Mem. School.. B. O. S. T. Kinki University No. 10 : 29 ~ 37 (2002)
The steady-state and the pre-steady-state kinetic analysis of the hydrolysis of
N-a -Cbz-L-Lysine p-nitrophenyl
ester catalysed by actinidain isozymes
Koichi Morimoto, Masaya Tamura, and Ben'ichiro Tonomura Summary
29
The steady-state and the pre-steady-state kinetics of two actinidain isozymes, AC-1 and AC-2, were analyzed by stopped-flow method with N-alpha-carbo-benzoxy-L- lysine p-nitrophenyl ester (Z-Lys-ONp) as substrate. These isozymes were purified from kiwifruit (Actinidia deliciosa) by anion-exchange chromatography. The steady-state kinetic parameters, Km and
v,
were determined. Rapid bursts observed in the pre-steady-state for two isozymes are consistent with a three-step mechanism involving an acyl-enzyme intermediate. The values of kca/Km and k+/KmacYl ratios for AC-l were estimated 1.83 and 8.63 pM-1 • S-l respectively, and were two times greater than those for AC-2 (kca/Km = 1.02, k+/K m acyl = 4.50 pM-1 • S-l). For AC-1 and AC-2, the de acylation step is rate-limiting in catalysis, so kcat is similar to k+3 under the experimental conditions.Introduction
A cystein proteinase actinidain [EC 3.4.22.14J is abundant in fruit of kiwifruit (Actinidia deliciosa) (1). The enzymatic properties of actinidain have been studied (2-8), the amino acid sequence of actinidain was first determined by Carne and Moore (9), and cDNAs encoding actinidain were isolated from kiwifruit (8,10). The three dimensional structure of actinidain has been solved at 1.7
A
resolution by X-ray crystallography (11). Though McDowall (2) purified two isozymes of actinidain by ion-exchange chromatography on DEAE-Sephadex A-25, their enzymatic properties could not be well resolved. Sugiyama et al. (12) suggested that kiwifruit contained six proteinase isozymes using two ion-exchange resins under acidic conditions.Kinetic studies on actinidain catalysed-hydrolysis have been done by several authors (13-21). Boland and Hardman (14) reported that the values of Km and kcat of actinidain with Z-Lys-ONp as substrate were 22 pM and 29 S-l, respectively, and were similar to those for papain. The major structural difference presumably related to the catalytic activity between actinidain and papain resides in the bottom of the S2 subsites, Met211 in actinidain and Ser205 in papain (22).
We have compared the steady-state and the pre-steady-state kinetic parameters with Z-Lys-ONp as substrate for two actinidain isozymes, AC-1 and AC-2; in order to reveal differences in the kinetic properties between the isozymes. We suggest that an acyl-enzyme formation is slightly more favored with AC-1 than with AC-2.
Department of Biotechnological Science, Kinki University, Wakayama 649-6493, Japan
Materials and Methods
The fruits of kiwifruit (Aclinidia deiiciosCl) were provided by the Yuasa Experimental Farm of Kinki University. The actinidain was extracted from kiwifruit pulp with 20 mM L-cysteine containing 1 mM EDTA, pH 5.0, and fractionated by salting out (20-80% saturated ammonium sulfate), and centrifuged at 10,000g for 30 min at 4°C. The precipitate was dissolved in 20 mM acetate buffer. pH 5.2, and dialyzed against the same buffer. The resulting solution was applied on aion-exchange chromatography using TSKgel Super-Q Toyopearl 650M (Tosoh, Japan) with a linear gradient of NaCI from 0.1 to 0.24 M. Each fraction (3 mlltube) of a main peak was analyzed by 12.5% acrylamide gel electrophoresis (native-PAGE) under acidic conditions (pH 4.0), and two purified actinidain isozymes were clearly identified. The isozymes were incubated to activate in 1 mM dithiothreitol for 60 min at 37°C, and excess dithiothreitol was removed by gel-filtration column PD-10 (Amersham Pharmacia Biotech, Japan). Then, each isozyme was immediately inactivated with 10 mM methyl methane thiosulfonate (SIGMA-ALDRICH JAPAN), and stored at 4°C until use. The inactivated isozyme was reactivated immediately before use by incubation with 1 mM dithiothreitol at 25°C for 60 min. Concentration of actinidain were calculated from measurements of UV absorbance with the molar absorption coefficient at 280 nm, 5.51
x 104M-1· cm-1 (2).
