DOI: http://dx.doi.org/10.14947/psychono.34.6
短時間順応下での時間知覚
―方位選択的処理の寄与―
佐 藤 駿
a,*・中 沢 仁
b a専修大学大学院文学研究科心理学専攻 b専修大学人間科学部心理学科Orientation selectivity contributes to time perception
under short adaptation condition
Shun Sato
a,* and Hitoshi Nakazawa
ba Graduate School of the Humanities, Major in Psychology, Senshu University, b Department of Psychology, School of Human Sciences, Senshu University
Human time perception is known to be distorted by adaptation to visual motion stimuli. Many studies have in-vestigated time perception distortion and its underlying mechanisms, but their results have been inconsistent. This may be because they have used different adaptation durations, as it is known that time duration can affect visual processing levels. Previous studies have elucidated how time perception is distorted under adaptation conditions of a few seconds or much longer, but have not investigated this distortion under shorter durations. Therefore, we investi-gated how time is distorted and how time perception mechanisms are affected under short adaptation durations (e.g., <2 s). We found that a short duration produced a characteristic time distortion. In addition to orientation
se-lectivity, we confirmed a transient effect on time perception distortion. These results suggest that visual processing concerning orientation selectivity contributes to time perception, and that transient components of the adaptation effect are useful for investigating the mechanisms of adaptation-based perceived time distortion.
Keywords: time perception, visual motion, visual adaptation, short time adaptation, orientation selectivity
1. は じ め に ヒトは何らかの方法で時間を知覚し,外界からの感覚 情報がないときであっても,時間だけに即して適応的な 行動を取ることができる場合がある。時間それ自体への 具体的な感覚器を持たない我々の内的機構において,時 間知覚はどのように実現されているのだろうか。短い時 間,特に数ミリ秒から数秒といった時間知覚の機序に関 しては未だ明らかになっていない(Mark & Buonomano, 2004)。内的な時計(internal timer)を仮定する感覚的処 理モデル(sensory processing model)(松田他,1996)と いったモデルが考えられているものの,このモデルを実
現する神経機構が実証されるには至っていない。近年の 時間知覚研究から,視覚的運動情報や変化頻度の情報 が,内的時計のパルス頻度を変化させている可能性も示 唆されている(e.g., Kaneko & Murakami, 2009)。感覚的 処理モデルが正しいとするならば,感覚刺激からどのよ うに時間情報が抽出され,どのような処理過程によって 内的時計のパルスが生成されているのかを,視覚運動処 理と時間知覚との関連とともに解き明かさなければなら ない。 たとえば視覚刺激の知覚時間への影響を,順応によっ てとらえようとする様々な実験が行われている。現象と して,運動刺激へ数十秒順応させた後では,続くテスト 刺激の呈示時間は,実際の物理的時間よりも過小に評価 される(Burr, Chicchini, Arrichi, & Morrone, 2011; Burr, Tozzi, & Morrone, 2007; Johnston, Arnold, & Nishida, 2006)。また Johnston et al. (2006)によれば,高頻度・高速度の刺激 Copyright 2015. The Japanese Psychonomic Society. All rights reserved. * Corresponding author. Graduate School of the Humanities,
