinctadafucata mature BMP-2 protein (Fig. 1), it is predicted that the mature protein expressed in E. coli that lacks the N terminal will have the function.
Figure 3 shows a comparison of the codon usage frequency between Pinctada jilcata BMP-2 gene and E. coli genes. The codon AGG or AGA for arginine (R) is very low frequency codon in E. coli. High frequency codon CGC for R in E. coli is hardly used in Pinctada jilcala BMP-2 gene.
100
50 0J
11) "" co
<1' M
r"I r"I
1.0 <1'
N 01
0 G c; G G A i'-. C C "
,:,o,lon C C C C G G G G 4~
A C G T A G Po. C G
I
I I Ala I II I
I I ! Arg I IC G T
I")
00 \0
M ("l U'l
r-Ul U'lr-
~ ~ C"J
A A
A A
C T
r- "I' I")
G G
A A
C T
r- 01
U'l U'l
G T C
G T T
(f)
\D
o o
r l
co
o 0
T T T
;.. A G
A G A
C A A
C G
A. A
G Po.
r-.
rl r l M
G A G
!I~~~II
End III~~
Fig. 3 Comparison of codon usage frequency in Pinclada fucata BMP-2 gene and general E. coli genes. The codon usage frequency was shown by percent. A gray bar chart is Pinctada fucata, and the black bar chart is E. coli.
A
B
kDa M 1 2 3 kDa M 1 2 3
225 ~
Hi t
150 100~ 100
75 ~ 75 50 ~
50 ~ 35~ ... 1
35~
~1 ' ... 2
25~
15 "
... 3
15~
10~
Fig. 4 SDS-PAGE analysis of total crude protein produce by E. coLi.
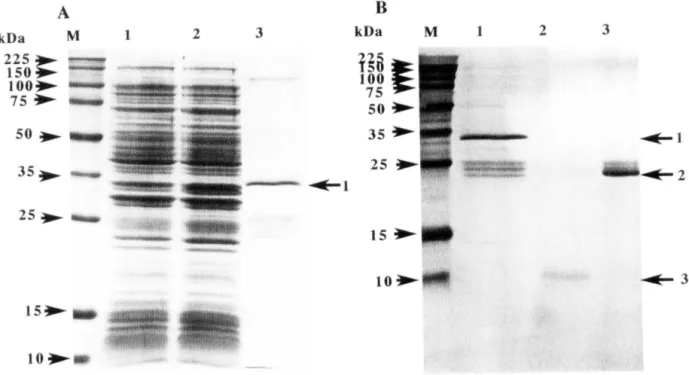
A. lane M: Protein molecular size marker; lane 1:200 /-ll of culture lysate before IPTG induction; lane 2: 200 /-ll of culture lysate of 4h after IPTG induction; lane 3: Affinity-purified GST-BMP fusion protein. The arrow I indicates the recombinant GST-BMP fusion protein. B. lane M: Protein molecular size marker; lane I: Affinity purified GST-BMP fusion protein (arrow I); lane 2: Purified mature BMP-2 protein after thrombin treatment (arrow 3); lane 3: Purified GST protein after thrombin treatment (arrow 2).
As shown in Fig. 1, in the amino acid sequence of the mature amino terminal region, there are two parts where several arginines (R) exist continuously. The codon for R is almost AGG or AGA. These co dons for R are very low frequency codon in E. coli. High frequency codon CGC for R in E. coli is not used in the mature region of Pinctada fucata BMP-2 gene. Therefore, to exclude the cDNA region that encodes a row of arginines that correspond to the N terminal portion of the mature Pinctada fucata BMP-2, HincH-Not! fragment that encodes two third of the mature protein was cut out from the cDNA (Fig. 1 ). The fragment from the manipulated cDNA was recloned into the pGEX-4T-2 vector and the expression was reexamined. The 10 kDa carboxyl-terminal mature fragment of BMP-2 was highly expressed as a glutathione-S-transferase(GST) fusion protein in E. coli BL21 after IPTG induction (Fig.
4A). Molecular size of BMP-GST fusion protein is about 35 kDa. The fusion protein was purified by using Glutathione Sephaiose 4B affinity chromatography. To separate GST and mature BMP-2, the fusion protein bound to the gel was treated with thrombin and the mature BMP-2 was purified by elution as described in Materials and Methods (Fig. 4B).
