九州大学学術情報リポジトリ
Kyushu University Institutional Repository
C型レクチン受容体を介する病原性真菌マラセチアの 認識機構と免疫応答に関する研究
石川, 哲章
https://doi.org/10.15017/1398540
出版情報:Kyushu University, 2013, 博士(医学), 課程博士 バージョン:
権利関係:Public access to the fulltext file is restricted for unavoidable reason (2)
1
Identification of distinct ligands for the C-type lectin receptors Mincle and Dectin-2
in the pathogenic fungus Malassezia
Running title: Fungal ligands for Mincle and Dectin-2
Tetsuaki Ishikawa1, Fumie Itoh2, Sayumi Yoshida2, Shinobu Saijo3,4, Tetsuhiro Matsuzawa5, Tohru Gonoi5, Takashi Saito6,7, Yoshio Okawa2, Nobuyuki Shibata2,
Tomofumi Miyamoto8* and Sho Yamasaki1*
1 Division of Molecular Immunology, Medical Institute of Bioregulation, 8 Department of Natural Products Chemistry, Graduate School of Pharmaceutical Sciences, Kyushu University, 3-1-1 Maidashi Higashi-ku, Fukuoka 812-8582, Japan
2 Department of Infection and Host Defense, Tohoku Pharmaceutical University, 4-4-1 Komatsushima Aoba-ku, Sendai 981-8558, Japan
3 Department of Molecular Immunology, 5 Department of Bio-resources, Medical Mycology Research Centre, Chiba University, 1-8-1 Inohana Chuo-Ku, Chiba 260-8673, Japan
4 PRESTO, Japan Science and Technology Agency (JST), Saitama 332-0012, Japan.
6 Laboratory for Cell Signaling, RIKEN Research Center for Allergy and Immunology, Yokohama 230-0045, Japan
7 WPI Immunology Frontier Research Center, Osaka University, Suita, 565-0871, Japan
Correspondence should be addressed to Tomofumi Miyamoto or Sho Yamasaki e-mail address; [email protected] or [email protected] Phone number: +81-92-642-6636 or +81-92-642-4614
Fax number: +81-92-642-6636 or +81-92-642-4614 http://dx.doi.org/10.1016/j.chom.2013.03.008
2 SUMMARY
Various C-type lectin receptors (CLRs), including Mincle and Dectin-2,
function as pattern-recognition receptors and play a central role in immunity to fungal
pathogens. However, the precise structures of the CLR ligands in various pathogenic
fungi have yet to be defined. Here we report that Malassezia, an opportunistic skin
fungal pathogen, is cooperatively recognized by Mincle and Dectin-2 through distinct
ligands. Solvent-based fractionation revealed that Mincle and Dectin-2 recognize
lipophilic- and hydrophilic-components of Malassezia, respectively. Mass spectrometry
and NMR revealed glyceroglycolipid and unique mannosyl-fatty acids linked to
mannitol as two Mincle ligands. An O-linked mannobiose-rich glycoprotein was
identified as a Malassezia ligand for Dectin-2. Cytokine production in response to the
Mincle-ligands and the Dectin-2-ligand was abrogated in Mincle–/– and Dectin-2–/–
dendritic cells, respectively. These results demonstrate that Mincle and Dectin-2
recognize distinct ligand in Malassezia to induce host immune responses.
3 Highlights
Mincle and Dectin-2 cooperatively recognize Malassezia through distinct
ligands
Mincle ligands in Malassezia are glucosyl-glycolipid and mannosyl-glycolipid
Dectin-2 ligand in Malassezia is O-linked mannobiose-rich glycoprotein
Cytokine response to the respective ligands was impaired in Mincle–/– and
Dectin-2–/– DCs
4 INTRODUCTION
Innate host response is mediated by pattern recognition receptors (PRRs) for
pathogen-associated molecular patterns (PAMPs), including Toll-like receptors (TLRs),
retinoic acid-inducible gene-I-like receptors (RLRs), nucleotide-binding
oligomerization domain-like receptors (NLRs) and C-type lectin receptors (CLRs)
(Robinson et al., 2006; Takeuchi and Akira, 2010). Among these PRRs, CLRs play a
crucial role in recognizing the complex structures, composed of carbohydrate residues,
of various fungal pathogens (Hardison and Brown, 2012; Robinson et al., 2006).