Major in Psychology, Senshu University, 2–1–1 Higashimita, Tama-ku, Kawasaki-shi, Kanagawa 214–8580, Japan. E-mail: [email protected]
(20 Hz; 20 deg/s)へ順応させた場合の方が,低頻度・低 速度の刺激(5 Hz; 5 deg/s)への順応の場合よりもテス ト刺激の時間過小評価量は大きい。一連の研究により, 運動刺激への順応の影響が同網膜位置上に限局し,テス ト刺激の知覚速度にかかわらず時間の過小評価が生じる ことから,視覚情報処理過程における初期段階の神経活 動のレベルでの時間知覚への関与が示唆された(Ayhan, Bruno, Nishida, & Johnston, 2009; Bruno, Ayhan, & Johnston, 2010; Johnston et al., 2006)。 他 方 で,Burr et al. (2007), Burr et al. (2011)は,順応による時間過小評価には少な くとも2つのメカニズムが関与するとしている。1つめ は網膜座標系において見かけの速度が影響する系であ り,2つめは外空間座標系で順応が影響する系である。 Burr et al. (2007)は,順応後のテスト刺激の見かけの速 度をprobe刺激と等しくなるように補正した場合,順応 刺激と網膜上で同位置のテスト刺激では時間過小評価が 生じず,外空間上で同位置のテスト刺激では時間過小評 価が生じることを示した。この結果から彼らは,順応に よる影響は,視覚処理過程における網膜座標系から外空 間座標系へと変換された後のより高次の処理過程での神 経機序において生じていると結論した。ところが,Bru-no et al. (2010)は,Burr et al. (2007)と同様の手続きで 実験を行い,外空間座標系における順応の影響はみられ ず,網膜座標系でのみ順応の影響が生じたとする相反す る結果を示しており,見解は一致していない。座標系の 視点を離れて,順応刺激とテスト刺激の関係に注目した Bruno, Ng, & Johnston (2013) や Curran & Benton (2012) は,順応刺激とテスト刺激の運動の向き関係を操作する 実験を行い,運動方向選択的な順応による影響を示し た。この結果からBruno et al. (2013)は,時間周波数依 存の低次の視覚情報処理レベルと,刺激の運動方向に依 存する高次の視覚処理レベルでの2つの関与が存在する ことを指摘している。時間知覚変化の機序に関しては異 なった説明を行ってはいるものの,これらの研究は, 順応によって時間が過小評価されるとする点では一致 している。しかしながら,順応によって時間が過大評価 されるとする研究も存在する。Ortega, Guzman-Martinez, Grabowecky, & Suzuki (2012)は,フリッカする正弦波状 の刺激へ数秒間順応させると,その後に同じ傾きの静止 刺激を呈示したときに,静止刺激の呈示時間が実際より も過大評価されることを示した。この結果からOrtega et al. (2012)は,方位選択性を持つV1野での符号化が時間 知覚変化に関与しているとした。 以上のように,順応による時間知覚の変化に関する研 究においては,時間知覚のひずみの表れ方が過小評価で あったり過大評価であったりと,一見矛盾した結果が用 いられており,その機序となっている視覚情報処理過程 についても統一される結論が得られているとは言えな い。これらの矛盾を解消して統一的な結論に至るために は,まずそれぞれの先行研究の違いを明確にする必要が ある。時間過小評価を示したBruno et al. (2010)やBurr et al. (2007)の研究と,時間過大評価を示したOrtega et al. (2012)の研究との実験手続き上の違いとしては,順 応刺激が運動する刺激かフリッカする刺激かという点に 加えて,順応刺激の呈示時間にも違いがみられる。Bru-no et al. (2010)やBurr et al. (2007)が用いた順応時間は 20–50 sで比較的長いものであり,過大評価を報告した Ortega et al. (2012)の研究で用いられた順応時間は5 sと それより短い。順応時間の違いが順応後の応答の変化に 影響することは,神経生理学的研究によって示されてお り(Fang, Murray, Kersten, & He, 2005; Henson, 2003),無 視できない要因である。Fang et al. (2005)は,fMRIを用 いて方位への選択的順応時の視覚処理段階ごとの血流量 変化を検討している。結果として,長い順応時間(25 s) では,V1からV4にわたる処理段階すべてにおいて方位 選択的な順応の影響がみられたものの,短い順応時間 (1 s)では,V3A, V4でのみ方位選択的な順応による影 響がみられたことから,順応による応答の変化が順応時 間に依存するとした。これは,V3A, V4といった高次皮 質では1 s程度の順応でも影響が見られるが,V1野等の 低次皮質では25 s程度の順応時間が応答の変化には必要 であるということであり,視覚情報処理過程の各レベル での方位への順応に要する時間が異なっていることを示 唆しているといえる。運動への順応においても,視覚情 報処理段階ごとに順応に必要な時間が異なっているとす れば,Bruno et al. (2010)やBurr et al. (2007), Ortega et al. (2012)の順応による時間知覚への影響の違いは,順応
