The molecular mechanism of hot defensive bee
ball in Apis cerana japonica
著者
上岡 駿宏
号
18
学位授与機関
Tohoku University
学位授与番号
生博第409号
氏 名 ( 本 籍 地 )
学 位 の 種 類
学 位 記 番 号
学 位 授 与 年 月 日
学 位 授 与 の 要 件
研 究 科 , 専 攻
論
文
題
目
博士論文審査委員
カミオカ タカヒロ上岡 駿宏
博士(生命科学)
生博第 409 号
令和2年9月25日
学位規則第4条第1項該当
東北大学大学院生命科学研究科
(博士課程)生態システム生命科学専攻
The molecular mechanism of hot defensive bee ball in Apis
cerana japonica(ニホンミツバチの熱殺蜂球形成に関わる
分子基盤の解明)
(主査)教 授 河田 雅圭
教 授 佐藤 修正
教 授 竹内 秀明
論文内容の要旨
BackgroundHoneybees inhabit a wide temperature range from tropical to temperate, which is exceptional in insects. This is because honeybees have the ability to regulate body and nest temperatures by metabolic regulation and temperature sensitivity, and high temperature tolerance. Therefore, studying thermoregulation and thermotolerance of honeybees is essential for understanding insect temperature adaptation.
Apis cerana japonica, which is a subspecies of the A. cerana and distributes from Honsyu to Kyusyu in
Japan, exhibits specific defensive behavior by utilizing thermoregulation and thermotolerance. This behavior is called a hot defensive bee ball, and is performed when the bee nest is attacked by hornets. A bee ball is formed by hundreds of bee workers catching a hornet. The bee ball state is maintained for about 30 minutes, and its core temperature reaches ~46 °C, which is higher than the lethal temperature of hornets. This temperature is produced by metabolic heat by the flight muscles of bees. Thus, the bee ball is a behavior in which bee ball specific thermoregulation near the lethal temperature is performed by utilizing the heat production ability of flight muscles. To clarify what kind of molecular mechanism is involved in bee ball specific thermoregulation not only leads to understanding of thermoregulation and thermotolerance of honeybees, but also to a new perspective on insect temperature adaptation. Therefore, I conducted two studies to elucidate the molecular mechanism of the hot defensive bee ball.
1. Genes associated with hot defensive bee ball in the Japanese honeybee, Apis cerana japonica
In this chapter, in order to detect candidate genes related to the hot defensive bee ball in Apis cerana japonica, we measured the gene expression level using RNA-seq. Honeybee workers 15 days after emergence were prepared and sampled under the following three conditions; bee ball formation 30 minutes (“balling”), 30 minutes incubation of standard temperature 31°C (“control”), 30 minutes high temperature 46°C (“heat”). RNA-seq was used to measure gene expression levels in the brain, fat body, and flight muscle. The gene expression levels of “balling” and “control” (“comparison 1”) or “heat” and “control” (“comparison 2”) were compared, and differentially expressed genes (DEG) were calculated. The DEGs included in comparison 1 from which the DEGs overlapping with comparison 1 and comparison 2 was removed was defined as the DEG only in the bee ball (“ball-only”).
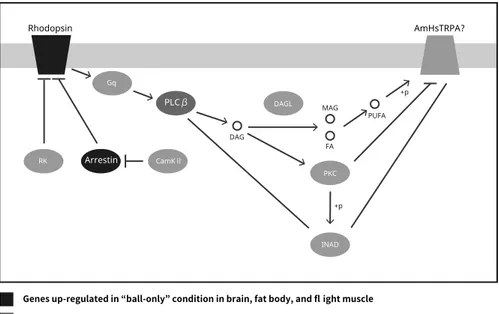
We observed that genes related to rhodopsin signaling (i.e., rhodopsin long wavelength and arrestin 2) were increased in all these tissues during balling in the group of “ball-only” (Figure 1). Since rhodopsin signaling was known to affect the thermotaxis via the thermosensitive channel TrpA1 in Drosophila, the results suggested that these DEGs contribute to the maintaining the bee ball by affecting the heat sensitivity. In the brain, genes related to behavior, inflammation, and glucose metabolism involved; in the fat body, genes related to energy metabolism, stress tolerance, and immunity involved; in the flight muscle, genes related to the exoskeleton involved in DEGs of “ball-only”.
