脱澱粉小麦ふすま培養における麹菌酵素生産
金子明裕1,2・尾関健二1(1金沢工業大学ゲノム生物工学研究所,
2日清ファルマ株式会社)
平成 28 年 1 月 15 日受理
Selective production of cellulase and hemicellulase from Aspergillus oryzae using starch-free wheat
bran.
Akihiro KANEKO1, 2, Kenji OZEKI1
(1Genome Biotechnology Laboratory, Kanazawa Institute
of Technology(Yatsukaho 3-1, Hakusan, Ishikawa
924-0838, Japan);2Nisshin Pharma Inc.(Kanda-Nishiki-cho
1-25, Chiyoda-ku, Tokyo 101-8841, Japan))
In order to produce enzymes for degrading hemicellulose--which is the main component of wheat bran(WB), starch-free wheat bran (SFWB)was prepared by acid hydrolysis in an
autoclave and cleaning with hot water. After culturing Aspergillus oryzae for 7 days with SFWB as a substrate, the expression of α-amy-lase was suppressed, while the production of hemicellulase and cellulase was confirmed; these were not observed in the WB culture. In addi-tion, the enzyme activity of β-xylanase and β-xy-losidase was increased. Enzyme production from wheat bran hemicellulose-derived hemicellulase is now feasible using Aspergillus cultured in SFWB.
Key words: Aspergillus oryzae,hemicellulase, starch, wheat bran,Aspergillus oryzae,ヘミセ ルラーゼ,澱粉,小麦ふすま ─────
緒 言
小麦ふすまは小麦製粉時の副生成物として,日本国 内だけでも年間 50 万トンに達する1)。集積の手間が かからず,かつ性状が均一であることから,他の食品 J. Brew. Soc. Japan.
Vol.112, No. 1, p. 77 ~ 81(2017) る。機能性食品素材2, 3)としての価値が見出されては いるが,その多くは飼料もしくは食品廃棄物となって いるのが現状である4, 5)。この小麦ふすまを利用した 物質生産や成分の有効活用は食物系バイオマス資源の 有効な活用となる。小麦ふすまを利用した麹菌 As-pergillus oryzae 酵素についての研究・生産はタカジ アスターゼといった消化酵素薬の開発から始まり,国 内外で長年行われている。麹菌は食品・発酵産業で永 年使用されてきた安全な微生物であり6),さまざまな 酵素を多量に生産・分泌する7, 8)。2005 年に全ゲノム 配列の解読が終了し,全遺伝子の解析が可能となっ た9)。小麦ふすまを有効利用するには,主成分である ヘミセルロース・セルロースを分解することが求めら れる。農産副産物のバイオマスを基質とした酵素生産 と し て Penicillium echinulatum や Aspergillus niger によるヘミセルロース分解の研究がなされている が10, 11),今回,食品用途に好適で全ゲノム配列の解 析が終了した麹菌を用いて小麦ふすま培養における酵 素生産を試みた。小麦ふすまは,約 10% の澱粉を含 んでいる12, 13)。