時間の長短によって,視覚情報処理過程の異なる段階か らの影響が生じた結果によるという可能性を考えること ができる。
先行研究で用いられた順応時間の長さは,長いもので 20–50 s (Bruno et al., 2010; Burr et al., 2007; Burr et al., 2011),中程度のもので5 s (Ortega et al., 2012)などを挙 げることができるが,それより短い順応時間を用いたも のとしては,Bruno & Johnston (2010)の1.5 s程度しか見 当たらない。Bruno & Johnston (2010)の研究は,順応刺 激のコントラストの高低に焦点を当てたものであり,運 動刺激への短時間順応による影響を詳細に検討したもの ではない。つまり,長・中程度の順応時間での時間知覚 に関する知見は豊富なのに対して,短時間の順応につい
ては,順応刺激の時間周波数あるいは速度によって影響 がどのように異なるのか,順応刺激とテスト刺激の運動 方向の関係によって影響が変化するか否かなど実験的な 検討はなされていないのである。短時間順応と長時間順 応では,知覚時間に影響する処理段階が異なっている可 能性があることから,「短時間」の順応が,順応刺激と テスト刺激の運動方向といった要因とどのような相互作 用の下に時間知覚に影響を及ぼすのかを調べることは, 未だ統一的な結論に至っていない視覚的順応による時間 知覚機序の検討に大きな意味を持つ。 2. 目 的
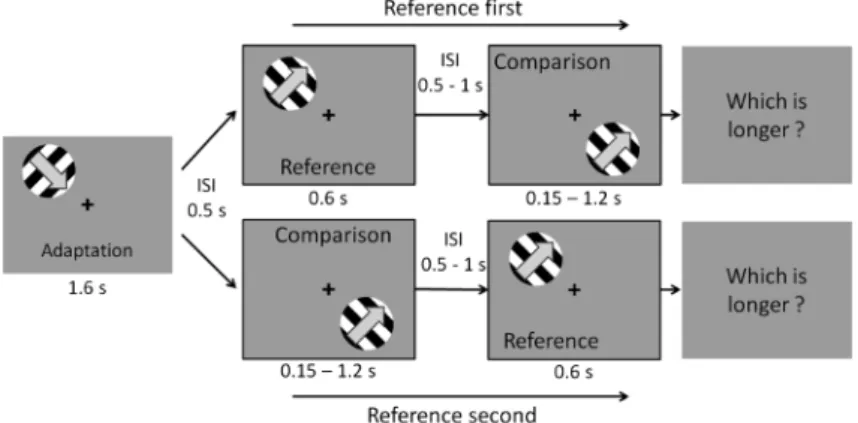
短い順応時間として,Bruno & Johnston (2010)で用い られた順応時間に近い 1.6 sの順応時間を設定し,順応 刺激の時間周波数または速度,続くテスト刺激との運動 方向の関係といった要因も含めて,短時間の運動刺激へ の順応の場合,時間知覚にどのような影響を与えている のかを検討する。 3. 方 法 3.1. 参加者 正常視力もしくは矯正正常視力を持つ大学生35名が 参加した。 3.2. 装置 刺激の呈示には,gamma補正を行ったCRTカラーモ ニタ(NANAO Flex Scan, 88F, 55 cm, 21型)を使用した。 解 像 度 は 1024×768 pixel で, リ フ レ ッ シ ュ レ ー ト は 60 Hzであった。刺激の制御と反応の取得には,デスク トップ型のパーソナルコンピュータとキーボードを使用 し, 刺 激 の 作 成 は Psychtoolbox 3.0 (Kleiner et al., 2007; Pelli, 1997)を導入したMATLABにて行った。実験時の 参加者の視距離を57 cmに固定するため,あご台を用い た。 3.3. 刺激・手続き Figure 1 に 実 験 1 試 行 の 流 れ を 示 す。 刺 激 図 形 は, Michelson contrast 100%で輝度変調させた空間周波数 1 c/degの正弦波縞を運動させ,これを円形窓(半径6.6 deg)に切り取ったものを用いた。円形窓の輪郭はガウ シアンフィルタでぼかし,灰色の背景(実測輝度 44.2 cd/m2)上に描画した。Michelson contrastをもとに算出 された正弦波縞図形の平均輝度は44.1 cd/m2であった。 この図形によって,順応刺激,参照刺激,比較刺激の3 種類の刺激を定義した。順応刺激は,2種類の時間周波 数5 Hzと20 Hzで変調させた。このときの正弦波縞の運 動速度はそれぞれ5, 20 deg/sとなる。参照刺激と比較刺 激の時間周波数は10 Hzとしたため,正弦波縞の運動速 度は10 deg/sとなる。刺激図形は,中央の注視点から画 面4隅に向かって偏心度8 degの4つの位置のいずれかに 円形窓の中心がくるように呈示した。1試行中の順応刺 激と参照刺激は常に同位置に呈示し,この位置に対して 画面中心に対して点対称の位置(画面対角線上)に比較 刺激を呈示した。順応刺激の呈示位置は4つの位置でカ ウンターバランスをとった。 実験は暗室中で行われ,参加者は,画面中央の黒色の 注視点を凝視し,自身でキーを押して試行を開始した。 キー押し後,まず順応刺激が 1.6 s呈示され,順応刺激 の消失後 0.5 s間,注視点の色が赤色に変化した。この 後,参照刺激と比較刺激のいずれかが先となり,2つの 刺激が順に呈示された。参照刺激が先に呈示される条件 を参照first条件,比較刺激が先に呈示される条件を参照 second 条件と呼ぶことにする。参照刺激の呈示時間は 0.6 sで固定し,比較刺激の呈示時間は0.15 sから1.2 sま
Figure 1. A trial is illustrated schematically. The Task was comparing durations of two stimuli (reference and comparison), and reporting which one is longer.