Concerning the function of the Pinctada fucata BMP-2 expressed in E. coli, it will be necessary to examine whether or not it induces mesenchymal stem cells to be osteogenic cell lines such as osteoblasts and bone cells.
4. References
(1) Wozney, J.M., Rosen,
v.,
Celeste, AJ., Mitsock, L.M., Whitters, M.I, Kriz, RW., Hewick, R.M., and Wang, B.A.(1988) Novel regulators of bone formation: molecular clones and activities. Science. 242, pp. 1528-1534.
(2) Massague, 1 (1998) TGF-p signal transduction. Ann. Rev. Biochem. 67. pp. 753-791.
(3) Harland, R.M. (1994) The transforming growth factor beta family and induction of the vertebrate mesoderm:bone morphogenetic proteins are ventral inducers. Proc Natl Acad Sci USA. 91, pp. 10260- 10264.
(4) Groeneveld, E.H.I, and Burger, E.H. (2000) Bone morphogenetic proteins in human bone regeneration.
European Journal of Endocrinology 142, pp. 9-21.
(5) Yamaguchi, A., Komori, T., and Suda, T. (2000) Regulation of Osteoblast Differentiation Mediated by Bone Morphogenetic Proteins, Hedgehogs, and Cbfa1. Endocrine Reviews 21, pp. 393-411.
(6) Rosen,
v.,
and Wozney, 1M. (2002) Bone morphogenetic proteins. In:Principles of Bone Biology (eds IP.Bilezikian, L.G Raisz, GA. Rodan, et aI.), 2nd edn, pp. 919-928. San Diego: Academic Press
(7) Dubois, C.M., Blanchette, F., Laprise, M.H., Leduc. R., Grondin. F., and Seidah, N.G (2001) Evidence that furin is an authentic transforming growth actor-betal-converting enzyme. Am J Pathoi. 158, pp. 305-316.
(8) Miyazawa, K., Shinozaki, M., Hara. T., Furuya, T., Miyazono. K., Rosen,
v.,
and Wozney, 1M. (2002) Two major Smad pathways in TGF-beta superfamily signalling. Genes Cells. 7, pp. 1191-1204.(9) Moustakas, A., Souchelnytskyi, S., and Heldin, C.H. (2001) Smad regulation in TGF-beta signal transduction. J Cell Sci. 114, pp~ 4359-4369.
(10) Whitman, M. (1998) Smads and early developmental signaling by the TGF beta superfamily. Genes Dev. 12, pp. 2445-2462.
(11) Lelong, C., Mathieu, M., and Favrel, P. (2000) Structure and expression of mGDF, a new member of the transforming growth factor-beta superfamily in the bivalve mollusc Crassostrea gigas. Eur Biochem. 267, pp.3986-93.
(12) Nederbragt, A.I, van Loon, A.E., and Dictus, W.J. (2002) Expression of Patella vulgata orthologs of engrailed and dpp-BMP2/4 in adjacent domains during molluscan shell development suggests a conserved compartment boundary mechanism. Dev BioI. 246, pp. 341-355.
(13) Herpin, A., Lelong, C., Becker, T., Rosa, F., Favrel, P., and Cunningham C. (2005) Structural and functional evidence for a singular repertoire of BMP receptor signal transducing proteins in the lophotrochozoan Crassostrea gigas suggests a shared ancestral BMP/activin pathway. FEBS J. 272, pp.3424-3440.
(14) Ahrens, M., Ankenbauer, T., Schroder, D., Hollnagel, A., Mayer, H., and Gross, G (1993) Expression of human bone morphogenetic proteins-2 or -4 in murine mesenchymal progenitor C3HI0T1I2 cells induces differentiation into distinct mesenchymal cell lineages. DNA Cell BioI. 1, pp. 871-880.
7
(15) Suzuki, Y, Tanihara, M., Suzuki, K., Saitou, A., and Nishimura, Y(2000) Alginate hydrogel linked with synthetic oligopeptide derived from BMP-2 allows ectopic osteoinduction in vivo. J Biomed Mater Res 50, pp.
405- 409.
(16) Saito, A., Suzuki, Y, Ogata, S., Ohtsuki, C., and Tanihara, M. (2005) Accelerated bone repair with the use of a synthetic BMP-2-derived peptide and bone-marrow stromal cells. J Biomed Mater Res A. 72. pp. 77-82.