そのため,小麦ふすまを培地とした麹 菌培養(発酵)では,この澱粉の存在によりα- アミ ラーゼ及びグルコアミラーゼといったアミラーゼ系酵 素が誘導され,ヘミセルロースを分解する加水分解酵 素の発現・生産が抑えられることとなっていた。我々 は,脱澱粉小麦ふすまを作製,基質として麹菌を培養 することで,アミラーゼ系酵素の発現を抑え,他の酵 素タンパク質生産を検討した。特に小麦ふすまの主成 分であるヘミセルロースを分解するヘミセルラーゼを はじめ,アミラーゼ系酵素以外の加水分解酵素生産を 誘導し,今まで産生されていなかった酵素を見つける こと,さらにキシラナーゼ・キシロシダーゼ主体の酵 素生産を検討したので報告する。 実験方法 1.菌株 麹菌 A. oryzae RIB40 株を使用した。 2.脱澱粉小麦ふすまの作製と処理後の澱粉量の測定 小麦ふすま(Wheat Bran:WB)に 10 倍量の 50 mM クエン酸溶液を加え 120 ℃・15 分間のオートク レーブ処理後,50 倍量の熱水に 1 分間浸漬,その後 150倍量の熱水で洗浄,さらに 20 倍量の蒸留水で 5
醸 協( ) 回 洗 浄 を 行 い, 脱 澱 粉 小 麦 ふ す ま(Starch Free Wheat Bran:SFWB)を作製した。処理により,一 部の可溶性ヘミセルロース,ビタミン,ミネラル等は 溶出していると考えられるが,脱澱粉を目的に行った。 WB,SFWB の澱粉量は F- キット スターチ(JK インターナショナル)を用いて測定を行った。試料を 溶液中でアミログルコシダーゼによりグルコースに分 解,グルコースをヘキソキナーゼ反応により ATP で リン酸化,次に NAD 存在下グルコース 6- リン酸デ ヒドロゲナーゼ触媒下で生成する NADH を 340 nm の波長を測定し,澱粉量の定量を行った。 3.培養方法 WB と 同 量 の 水 分 量 に な る ま で 乾 燥 処 理 し た SFWB 及び対照として WB をそれぞれフラスコに 2 g ずつを取り,滅菌水 1.2 mL を加え,121 ℃・15 分間 のオートクレーブ後,麹菌の胞子 108を植菌し,30 ℃にて 7 日間固体培養を行った。培養後,培養固体か ら抽出 Buffer(10 mM 酢酸 Buffer(pH 5.0)10 mL, 0.5%NaCl)を用い,3 時間常温で抽出した後,ろ紙 (No2.アドバンテック東洋)でろ過,さらに 0.45 μm の Minisart フィルター(Sartorius stedim japan)を 用いてろ過を行い,サンプル酵素液とした。
4.タンパク質の定量
タンパク質の定量は Bovine Serum Albumin 及び Quick Start プロテインアッセイ染色液(Bio-Rad)を 用い,Bradford 法により行った。 5.プロテオーム解析 タンパク質を Centrifugal Device(日本ポール)を 用いて 300 μg になるように調製し,2 次元電気泳動 を AE-6540B(アトー)により行った。この電気泳動 ゲルからスポットを切り出し,タンパク質の同定をペ プチドマスフィンガープリント(PMF)法11)により 行った。2 次元電気泳動から分離されたタンパク質を トリプシンなどの基質特異性の高いプロテアーゼで消 化し,得られたペプチド断片をマトリックス支援レー ザーイオン化飛行時間質量分析計 MALDI-TOF MS AXIMA-CFR PLUS(島津製作所)を用いて,精密な 質量分析を行い,タンパク質のフィンガープリントを 得,DOGAN(www.bio.nite.go.jp/dogan/project/ view/AO)のデータベースを使用し,酵素のアミノ 酸配列からタンパク質を同定した15)。 6.酵素活性の測定 β- キシラナーゼ活性は,Birchwood xylan(シグマ アルドリッチ)を用い,基質濃度 2(w/v)%,pH 5.0,40 ℃にて 10 分間の反応により生成したキシロー スをジニトロサリチル酸(DNS)法16)により測定し, 1分間に 1 μmol の還元糖を生成する活性を 1 酵素単 位(U)とした。β- キシロシダーゼ活性は p- ニトロ フェニル -β-D- キシロピラノシド(シグマ アルドリッ チ)を用い,基質濃度 1 mM,pH 5.0,40 ℃で 10 分 間の反応により生成した p- ニトロフェノール(p-NP)を 420 nm の吸光度から測定し定量した。