つくばリポジトリ NC 9 499
全文
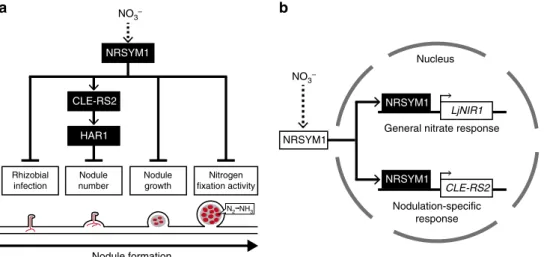
図
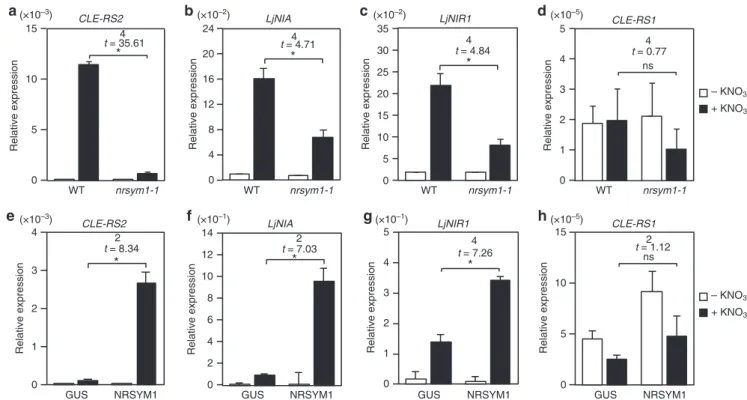
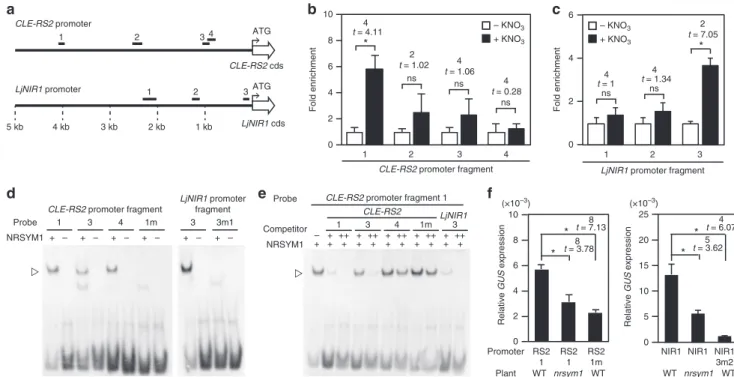
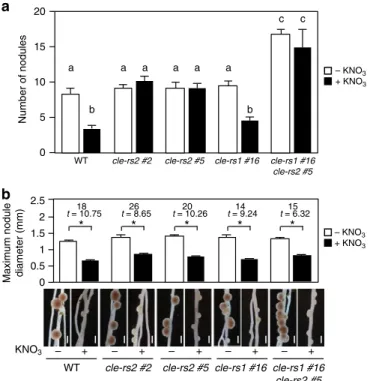
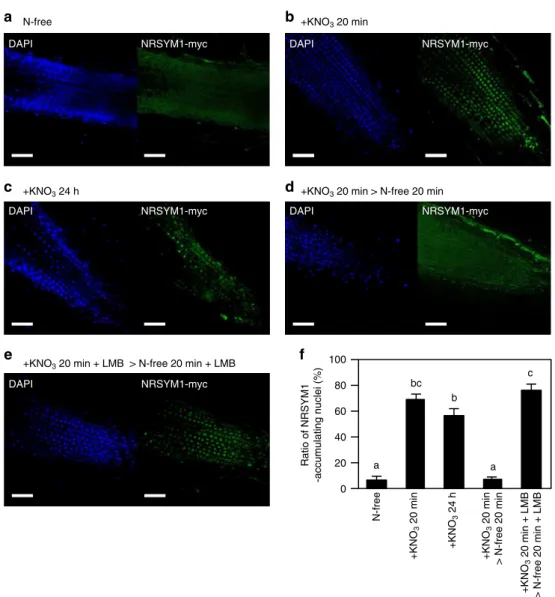
関連したドキュメント
initial functions are proved in the form of an integral maximum principle and conditions of transversality for nonlinear systems with a variable structure, delays and a
The only thing left to observe that (−) ∨ is a functor from the ordinary category of cartesian (respectively, cocartesian) fibrations to the ordinary category of cocartesian
The inclusion of the cell shedding mechanism leads to modification of the boundary conditions employed in the model of Ward and King (199910) and it will be
In this paper we develop a general decomposition theory (Section 5) for submonoids and subgroups of rings under ◦, in terms of semidirect, reverse semidirect and general
On the other hand, when M is complete and π with totally geodesic fibres, we can also obtain from the fact that (M,N,π) is a fibre bundle with the Lie group of isometries of the fibre
This paper develops a recursion formula for the conditional moments of the area under the absolute value of Brownian bridge given the local time at 0.. The method of power series
Answering a question of de la Harpe and Bridson in the Kourovka Notebook, we build the explicit embeddings of the additive group of rational numbers Q in a finitely generated group
Then it follows immediately from a suitable version of “Hensel’s Lemma” [cf., e.g., the argument of [4], Lemma 2.1] that S may be obtained, as the notation suggests, as the m A