で0.15 s刻みで8種類とした。また,参照刺激と比較刺 激の刺激間間隔(ISI)は,0.5–1 sで試行ごとにランダ ムとした。 正弦波縞の運動方向は,参照刺激と比較刺激では常に 一致させた。順応刺激の運動方向は水平から 45, 135, 225, 315°の4方向から試行ごとにランダムに選択された。 順応刺激の運動方向に対して,参照・比較刺激の運動方 向が,直交する向きに運動する直交条件,同じ向きに運 動する同向き条件,反対向きに運動する反対向き条件を 設けた。加えて統制条件として,運動せず静止している 順応刺激静止条件,順応刺激を呈示せず 1.6 sのブラン クとする順応刺激非呈示条件を設けた。順応刺激静止条 件での順応刺激と参照・比較刺激は同一方位であった。 参加者の課題は,注視点の色が赤色に変わった後の2 つの刺激(すなわち,参照刺激と比較刺激の2つ)のう ち,どちらが呈示された時間が長いと感じたかを2件法 のキー押しで反応することであった。反応キーを押すと 1試行が終了し,次の試行へ移行した。 順応刺激と参照・比較刺激の運動方向関係は参加者間 要因であり,直行,同向き,逆向き条件に割り当てられ た参加者には,参加者内要因として,順応刺激時間周波 数 2 通り(5, 20 Hz)×比較刺激呈示時間 8 通り(0.15– 1.2 s)×参照・比較刺激呈示順序 2 通り(参照 first, 参照 second)からなる32試行を1セットとして,参加者1人 につき10 セットの計 320 試行を行わせた。順応刺激静 止,順応刺激非呈示条件に割り当てられた参加者には, 前述の参加者内要因から順応刺激時間周波数の要因を除 いた計160試行を行わせた。参加者間要因である運動向 き関係には,各条件に参加者7名ずつを無作為に割り当 てた。各試行の呈示順序は参加者ごとにランダムとし た。なお,1セット終了ごとに参加者は任意の休憩をと ることが可能で,休憩終了後に参加者はキーを押すこと で実験を再開した。 4. 結 果 各条件において,参加者が参照刺激よりも比較刺激の 方が長い時間呈示されたと反応した比率を求め,MAT-LAB上で累積正規分布関数によるフィッティングを行っ た。フィッティングした曲線において,反応比率が 50%となる比較刺激呈示時間を,参照刺激の物理的な呈 示時間(0.6 s)に対するその参加者の主観的等価点 (PSE)とした。参照刺激の物理的な呈示時間(0.6 s)を 正確に知覚できていた場合,PSEは0.6 sと等しくなる。 PSEが0.6 sより大きい場合は参照刺激の呈示時間の過大 評価を,小さい場合には過小評価を意味する。PSEの全 体平均は0.59 s (SD=0.05)であった。 短時間順応下でも,運動刺激への順応によって時間知 覚が変化していたか,すなわち順応刺激の有無によって 時間知覚が異なるかを検討する。Figure 2に順応刺激の 条件ごとの平均PSEを示した。横軸は順応刺激静止・順 応刺激非呈示の統制条件を含めた運動向き関係を表し, 各条件での平均PSEの値が示されている。ここでは順応 刺激時間周波数の条件は無視してある。運動刺激へ短時 間順応させた条件と,運動刺激への順応がない順応刺激 静止条件および順応刺激を呈示しない順応刺激非呈示条 件の2つの統制条件とで比較を行うため,参照・比較呈 示順序2水準(参照first,参照second)×統制条件を含む 運動向き関係5水準(直交・同向き・反対向き・順応刺 激静止・順応刺激非呈示)の2要因分散分析を行った。 その結果,運動向き関係と,参照・比較呈示順序に有意 な主効果は認められなかったが(F(1, 30)=3.65, p=.07; F(4, 30)=0.43, p=.79), 交 互 作 用 は 有 意 で あ っ た (F(4, 30)=2.78, p<.05)。参照 first,参照 second 条件そ れぞれについて,順応刺激非呈示条件を対照群とした Dunnettのt検定を行ったが,参照first条件における逆向 き条件と順応刺激非呈示条件間,参照 second条件にお ける同向き条件と順応刺激非呈示条件間での差に有意な 傾向が見られたものの(M=0.11, p=.06; M=.14, p=.05), 全体として,各条件と順応刺激非呈示条件の間に有意な 差は認めらなかった(ps>.05)。