2. Thermal sensitivity in the Japanese honeybee, Apis cerana japonica
The results in Chapter 1 suggested that the rhodopsin signal(signal or signaling) may influence the heat sensitivity through the thermosensitive channel during the hot defensive bee ball. However, it remains unclear whether the
thermosensitive channel of Apis cerana japonica is involved in heat sensitivity. Here, in Chapter 2, in order to clarify the detailed molecular mechanism involved in thermosensing in Apis cerana japonica, I performed heat avoidance experiments on workers which inhibited the temperature sensitive channel AmHsTRPA. The bee workers that had been incubated overnight were anesthetized, and 1 μl of 10 mM of menthol, which is an AmHsTRPA inhibitor, or water was injected into the head. The bee workers awakened from anesthesia were placed in a case, the case was placed in a heat block heated at high temperature (55℃) or standard temperature (31℃), and the time the honey bee stayed on the heated surface of the case was calculated.
In workers injected with water into the brain, the heat avoidance of workers placed in a heat block heated to high temperature was lower than that in low temperature. I also demonstrated that workers injected with menthol reduced high heat avoidance than those injected with water.
Conclusion
In Chapter 1, I demonstrated that the expression level of genes related to the rhodopsin signaling was increased in “ball-only” group. In Chapter 2, I showed that inhibition of thermosensitive channel reduced the heat avoidance. These results suggest that rhodopsin signaling may inhibit AmHsTRPA and reduce its heat avoidance during bee ball. In chapter 1, I also demonstrated that genes whose expression was changed in each tissue in the group of “ball-only”. These findings will give a new perspective to bee ball specific thermoregulation in A. c. japonica.
Figure 1. A model for the molecular mechanism of thermosensing by rhodopsin signaling during the balling
behavior. The boxes indicate the genes involved in rhodopsin signaling (see “phototransduction - fly” in KEGG pathway) and expressed in all tissues in A. cerana japonica
論文審査結果の要旨
上岡駿宏氏提出の論文では、ニホンミツバチApis cerana japonica はオオスズメバチなどの捕食者に 対し熱殺蜂球形成に関わる分子機構の解明を試みた。熱殺蜂球では、数百匹のワーカーが捕食者を 取り囲み、飛翔筋を緊張させて約46°C の高温を発生させる。この高温状態は数十分間維持される が、どのような分子メカニズムによりワーカーが高温を感知し、発熱を調節しているのかについて は不明であった。上岡駿宏氏は、熱殺蜂球形成の高温維持に関わる候補遺伝子を探索することを目 的とし、RNA-seq 法を用いて熱殺蜂球形成時・実験的温度上昇時(46℃)・常温時(31℃)の3条 件におけるワーカーの遺伝子発現を測定した。熱殺蜂球形成時と常温時を比較して発現変化した遺 伝子から、実験的温度上昇時と常温時を比較したときに発現変化した遺伝子を取り除くことにより、 熱殺蜂球形成時に積極的に発現変化する遺伝子を絞り込んだ。脳・飛翔筋・脂肪体の3組織で熱殺 蜂球形成時のみ発現が上昇している遺伝子にロドプシンとアレスチンが検出された。ロドプシンは G タンパク質共役受容体として、温度感受性チャネルである AmHsTRPA シグナルの上流に位置す ることが示唆されている。AmHsTRPA の阻害剤をもちいて、行動実験を行った結果、AmHsTRPA が高温忌避に関与していることが示された。これらの結果から、ニホンミツバチは熱殺蜂球時にロ ドプシンからのシグナルを通じて、通常体温時とは異なる温度感受や調節を行っていることが示唆 された。これらの結果は、ニホンミツバチにおける他の昆虫ではみられない高温での行動の機構解 明に大きな示唆を与えるものである。これらの研究は、上岡駿宏氏が主導で行った研究で、上岡駿 宏氏は研究活動を行うに必要な高度の研究能力と学識を有することを示している。したがって,上 岡駿宏氏提出の論文は,博士(生命科学)の博士論文として合格と認める。