1 分間 に 1 μmol の p-NP を遊離させる活性を 1 U とした。 エンドペプチダーゼ活性は 2% カゼインを基質として 用い,pH 3.0 と pH 6.0 の活性を測定した17)。酵素活 性 1 U はカゼインに 37 ℃で作用して 1 分間に 1 μmol のチロシン相当量の可溶性ペプチドを遊離させる酵素 量とした。酸性カルボキシペプチダーゼ活性の測定は 酸性カルボキシペプチダーゼ測定キット(キッコーマ ン)を用いた。カルボベンゾキシ -L- チロシル -L- ア ラニンから 37 ℃で 1 分間に 1 μmol の L- アラニンを 遊離する活性を 1 U とした。α- アミラーゼ活性は,α-アミラーゼ測定キット(キッコーマン)を用いた。37 ℃で 1 分間に 2- クロロ -p-NP-65- アジド -65 デオキシ -β- マルトペンタオシドから 1 μmol の 2- クロロ -p-NP を遊離させる活性を 1 U とした。糖化力の測定は糖 化力測定キット(キッコーマン)を用いた。p-NP-β-マルトシドから 37 ℃で 1 分間に 1 μmol の p-NP を遊 離する活性を 1 U とした。酵素活性は培養抽出液の 酵素タンパク質量(μg)当たりの活性値(Unit)で 表される比活性(Unit/μg)で表した。 実験結果および考察 1.脱澱粉処理後の澱粉量とアミノ酸 SFWB の澱粉量は検出限界(6 mg/g)以下であっ た。WB に含まれている澱粉の 90% 以上を除去する こ と が で き た。 ま た, ア ミ ノ 酸 量 は WB の 721.7 mg/100 g に 対 し て,SFWB は 542.1 mg/100 g と 減 少したが,アミノ酸比に大きな変化は無かった(data not shown) 2.タンパク質量 培養後,抽出酵素液中のタンパク質量は SFWB が 118 μg/mL,WB が 969 μg/mL であった。その抽出
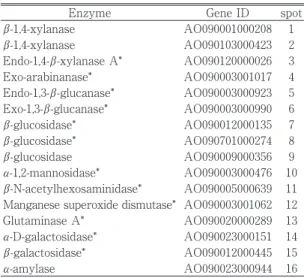
液をタンパク質濃度が 300 μg となるように調製し,2 次元電気泳動を行った。 3.プロテオーム解析 2 次元電気泳動によるプロテオーム解析を行った結 果,澱粉質を除去していない WB 培養から調製した 酵素液ではα- アミラーゼの強いスポット(Fig. 1(A) のスポット No.16)を検出したが,SFWB で培養・調 製した酵素液では,α- アミラーゼの産生がみられな くなり,新たな酵素の産生が見られた(Fig. 1(B) のスポット No.3-8,No.10-15)。この電気泳動により 現れた酵素を同定したところ,WB 培養では生産が認 められなかった Endo-1,4-β- キシラナーゼ A,アラビ ナナーゼ,Endo-1,3-β- グルカナーゼ,Exo-1,3-β- グル カナーゼ,β- グルコシダーゼといったヘミセルラー ゼ,セルラーゼ系の産生が確認された(Table 1)。ア ミラーゼ系酵素の生産が抑えられることで,ヘミセル ラーゼ・セルラーゼ系酵素の生産が上昇し,WB を用 いた培養方法では産生されなかった酵素を産生するこ とができた。 4.酵素活性 麹菌を 7 日間培養後のβ- キシラナーゼ活性は,WB 培養では 135 U であったの対し,SFWB 培養では 582 U と活性が上昇した。小麦ふすまの主成分であるキシ ランを分解する酵素を主に生産することができた。β-キシロシダーゼ活性も高くなった。β- キシロシダー ゼは基質が増大することで,その生産量が多くなった と考えられた。また,Endo 型ペプチダーゼ(酸性プ ロテアーゼ,中性プロテアーゼ),さらに Exo 型ペプ チダーゼである酸性カルボキシぺプチダーゼの酵素活 性が上昇した。小麦ふすまは 10% 以上のタンパク質 を含む14)が,これらの分解に関する酵素生産を上げ ることもできた。澱粉の代わりとしてタンパク質を栄 養源として利用していると考えられた。なお,α- ア
Fig. 1 Comparative proteome analysis of secreted proteins using control wheat bran WB:(A), and SFWB(B). Each gel was loaded with 300 μg of secreted protein and stained with CBB.