長時間の順応を用いた Johnston et al. (2006)やBruno et al. (2013)では,順応刺激と続くテスト刺激の運動向 き関係や,順応刺激の時間周波数の違いによって,時間 評価量が変化することを示した。短時間順応下でも同様 Figure 2. Average PSEs are plotted as a function of
mo-tion direcmo-tion condimo-tions (including two control con-ditions). Error bars show standard error (SE).
の影響が生じるかを検討するため,運動向き関係,順応 刺激時間周波数,参照・比較呈示順序条件によるPSEの 値を比較検討する。各条件における平均 PSEをFigure 3 に示した。横軸は順応刺激と参照・比較刺激の運動向き 関係を表し,(a)は参照first条件の,(b)は参照second 条件の PSEを表している。運動向き関係3水準(直交・ 同向き・反対向き)×順応刺激時間周波数 2水準(5・ 20 Hz)× 参 照・ 比 較 呈 示 順 序 2 水 準(参 照 first・ 参 照 second)の3要因分散分析を行ったところ,全ての 要因について主効果も交互作用にも有意性は認められ なかった(F(2, 18)=0.30, p=0.74; F(1, 18)=1.30, p=0.27; F(1, 18)=0.14, p=0.72; 全交互作用においてps>.05)。 続いて参照・比較呈示順序の影響を検討する。順応の 影響が,同位置に呈示された刺激の時間知覚にのみ限 定されるとする先行研究(Ayhan et al., 2009; Bruno et al., 2010; Burr et al., 2007; Johnston et al., 2006)に従えば,参 照firstのPSEは,順応直後すなわち順応の効果が最大で あると考えられるときの参照刺激の呈示時間評価であ り,これに対して参照secondのPSEは,順応刺激消失後 の時間経過によって順応の効果が減衰している可能性が ある状況下での参照刺激の呈示時間評価ということにな る。参加者ごとに,参照first条件のPSEから参照second のPSEを減じた差分PSEの指標を導入すれば,これは, 順応の効果の時間的推移,あるいは効果の減衰量を示す 指標として解釈することができる。次に,順応刺激非呈 示条件の差分PSEを考える。例えば,先に呈示された刺 激をより長いと判断するならば,参照first条件でPSEが 大きくなり,参照second条件でPSEが小さくなるため, 差分PSEは正の値をとることになる。これは,判断バイ アスと解釈することができる。つまり,順応刺激が呈示 される条件における差分PSEは,この判断バイアスに, 順応刺激による効果の時間的推移を可算したものと考え ることができる。条件間で差分PSEが異なることは,条 件によって順応効果の時間的推移が異なることを意味 し,異なる視覚処理過程における順応であることを示唆 することになる。Figure 4に各条件における平均差分 PSEの値を示した。条件による差分PSEの違いを検討す るため,運動向き関係5水準の1要因分散分析を行った ところ,有意な差が認められた(F(4, 30)=2.78, p<.05)。 順応刺激非呈示条件を対照群としたDunnettのt検定を 行った結果,順応刺激非呈示条件よりも,同向きおよび 逆向き条件での差分 PSE が有意に低かった(M=.23; M=.24, ps<.05)。順応刺激非呈示条件と直交条件,順 応刺激非呈示条件と順応刺激静止条件間には有意な差は 認められなかった(ps>.05)。 5. 考 察 本研究は,短時間の運動刺激への順応によって,知覚 時間がどのように変化するかを検討した。1.6 sの短時間 Figure 3. Average PSEs are plotted as a function of
mo-tion direcmo-tion condimo-tions without static adaptamo-tion and no adaptation conditions. (a) PSEs indicate reference first condition. (b) PSEs indicate reference second condition. Dashed lines indicate the duration of the reference stimulus. Error bars show standard error (SE).