AC-1
I
I I
Fraction No. 22 29 31 33 35 55 75
AC-2
I 77 80 85 92
Fig.1 Two isozymes of actinidain separated on PAGE.
Partially purified actinidain was applied to anion-exchange chromatography with TSKgel Super-Q Toyopearl 650M under acidic conditions, 20 mM acetate buffer, pH 5.2, at 4°C. Each fraction was analyzed by 12.5% native-PAGE, pH 4.0.
105
Catalytic reaction of actinidain at 25°C in phosphate buffer (pH 6.5) was monitored at 325 nm with a stopped-flow spectrophotometer RA-2000 (Otsuka Electronics, Japan) with a 2.0 mm optical path. An equal volume of the substrate Z-Lys-ONp dissolved in acetonitrile/water (19: 1, v /v) and the reactivated actinidain in 50 mM phosphate buffer (pH 6.5) were mixed.
E
[E]o=
5.0 pM, [8]0=
500 pMC pH 6.5, 25°C II)
N
(f)
... ca
CI)
u
C
ca .c
a..o
tn.c
c:(
0.15
Abs=0.01
0.15
0.16 0.17 0.18 0.19
Time (sec)
0.45 0.20
Fig.2 Time-course of the release of p-nitrophenol from Z-Lys-ONp catalysed by actinidain.
These are the traces of the stopped-flow measurement. The main frame shows the transient phase of the reaction, and the inset shows the trace for longer period representing the steady-state.
31
The steady-state and the pre-steady-state kinetic parameters were evaluated according to the three-step mechanism (Scheme I).
k+l k+z k+3
E + S ~ E· S ~ ES' ~ E + P z (I)
kl +PI
In scheme I, E is the enzyme actinidain, S is the substrate Z-Lys-ONp, E . S is the reversible rapidly formed complex, ES' is the acyl • intermediate, PI and Pz are the hydrolysis products (p-nitrophenol and Cbz-Lys, respectively). k's are the reaction rate constants for each step.
The values of Km, kcat' and kca/Km were determined by Hanes-Woolf plots of [SJo/v o versus [SJ o with [SJ o» [EJ o, where suffix 0 means the initial values. Eqns. (0-(8) were used in the determination of kinetic parameters, Km acyl, B, and If (23). For the estimation of Vo and If,
the molar absorption coefficient E of PI, p-nitrophenol, at 325 nm, pH 6.5 was determined to be 9,310 M-I • cm-I.
-
U CD-
fI) ~...
...
3.0
2.0
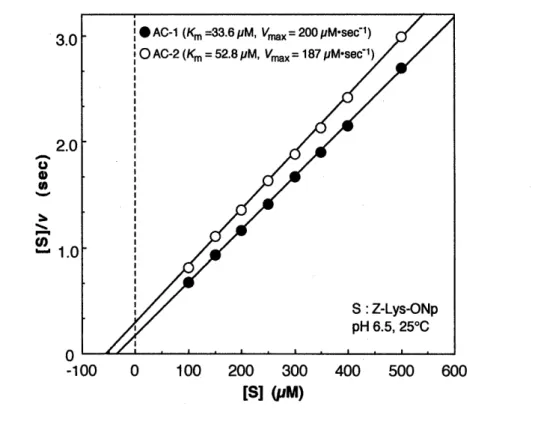
eAC-1 (Km=33.6pM, Vrnax =200pM·sec-1) o AC-2 (Km = 52.8 pM, Vrnax= 187 pM-sec-1)
!!!.