Enzyme Gene ID spot
β-1,4-xylanase AO090001000208 1 β-1,4-xylanase AO090103000423 2 Endo-1,4-β-xylanase A* AO090120000026 3 Exo-arabinanase* AO090003001017 4 Endo-1,3-β-glucanase* AO090003000923 5 Exo-1,3-β-glucanase* AO090003000990 6 β-glucosidase* AO090012000135 7 β-glucosidase* AO090701000274 8 β-glucosidase AO090009000356 9 α-1,2-mannosidase* AO090003000476 10 β-N-acetylhexosaminidase* AO090005000639 11 Manganese superoxide dismutase* AO090003001062 12 Glutaminase A* AO090020000289 13
α-D-galactosidase* AO090023000151 14
β-galactosidase* AO090012000445 15
α-amylase AO090023000944 16
*:The spot that was detected newly in SFWB Table 1 Identification of proteins(Fig. 1)
醸 協( ) ミラーゼ活性は WB 培養の 16.2 U に比べ SFWB 培養 では 0.2 U と抑えられた。糖化力も 1/10 以下に低下 した(Fig. 2)。 微生物のタンパク質生産能には限界があり,遺伝子 の変異や破壊を行わずに,酵素の生産比率を制御する ことは難しい。本工程で作製した SFWB を基質に培 養することで,遺伝子の破壊や変異を行うことなく, アミラーゼ系酵素の生産を抑えることができ,今まで 産生の確認できなかった酵素の生産を可能とした。小 麦ふすま中に豊富に存在するキシランを分解するβ-キシラナーゼ,β- キシロシダーゼ,さらにキシラン の側鎖につくアラビノース18)を分解するアラビナナ ーゼといった酵素を効率よく生産することに成功した。 ヘミセルロースは植物種によって構造が異なり,市販 の酵素剤を使用しても分解効率は低い。植物種に応じ たヘミセルラーゼ組成の酵素剤を作製することで効率 の良い酵素分解が可能になる。小麦ふすまで生産され たヘミセルラーゼ製剤は小麦ヘミセルロースの分解に 適したものと想定される。小麦ふすまヘミセルロース の構成糖である D- キシロース,L- アラビノース,ア ラビノキシランなどには血糖値上昇抑制作用や免疫賦 活作用3)等の機能が知られている。本研究により SFWB から開発されたヘミセルラーゼ製剤は,それ ら機能性糖の小麦ふすまからの可溶化にも有効である と考えられる。さらに,グルコース飢餓状態ではアミ ノ酸を栄養源として活発に利用する必要が生じ,エン ドペプチダーゼ活性が上昇したと考えられた。ヘミセ ルラーゼの生産量の増加と並んで,Endo 型,Exo 型 両ペプチダーゼの生産上昇も特徴的な酵素生産となっ た。今後は,様々な異種タンパクの高生産培地として の利用も検討する予定である。脱澱粉処理により水溶 性ビタミン・ミネラル類が溶出してしまうことが生育 に影響していると考えられるので,これら成分を添加 しての培養試験を行い,酵素生産性の向上,新規酵素 剤開発も検討していきたい。 要 約 小麦ふすま(WB)をクエン酸水溶液中でオートク Fig. 2 Enzyme production by A. oryzae in SFWB and WB medium. (A)β-xylanase, (B)β-xylosidase, (C)α
-amylase, (D)Diastatic power, (E)Acid-carboxypeptidase, (F)Endo-peptidase pH3.0, (G)Endo-peptidase pH6.0
レーブ処理する酸加水分解と熱水洗浄により脱澱粉小 麦ふすま(SFWB)を調製した。この SFWB を基質 に麹菌の培養を行い,産生する酵素について 2 次元プ ロテオーム解析と各種酵素活性を比較した。SFWB 培地における培養では,WB 培養では認められなかっ た Endo-1,4-β- キシラナーゼ A,アラビナナーゼ, Endo-1,3-β- グルカナーゼ,Exo-1,3-β- グルカナーゼ, β- グルコシダーゼといったヘミセルラーゼ,セルラ ーゼ系酵素の産生が確認された。アミラーゼ系酵素の 生産が抑えられ,β- キシラナーゼ,β- キシロシダー ゼの酵素活性が増加した。澱粉質のない小麦ふすま培 養で,小麦ふすまヘミセルロースから誘導されるヘミ セルラーゼ主体の酵素の生産を可能とした。 文 献 1) 89 次 農 林 水 産 省 統 計 表: 農 林 水 産 省,613 (2014)