Figure 4. Average Difference PSEs are plotted as a function of motion direction conditions. Error bars show standard error (SE).
順応下での参照刺激の呈示時間の評価は,順応を行わな い順応刺激非呈示条件と比較して,顕著なPSE (時間評 価量)の違いは見られなかった(Figure 2)。また,本研 究での順応刺激とテスト刺激の時間周波数は,長時間の 順応による時間評価研究(Bruno et al., 2010; Johnston et al., 2006)にならい設定したが,順応刺激の時間周波数 もしくは速度の高低や,順応刺激とそれに続く参照・比 較 刺 激 の 運 動 向 き 関 係 に よ っ て も 違 い は 見 ら れ ず (Figure 3),長時間順応によって時間過小評価が生じた とする研究(Bruno et al., 2010; Bruno et al., 2013; Burr et al., 2007; Johnston et al., 2006)や,中程度の順応時間(5 s) において時間の過大評価が生じたとする研究(Ortega et al., 2012)とも異なる結果が示された。PSEの条件間の直 接比較では差が認められなかったが,参照刺激と比較刺 激の呈示順序によるPSEの違いを,本研究では差分PSE という指標によって検討したところ,統制条件の順応刺 激非呈示条件と,同向き・逆向き条件で異なる傾向が見 られた(Figure 4)。差分PSEにおいて条件間の違いがみ られたことは,単純な条件間のPSEの比較からでは得ら れなかった短時間順応の時間知覚への影響の存在を示し ていると考えることが可能である。 今回の結果が生じた要因について検討する。まず,有 意な時間評価量の変化が見られなかったことに関して は,運動刺激への順応時間が短かったために,後続する 参照刺激の時間評価に対する影響が小さかったという可 能性を考えることが可能である。傾きへの順応後の視覚 処理皮質の活動量を,順応時間を変化させて検討した Fang et al. (2005)の研究においても,全体の活動量は, 短時間の順応条件に比べて,長時間の順応条件でより大 きな変化が見られていた。運動刺激への順応状況であっ ても,順応時間が短いことによって,時間知覚へ関与す る視覚情報処理過程への影響が比較的小さいために,有 意な時間評価量の違いとして表れなかった可能性はあ る。 続いて,有意な差が生じた差分 PSEについて検討す る。各条件におけるPSEの違いという先行研究に従う指 標に対して,本研究において用いた差分PSEは,参照・ 比較刺激を順応刺激と同方位(同向き,逆向き)で呈示 した場合に,順応刺激非呈示時より有意に低い値を示し た。順応刺激非呈示条件における差分PSEは正の値を とっており,本研究では,先に呈示された刺激がより長 い時間呈示されたと判断されていたことになる。これ は,刺激のコントラストや大きさ,刺激間間隔によら ず,先に呈示された刺激の呈示時間が,それ以降に呈示 された刺激の呈示時間よりも過大評価されるとする先行
研究(Rose & Summers, 1995)の知見に合致している。 加えて順応刺激条件においては,順応刺激を呈示するこ とによって,判断バイアスのほかにも差分PSEを変動さ せるいくつかの影響の可能性を考えることができる。例 えば,順応刺激が参照刺激や比較刺激より先に呈示され たことそれ自体による呈示順序バイアスの影響,1.6 sと いう順応刺激の固定された呈示時間による手掛かり的影 響,順応刺激の呈示位置によって,参照刺激や比較刺激 の呈示位置が予測可能であることによる刺激呈示位置へ の注意シフトの影響などである。呈示順序バイアスと類 似した影響として,Bruno et al. (2010)は,呈示順序に よる時間評価の違いが呈示時間判断の方略が異なること で生じるという考察を行っている。このような様々な影 響の可能性が考えられるが,それぞれの影響の大きさ は,順応刺激が呈示される条件においては一定であると 推定される。つまり,順応刺激が呈示された条件間での 比較に際してはこれらの影響を無視することができる。 本研究での差分PSEでは,順応刺激非呈示条件を基準と した場合,同向き条件と逆向き条件との間でそれぞれ有 意な差が検出されたが,直交条件や静止条件との間には 有意な差は見られなかった。これは,少なくとも本研究 における実験状況下では,先に挙げた差分PSEに反映す る可能性のある様々な要因による影響は,たとえ存在し たとしても,同向き・逆向き条件における順応の影響に 比して大きいものではなかったということを示唆してい る。つまり,同向き・逆向き条件という順応刺激とテス ト刺激の方位一致条件下において,短時間の運動刺激へ の順応による時間知覚への影響が存在することが間接的 にではあるが示唆されたと考えることができる。また, 静止した順応刺激と参照・比較刺激の方位が一致してい た順応刺激静止条件の差分PSEが,順応刺激非呈示条件 の差分PSEと有意な差がなかったという結果は,短時間 順応においては運動刺激への順応が必要である可能性を 示している。