1.0S : Z-Lys-ONp pH 6.5, 25°C
o
~~~--~~----~~--~~~--~~~~-100
o
100 200 300 400 500 600[S] (PM)
Fig. 3 Determination of steady-state kinetic parameters for the hydrolysis of Z-Lys-ONp by actinidain isozymes.
Solid lines are the theoretical curves drawn with these parameter values.
(0
(2) (3) (4) (5) (6) (7) (8) (9)
J[ and B are the amplitude of the burst and the apparent first-order rate constant of the transient phase of p-nitrophenol release, respectively. The Vo in Eqn (5) is the steady-state
33 velocity defined by the Michaelis-Menten equation (Eqn (6)). When k+2 » k+3, the active center concentration of the enzyme can be determined from the intercept on the ordinate in the plots of 1/ ..rJ[ versus 1/[SJo according to Eqn (9). The values of B were determined from the slope of Guggenheim plot for the transient phase with In (J[ -a) versus time, where "a" represents A325 at each time. The plots of B against [SJ 0 were fitted to Eqn. (8) by non-linear least-squares regression. The values of k+3 and (k+2+k+3) were determined from the intercept on the ordinate and the maximum value of B at [sJ 0 -7 00 , respectively.
...
en ...
'"
Jc+
II CQ
x102
20 ..-....-...
~~.,...-...
15
5
o
e
AC-1o
AC-2pH 6.5, 25°C
0.2 0.4 0.6 0.8
[Z-Lys-ONp] (mM)
1.0
Fig. 4 Determination of rate constants for acylation and deacylation of enzyme in the hydrolysis of Z-Lys-ONp by actinidain isozymes.
Solid lines are the theoretical curves according to Eqn (8) drawn with the values of parameters shown in Table 2.
Table 1 The steady-state kinetic parameters of AC-l and AC-2 actinidain isozymes
AC-1 AC-2
Km
(uM)
33.6 52.8
k cat ( S -1 )
61.4 53.6
k cat I K m -1 -1 (UM ·s )
1.83.
1.02
Table 2 The presteady-state kinetic parameters of AC-1 and AC-2 actinidain isozymes
K acyl
m k +2 k +21K mBcy1
k +3 (ttM) ( s -1 ) (ttM -1 .S -1) ( S -1 )
AC-1 219 1890 8.63 51.4
AC-2 635 2860 4.50 61.9
Results and Discussion Separation of isozymes
Two isozymes of actinidain, AC-1 and AC-2, were clearly separated by using anion- exchange chromatography with a TSKgel Super-Q Toyopearl 650M column under acidic conditions (pH5.2). Fig. 1 showed that kiwifruit contained at least two isozymes of actinidain. The early fraction (fraction No. 29 - 35) and the late fraction (fraction No.
75 - 92) of peak showed different electrostatic properties in 12.5% native PAGE with acidic buffer (pH4.0), and these fractions were named AC-1 and AC-2, respectively. The pI values of AC-1 seemed higher than that of AC-2. The molecular mass of both isozymes we-r:e estimated 23,500 Da in 12% SDS-PAGE (data not shown). These results were in good agreement with a published report (2). AC-1 and AC-2 may correspond with KP4 and'KP6, respectively, which are the major components of six isozymes of actinidain r~ported by Sugiyama et al. (24).