(17) DDBJ/EMBL/GenBank data base (Accession no. AB 176952).
(18) T. Miyashita, T., Hanashita, T., Toriyama, M., Takagi, R., Akashika, T., and Higashikubo, N., Cloning and Characterization of Pinctadafucata Bone Morphogenetic Protein-2 (BMP-2) gene. (Submitted)
(19)Laemmli, U.K., (1970). Cleavage of structural proteins during the assembly of the head of bacteriophage T4 Nature 227, pp. 680-685.
(20) Kubler, N.R., Reuther, 1.E, Faller, G., Kirchner, T., Ruppert, R., and Sebald, W. (1998) Inductive properties of recombinant human BMP-2 produced in a bacterial expression system. Int J Oral Maxillofac Surg.
27, pp. 305-309.
(21)Derynck, R., Jarrett, 1.A., Chen, E.Y, and Goeddel, D.V (1986) The murine transforming growth factor-beta precursor. 1. BioI. Chern. 261, pp. 4377-4379.
(22) Cui, Y, Jean, E, Thomas, G., and Chrisian, 1.L. (1998) BMP-4 is proteolytic ally activated by furin and/or PC6 during vertebrate embryonic development. EMBO 1. 17, pp. 4735-4743.
(23) Ikemura, T. (1981) Correlation between the abundance of Escherichia coli.transfer RNAs and the occurrence of the respective co dons in its protein genes. 1. Mol. BioI. 146, pp.1-21.
(24)Kurland, C., and Gallant, J., Errors of heterologous protein expression (1996) Curro Opin. BiotechnoI. 7, pp.
489-493.
(25) Saito, A., Suzuki, Y, Ogata, S., Ohtsuki, C., and Tanihara, M. (2003 ) Activation of osteo-progenitor cells by a novel synthetic peptide derived from the bone morphogenetic protein-2 knuckle epitope.Biochim Biophys Acta.
1651, pp. 60-67.
(26) Saito, A., Suzuki, Y, Ogata, S., Ohtsuki, C., and Tanihara, M. (2004) Prolonged ectopic calcification induced by BMP-2-derived synthetic peptide. J Biomed Mater Res A. 70, pp. 115-121.
7"::r -y ffit it%pjGlZ9-f-
2 O)pjG~~~~:r.ijJG~{*1f:::r ~FT 0
cDNA 1f:3Bm~:7 ?r ~pGEX4T-2 ~~:7 p ~==-/
:~lL,
*~~li BL21 C'~'lv?r
77t/ r:7 /
A 7:r: :7~ii ~ O)r;j~~?r/
/'~:7W
~L
-C3Bm1f:~J;.t~7Y~,pjG:9J
Lftip o t~o r;j~~?r/
/'~:7 wtJ~3Bm Lftv \O)~'ipjG~~?r/
/'~:7WO)
N *trffiHf~:5t~~7" lV::Y==-
/tJ~~*j(1ffrL
-C~ ~ ,;cO)~pt::r
F/C'lb0
AGA, AGG ~J:lJ CGG tJ~*~~liC'~'ijitt'{~ffl ~~1Jtv\:' ~ ~~~IZ9T0 ~~*-Gn 0
07" lv::Y==-
/tJ~~*j(1ffrT 0if~:5t~'i:m~ftt~l~!g~~:r.ijJGC'~'iftv\ ~ T~~~n0 t~CJ6, cDNA ~~:r.ijJGtJ~G
;cO)if~:5t1f:!x.'1t-~
it,
3Bm~:7?r ~pGEX4T-2 ~~:7 p ~==- /~' L -c¥.}oc*~~li BL21 C'3Bm1f:1T0
t~o;c
0)fffl* ,
IPTG ~j]D~~~~J: ~ T~~~n0*~ ~ 35 kDa O)r;j~~?r //'~:7WtJ~*~~liNC'*~~~~pjG~nt~o r;j~~?r
/
/~:7 W
~'i ~'lv?r77t
/ t 7 7 p ~ A 4B ~~ J:0
~fo'~~:7 P yr :7":7
7 -1 ~ C'~~L
t~o ~G
~~,W
~~ ~~r;j~~?r
/
/~:7 W1f:fffl~~itkt'\~~C'r
P / ~"/1f:~j]DL,
~'lv?r77t/ r:7 /
A 7:r: :7~ii ~ pjG~~ BMP-2?r / /~:7 W1f:tJJ~