ただし本研究においては,比較刺激の呈示 時間が 0.15–1.2 sで変化する上に,参照刺激と比較刺激 間の刺激間間隔は 0.5–1 s でランダムであり,刺激の SOAは不定となっている。そのため,今後はSOAを統 制した実験条件を設定したうえで,厳密に順応効果の時 間的推移を検討する必要があるものの,少なくとも本研 究の結果は,方位一致時に特異的な運動刺激への順応の 影響の存在を示しているといえる。 本研究と同様に短時間の順応下での時間知覚への影響 を検討した Bruno & Johnston (2010)は,刺激の向きが 直交関係であるにもかかわらず,順応刺激とテスト刺激 の時間周波数が10 Hz以上であるときに,低コントラス
ト(10%)順応時に比べて,高コントラスト(90%)順 応時のテスト刺激が過小評価されることを示し,外側膝 状体の大細胞系路におけるコントラスト順応の影響が, 知覚時間変化の要因であるとした。本研究では,コント ラストを十分な高さで一定としたが,順応刺激とテスト 刺激の向き関係について,順応刺激が5 Hzであっても 20 Hzであっても,直交条件においては時間過小評価は 生じず,同向き・逆向きの方位一致条件下においてのみ 知覚時間に影響が生じていた。この結果は,短時間の順 応状況下において,Bruno & Johnston (2010)が示した要 因以外の時間知覚への影響要因が存在することを示唆し ているといえる。また,Bruno et al. (2013)は,向きも しくは方位に依存し,広い範囲の時間周波数への順応に よって生じる皮質レベルでの時間知覚変化と,向きや方 位によらず高時間周波数への順応によって生じる皮質下 での時間知覚変化の2つの機序が存在することを示唆し た。本研究では,順応刺激の時間周波数の高低によら ず,方位選択的な順応による影響がみられたことから, 皮質レベルでの処理過程による時間知覚変化に対応する といえる。さらに,短時間の順応によって応答が変化す るのは高次皮質であること(Fang et al., 2005)から,高 次の皮質での処理過程による影響の可能性がある。しか し,本研究と同じく方位選択的な順応の影響が存在する としたOrtega et al. (2012)は,低次皮質の V1レベルの 影響であるとしている。彼らは,運動のないフリッカ刺 激への順応による影響が方位選択的であることをもっ て,最初に方位選択的な応答が見られるV1が時間知覚 変化に寄与すると指摘したが,複数処理レベルでの順応 の影響を考慮していないため,V1以降での方位選択的 な影響によって時間知覚が変化していた可能性は否定で きない。また, Ortega et al. (2012)が用いた順応刺激は フリッカ刺激であり,運動刺激への順応ではなかったこ とや,Johnston et al. (2006)の実験条件においては方位 選択的な影響が見られなかったことから,Ortega et al. (2012)はフリッカ刺激への順応による時間過大評価と, 運動刺激への順応による時間過小評価は異なる機序によ るものであると指摘しており,連続的な運動への順応と 向きのない頻度情報への順応の違いについては,より詳 細な検討が必要だといえる。 本研究では,運動刺激への短時間順応による時間知覚 への影響の様相を明らかにした。短時間順応下での本研 究,中程度の時間の順応下での研究(Ortega et al., 2012), 長時間順応下での研究(Bruno et al., 2010, 2013; Burr et al., 2011)などの様々な順応時間による時間知覚の変容の違 いは,順応時間の長短によって異なる過程の影響が反映 さ れ た も の で あ る可 能 性 が あ る。 例 え ば Burr et al. (2011)は,運動刺激への順応が視覚情報処理過程の複 数レベルで生じ,それぞれのレベルでの時間知覚への影 響が混在している可能性を指摘している。実験における 順応時間を網羅的に変化させていった場合の知覚時間を 比較し,それぞれの視覚処理段階における順応の最適時 間と知覚時間を対応させることで,混在する影響を分離 できる可能性を検討していくことが,今後の課題となる であろう。また,本研究では順応効果の時間的推移,減 衰量という情報を利用できる可能性を示した。これは, 複数の視覚処理段階における時間知覚への順応の影響を 分離し,その神経基盤を明らかにするうえで有用であ る。これまでの研究では,順応による時間評価の変化量 に主眼を置いていたが,順応による影響が持続する場合 と,急速に減衰してしまう場合とでは,時間知覚変化の 基盤となる神経機序が異なっている可能性がある。この 可能性を検討するため,順応刺激と標準刺激の時間間隔 を体系的に変化させるような実験を行い,検討していく 必要があるだろう。 引用文献
Ayhan, I., Bruno, A., Nishida, S., & Johnston, A. (2009). The spatial tuning of adaptation-based time compression.
