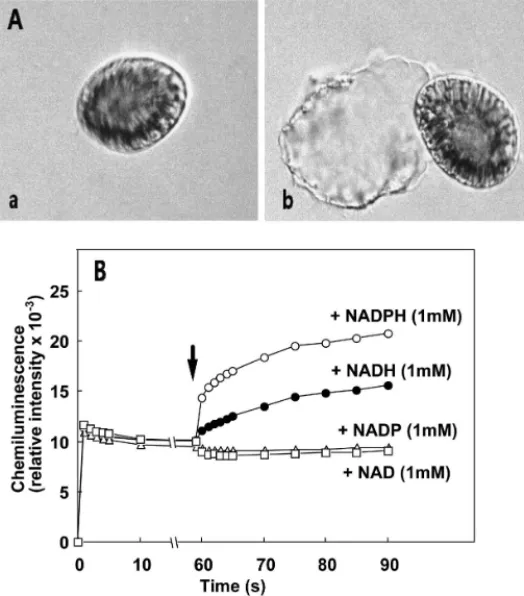
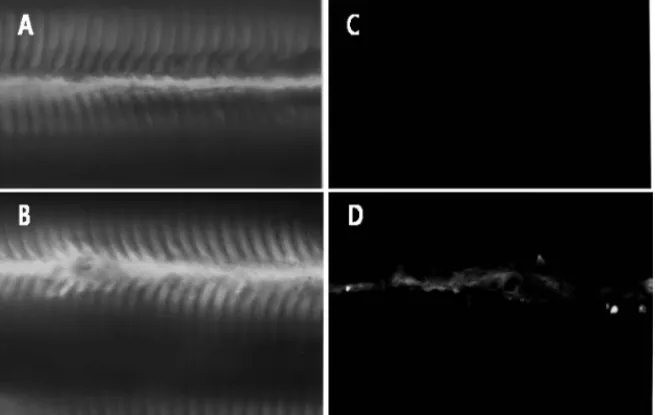
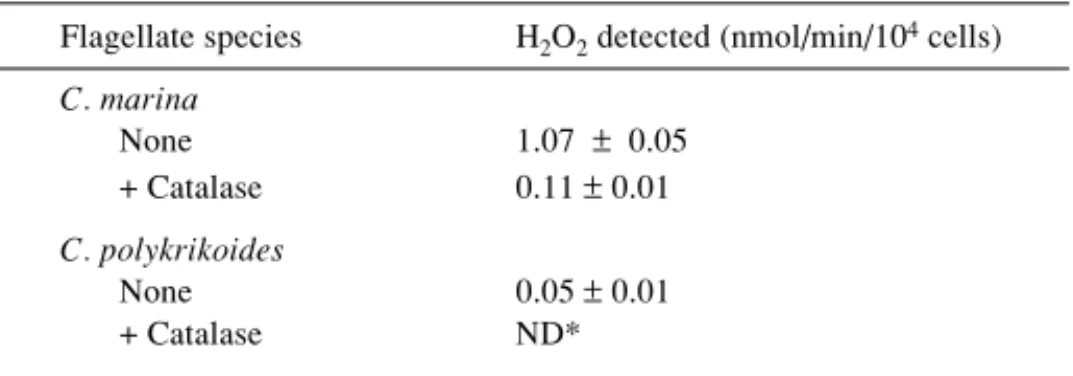
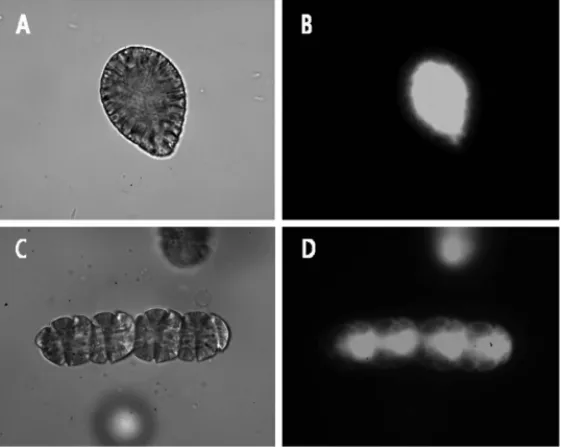
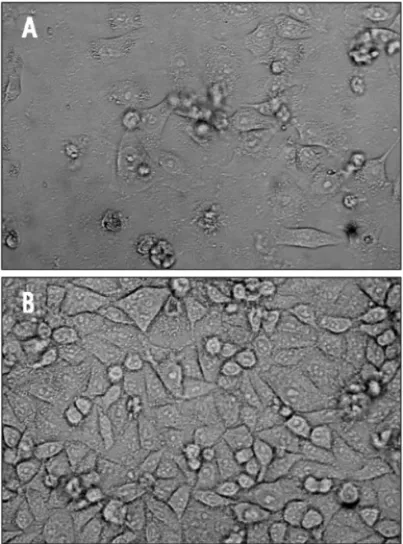
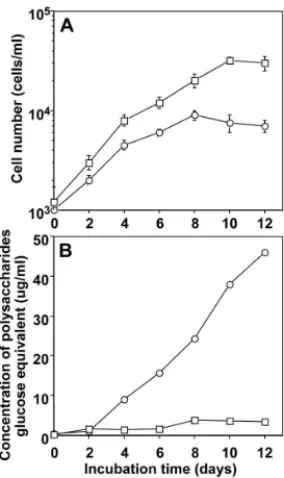
245
© by TERRAPUB and Nagasaki University, 2010.
Possible Factors Responsible for the Fish-Killing Mechanisms of the Red Tide Phytoplankton, Chattonella marina and
Cochlodinium polykrikoides
Daekyung K IM
1and Tatsuya O DA
21
Jeju Center, Korea Basic Science Institute (KBSI), 66 Jejudaehakno, Jeju-Si,
Jeju Special Self-Governing Province 690-756, Korea
2