Kinetic properties of the isozymes
The steady-state and the pre-steady-state kinetics for the hydrolysis of Z-Lys-ONp catalysed by actinidain isozymes, AC-1 and AC-2, may be consistently fitted to the three- step mechanism (Scheme 1). Under the steady-state conditions, actinidain catalysed the hydrolysis of Z-Lys-ONp according to Michaelis-Menten kinetics (Figs. 2 and 3). The kinetic parameters were obtained: Km=33.6 pM, Vmax=200 pM • sec-1 for AC-1 and Km=52.8 pM, Vmax=187 pM • sec-1 for AC-2 (Table O. The active enzyme concentration estimated from the burst amplitude n, according to Eqn (9) under the condition of [S]o --7 00
and k+2 » k+3, were 3.3 pM and 3.5 pM for AC-1 and AC-2, respectively. The total enzyme concentration employed, which were estimated from UV absorption, were 5.0 pM for each isozyme. Accordingly, the functional purity of the isozyme were 65.2% and 69.6% for AC-1 and AC-2, respectively. The kcat for AC-1 and AC-2 were estimated by kcat= V maxi [E] 0, in which
[E] 0 was the active enzyme concentration estimated from the burst as described above, to be 61.4 S-l and 53.6 s-\ respectively; however, kca/Km ratio was 1.8 times larger for AC-1 than for AC-2 (Table 1).
An initial burst can be observed in the progress curve (Fig. 2). The pre-steady-state kinetics can determine rate constants k+2 and k+3 according to Eqn (8) by non-linear fitting (Fig. 4). The values of k+2 and k+3 for AC-1 were 1,890 S-l and 51.4 S-l, and those for AC-2 were 2,860 S-l and 61.9 S-l, respectively (Table 2). Under the experimental conditions, the deacylation step (k+3) was clearly rate-limiting. The results show evidently that k+2 is much larger than k+3, and therefore that kcat should be closely similar to k+3 (Eqn (1)). The values
35 of k+z/ Km acyl ratio, which may be considered as the specificity constant for the formation of the acyl-enzyme intermediate (Scheme I), for AC-1 (8.63 pM-I·s- I) is two times larger than that for AC-2 (4.50 pM-I·s- I). The larger value of k+zlKmacYI suggests the higher stabilization of the tetrahedral intermediate in acylation step in AC-1.
Conclusion
The kinetic studies, steady-state and pre-steady-state, revealed that two isozymes of actinidain, AC-1 and AC-2, were essentially same in their kinetic behavior in the hydrolysis of Z-Lys-ONp as substrate. The de acylation step was clearly rate-limiting, and k+3 value was practically same to kcat. A small difference, however, in the values of kca/ Km and k+zI Km acyl
may suggest that the tetrahedral intermediate, which is considered as a transition state complex, is more stable with AC-1 than with AC-2. How the difference in the stability is related with the difference in electrostatic properties between two isozymes remains to be analyzed.
Acknowledgment
The authors wish to thank Prof. Akio Takebayashi of the Experimental Farm of Kinki University for supplying kiwifruit.
References
( 1) Arcus, A C. (1959) Proteolytic enzyme of Actinidia chinensis. Biochem. Biophys. Acta 33, 242-244.
(2) McDowall, M. A (1970) Anionic proteinase from Actinidia chinensis, preparation and properties of the crystalline enzyme. Eur. J. Biochem. 14, 214-221.
( 3) McDowall, M. A. (1973) The action of proteinase A2 of Actinidia chinensis on the f3 -chain of oxidized insulin. Biochem. Biophys. Acta 293, 226-231.
( 4) Baker, E.N. and Drenth, J. (1987) The cysteine proteinases: structure and mechanism.
In: Biological Macromolecules and Assemblies, vol. 3 (Jurnak, F. and McPherson, A, eds.) . pp. 313-368. John Wiley and Sons. New York.
( 5) Brocklehurst, K., Salih, E. and Lodwig, T.S. (1984) Differences between the electric fields of the catalytic sites of papain and actinidin by using the thiol-located nitrobenzofurazan label as a spectroscopic reporter group. Biochem. J. 220, 609-612.
(6) Brocklehurst, K., Willenbrock, F. and Salih, E. (1987) Cysteine proteinases. In:
Hydrolytic Enzymes (Neuberger, A. and Brocklehurst, K., eds.). pp. 39-158. Elsevier.