Jour-nal of Vision, 9(11), 1–12.
Brainard, D. H. (1997). The Psychophysics Toolbox. Spatial
Vision, 10, 443–446.
Bruno, A., Ayhan, I., & Johnston, A. (2010). Retinotopic adap-tation-based visual duration compression. Journal of Vision,
10(10), 1–18
Bruno, A., & Johnston, A. (2010). Contrast gain shapes visual time. Frontiers in Psychology, 1(170), 1–8.
Bruno, A., Ng, E., & Johnston, A. (2013). Motion-direction specificity for adaptation-induced duration compression depends on temporal frequency. Journal of Vision, 13(12), 1–11.
Burr, D. C., Cicchini, G. M., Arrighi, R., & Morrone, M. C. (2011). Spatiotopic selectivity of adaptation-based compres-sion of event duration. Journal of Vicompres-sion, 11(2), 1–9. Burr, D. C., Tozzi, A., & Morrone, M. C. (2007). Neural
mech-anisms for timing visual events are spatially selective in re-al-world coordinates. Nature Neuroscience, 10, 423–425. Curran, W., & Benton, C. P. (2012). The many directions of
time. Cognition, 122, 252–257.
Fang, F., Murray, S. O., Kersten, D., & He, S. (2005). Orienta-tion-tuned FMRI adaptation in human visual cortex.
Jour-nal of Neurophysiology, 94, 4188–4195.
Henson, R. N. (2003). Neuroimaging studies of priming.
Prog-ress in Neurobiology, 70, 53–81.
Johnston, A., Arnold, D. H., & Nishida, S. (2006). Spatially lo-calized distortions of event time. Current Biology, 16, 472–
479.
Kaneko, S., & Murakami, I. (2009). Perceived duration of visu-al motion increases with speed. Journvisu-al of Vision, 9(7), 1–12. Kleiner, M., Brainard, D., Pelli, D., Ingling, A., Murray, R., &
Broussard, C. (2007). What’s new in Psychtoolbox-3.
Per-ception, 36(14), 1.
Mark, M. D., & Buonomano, D. V. (2004). The neural basis of temporal processing. Annual Review of Neuroscience, 27, 307–340.
松田文子・調枝孝治・甲村和三・神宮英夫・山崎勝之・ 平 伸二 (編著) (1996).心理的時間その広くて深い なぞ 北大路書房 pp. 38–48.
(Matsuda, F., Choshi, K., Kohmura, K., Jingu, H., Yamasaki,
K., & Hira, S.)
Ortega, L., Guzman-Martinez, E., Grabowecky, M., & Suzuki, S. (2012). Flicker adaptation of low-level cortical visual neu-rons contributes to temporal dilation. Journal of
Experimen-tal Psychology: Human Perception and Performance, 38,
1380–1389.
Pelli, D. G. (1997). The VideoToolbox software for visual psy-chophysics: Transforming numbers into movies. Spatial
Vi-sion, 10, 437–442.
Rose, D., & Summers, J. (1995). Duration illusions in a train of visual stimuli. Perception, 24, 1177–1187.