2) S.R. Bandaru, H. Yoshinobu, A.C. Leonard, S. Barbara, C. Indrane, and V.R. Chinthalapally: Cancer Res., 60, 4792-4297(2000)
3) 物部真奈美,山本(前田)万里,松岡由記,金 子 明 裕, 平 本 茂: 食 科 工,55,245-249 (2008)
4) J. Angulo, L. Mahecha, S. Yepes, A. Yepes, G. Bustamante, H. Jaramillo, E. Valencia, T. Villa-mil, and J. Gallo: J Environ Manage., 95, 203-209(2012)
5 ) 渡辺久典:日本エネルギー学会誌,87,846-851(2008)
6) P. Barbesgaard, H. P. Heldt-Hansen, and B. Diderichsen: Appl. Microbiol. Biotechnol., 36, 569-572(1992)
7) 布川弥太郎:澱粉科学,33,95-103(1986) 8) N. Kitamoto, S. Yoshino, K. Ohmiya, and N.
Tsukagoshi: Biosci. Biotechnol. Biochem., 63, 1791-1794(1999)
9) M. Machida, K. Asai, M. Sano, T. Tanaka, T.
Kumagai, G. Terai, K. Kusumoto, T. Arima, O. Akita, Y. Kashiwagi, K. Abe, K. Gomi, H. Hori-uchi, K. Kitamoto, T. Kobayashi, M. TakeHori-uchi, D.W. Denning, J.E. Galagan, W.C. Nierman, J. Yu, D.B. Arche, J.W. Bennett, D. Bhatnagar, T.E. Cleveland, N.D. Fedorova, O. Goto, H. Horikawa, A. Hosoyama, M. Ichinomiya, R. Ig-arashi, K. Iwashita, P.R. Juvvadi, M. Kato, Y. Kato, T. Kin, A. Kokubun, H. Maeda, N. Maeyama, J. Maruyama, H. Nagasaki, T. Naka-jima, K. Oda, K. Okada, I. Paulsen, K. Sakamo-to, T. Sawano, M. Takahashi, K. Takase, Y. Terabayashi, J.R. Wortman, O. Yamada, Y. Ya-magata, H. Anazawa, Y. Hata, Y. Koide, T. Ko-mori, Y. Koyama, T. Minetoki, S. Suharnan, A. Tanaka, K. Isono, S. Kuhara, N. Ogasawara, and H. Kikuchi:Nature, 438, 1157-1161(2005) 10) M. Camassola, and A. J. Dillon:J Appl
Micro-biol., 103, 2196-2204(2007)
11) C. Ottenheim, C. Verdejo, W. Zimmermann, and J. C. Wu: J Biosci Bioeng., 118, 696-701 (2014)
12) R.A. Burton, and G.B. Fincher:Front Plant Sci., 130, 1-6(2012)
13) A. Bacic, and B. Stone:Plant Biol., 8, 475-495 (1981)
14) 吉野健一,大城紀子,徳永千春,米澤一仁:J. Mass Spectrom. Sos. Jpn., 52, 1157-1161(2004) 15) K. Oda, D. Kakizono, O. Yamada, H. Iefuji, O. Akita, and K. Iwashita:Appl Environ Micro-biol., 72, 3448-3457(2006) 16) 福井作造:生化学実験法 1 還元糖の定量法 第 2版,学会出版センター,23(1990) 17) 横山定治,木場洋次郎,亀岡恵子,峰 弘子: 醸協,106,253-258(2011) 18) 長尾精一:ふすまと胚芽,「小麦の科学」,第 1 版,長尾精一編,(朝倉書店,東京),193-197 (1995)