Amsterdam.
(7) Brocklehurst, K., O'Driscoll, M., Kowlessur, D., Phillips, I.R., Templeton, W., Thomas, E.W., Topham, C.M. and Wharton, C.W. (1989) The interplay of electrostatic and binding interactions determining active centre chemistry and catalytiC activity in actinidin and papain. Biochem. J. 257, 309-312.
( 8) Praekelt, U.M., McKee, R.A and Smith, H. (1988) Molecular analysis of actinidin, the cysteine proteinase of Actinidia chinensis. Plant Mol. Bioi. 10, 193-202.
( 9) Carne, A. and Moore, C.H. (1978) The amino acid sequence of the tryptic peptides from actinidin, a proteolytic enzyme from the fruit of Actinidia chinensis. Biochem. J.
173,73-83.
(10) Podivinsky, E., Forster, RL.S. and Gardner, RC. (1989) Nucleotide sequence of actinidin, a kiwi fruit protease. Nucleic Acids Res. 17,8363.
(11) Baker, E. N. (1980) Structure of actinidain, after refinement at 1.7
A
resolution. 1.Mol. Bioi. 141, 441-484.
(12) Sugiyama, S., Ohtsuka, K, Sata, K., and Kawabata, M. (1996) Purification and characterization of six kiwifruit proteases isolated with two ion-exchange resins, Toyopearl-Super Q and Bakerbond WP-PEI. Biosci. Biotech. Biochem. 60, 1994-2000.
(13) Boland, M. ]., and Hardman, M.]. (1972) Kinetic studies on the thiol protease from Actinidia chinensis. FEBS Lett. 27, 282-284.
(14) Boland, M. ]., and Hardman, M.]. (1973) The actinidain-catalyzed hydrolysis of N-alpha-benzyloxycarbonyl-L-Iysine p-nitrophenyl ester. Eur. 1. Biochem. 36, 575-582.
(15) Brocklehurst, K, Willenbrock, S.].F. and Salih, E. (1983) Effects of conformational selectivity and of overlapping kinetically influential ionizations on the characteristics of pH -dependent enzyme kinetics. Implications of free enzyme pKa variability in reactions of papain and for its catalytic mechanism. Biochem. 1. 211, 701-708.
(16) Brocklehurst, K, Brocklehurst, S.M., Kowlessur, D., O'Driscoll, M., Patel, G., Salih, E., Templeton, W., Quigley, K., Thomas, E.W., Wharton, C.W., Willenbrock, F. and Szawelski, R (1988) Supracrystallographic resolution of interactions contributing to enzyme catalysis by use of natural structural variants and reactivity-probe kinetics.
Biochem.l. 256, 543-555.
(17) Brocklehurst, K, Watts, AB., Patel, M., Verma, C. and Thomas, E.W. (1997) Cysteine proteinases. In: Comprehensive Biological Catalysis: A Mechanistic Reference (Sinnott, M.L., ed.) . pp. 381-424. Academic Press. London.
(18) Salih, E. and Brocklehurst, K. (1983) Investigation of the catalytic site of actinidin by using benzofuroxan as a reactivity probe with selectivity for the thiolate-imidazolium ion-pair systems of cysteine proteinases. Evidence that the reaction of the ion-pair of actinidin (pKr 3.0, pKIl 9.6) is modulated by the state of ionization of a group associated with a molecular pKa of 5.5. Biochem. 1. 213, 713-718.
(19) Salih, E., Malthouse, ].P.G., Kowlessur, D., Jarvis, M., 0' Driscoll, M. and Brocklehurst, K (1987) Differences in the chemical and catalytic characteristics of two crystallographic ally 'identical' enzyme catalytic sites. Characterization of actinidin and papain by a combination of pH-dependent substrate catalysis kinetics and reactivity probe kinetics and reactivity probe studies targeted on the catalytic-site thiol group and its immediate microenvironment. Biochem. 1. 247, 181-193.
(20) Kowlessur, D., O'Driscoll, M., Topham, C.M., Templeton, W., Thomas, E.W. and Brocklehurst, K (1989) The interplay of electrostatic fields and binding interactions determining catalytic-site reactivity in actinidin. A possible origin of differences in the behaviour of actinidin and papain. Biochem. 1. 259, 443-452.
(21) Topham, C.M., Salih, E., Frazao, C., Kowlessur, D., Overington, J.P., Thomas, M., Brocklehurst, S.M., Patel, M., Thomas, E.W. and Brocklehurst, K (1991) Structure- function relationships in the cysteine proteinases actinidin, papain and papaya proteinase O. Three dimensional structure of papaya proteinase 0 deduced by knowledge-based modelling and active centre characteristics determined by two- hydronic-state reactivity probe kinetics and kinetics of catalysis. Biochem. 1. 280, 79-92.
37
(22) Patel, M., Saleem, I., Mellor, C.W., Sreedharan, S., Templeton, W., Thomas, E.W., Thomas, M. and Brocklehurst, K. (1992) Variation in the P2-S2 stereochemical selectivity towards the enantiomeric N-acetylphenylalanylglycine-4-nitroanilides among the cysteine proteinases papain, ficin and actinidin. Biochem. 1. 281, 553-559.
(23) Fersht, A. (1999) Chapters 3 and 4. In: Structure and Mechanism in Protein Science.
pp. 103-168. W. H. Freeman & Co. New York.
(24) Sugiyama, S., Ohtsuki, K., Sato, K., and Kawabata, M. (1997) Enzymatic properties, substrate specificities and pH-activity profiles of two kiwifruit proteases. 1. Nutr. Sci.
Vitaminal 43, 581-589.
'[]Sl:f.:!>~
Y 7 =f-
-=-
~'~:/·
Y ~ V 'if ~ L. ~c J: lJ ~!k~ ~ tltc
N-a -Cbz-L-Lysine p --=-
r 07 J.. -=-JvIA T Jvt1Dzk7t~T,i1ZJltO)JE1#~:f\i~:tD J: L§iWJE1#~:f\i~O)~}jt~s~~T,i*JT
-t- ry
~ 7 Jv-'Y (Actinidia deliciosa) ~~iJ\ G" V A T ~ :/. 7°0 T Y --{i'C' d5 G Y 7 =f--=-~'~:/o)Y -iV'if-i L. 2*l~J{ (AC-1 t AC-2) ~~-i?t:/Xt~707 r 7'57 -{-~Cl¥l1$
m~ Ltc o ~ Y -i v'if-i L.O)JE1#~:f\i~tiWJE1#~:f\i~O)M~1ZJIt~" E1px~~'C'd5G N-alpha- carbo-benzoxy-L-lysine p-nitrophenyl ester ~ fflt.-'l" A r 'Y 7° r 7 0 -$~C J: lJ rWJJE L l
~~*JTLtco AC-1 t AC-2 O)filii~~C:tDt.-'l" YVJv1t1ZJIt~c1*-)J\-A r~mr~UL" M~1Z JltiJ"YiJ:
«
t t-=t~~g'C'Jiu c: t ~S)j G iJ'~c Ltco
filiiY -i V 'if -i L.O)M~1ZJltO)1$~t~~g~j:ij~YVJv1t1ZJIt'C'd5lJ" k cat ~j: k+3 ~clli:1~'C'~tco AC-1 0) kca/Km t k+/Kmacyl ~j: 1.83 IlM-1 • S-l
t 8.63IlM-1·s-1 'C'd5lJ" i-tl;C'tlAC-2 0)1~0) 21ti'C'd5-:Jtc o c:0)c: tiJ'G" AC-10)YV Jv1tM~$Fs'1*~j: AC-2 O)i-tlJ: lJ ~JE'C'd5 G c: t iJ"~~~ tltc o