Recently, some CLRs have also been demonstrated to directly transduce the signals to
produce inflammatory cytokines through signaling subunits such as FcR (Robinson et
al., 2006), however, the ligands of most CLRs remain largely unclear.
Mincle (also called Clec4e or Clecsf9) is a CLR expressed in activated
macrophages and dendritic cells (DCs) subjected to several types of stress (Matsumoto
et al., 1999). We have previously shown that Mincle is an activating receptor coupled
with Fc receptor (FcR) chain, an immunoreceptor tyrosine-based activation motif
(ITAM)-containing adaptor (Yamasaki et al., 2008). We found that Mincle recognizes
5
mycobacteria, and identified trehalose-6'6-dimicolate (TDM, also called cord factor) as
a specific ligand (Ishikawa et al., 2009). TDM is a glycolipid derived from the
mycobacterial cell wall, and it has been shown to possess potent adjuvant activity
(Hunter et al., 2006). Although the TDM expression is restricted to mycobacteria,
corynebacteria and Nocardia, we have recently found that Mincle also recognizes
Malassezia species (Yamasaki et al., 2009). However, the Mincle ligand in Malassezia
has not yet been identified, thus suggesting the existence of a ligand other than TDM in
this fungus.
Fungi of the Malassezia genus are found in the normal flora of human skin.
These species are considered to be harmless commensal organisms under normal
circumstances, however, they are also widely known as opportunistic pathogens
(Gaitanis et al., 2012; Guillot and Bond, 1999). They have been reported to be
associated with diverse dermatological pathologies, including pityriasis versicolor,
seborrheic dermatitis, atopic dermatitis and folliculitis (Ashbee and Evans, 2002;
Guillot and Bond, 1999). Malassezia species also cause lethal systemic infections in
newborn infants receiving intravenous lipid emulsions (Marcon and Powell, 1992;
6
Redline and Dahms, 1981). Intriguingly, Malassezia species are unique among other
fungi in that they require lipid for their growth (Schmidt, 1997). However, the
mechanism underlying the recognition of Malassezia by host cells has not been fully
elucidated.
Dectin-2 (also called Clec4n) is another FcR-coupled CLR that is
constitutively expressed on DCs, tissue macrophages and inflammatory monocytes
(Sato et al., 2006; Taylor et al., 2005). Dectin-2 is reported to recognize a variety of
fungi, including Candida albicans, Saccharomyces cerevisiae, Histoplasma capsulatum,
Microsporum audouini and Trichophyton rubrum (McGreal et al., 2006; Ritter et al.,
2010; Sato et al., 2006). It is therefore possible that Dectin-2 also recognizes Malassezia.
Dectin-2 is reported to bind to high mannose structures (McGreal et al., 2006; Sato et al.,
2006), especially mannans in C. albicans (Saijo et al., 2010), however, the precise
structure of the Dectin-2 ligand has not yet been fully defined.
In this study, we demonstrate that two CLRs, Mincle and Dectin-2,
cooperatively induce the immune response to the same fungus, Malassezia, through the
recognition of distinct ligands.
7 RESULTS
Dectin-2 recognizes Malassezia species
We first tried to compare the recognition property of Mincle and Dectin-2.
More than 45 species of pathogenic fungi were analyzed by using NFAT-GFP reporter
cells expressing FcR with Mincle or Dectin-2. As we have previously reported, Mincle
selectively recognizes Malassezia species (Figure S1A) (Yamasaki et al., 2009). In
sharp contrast, Dectin-2 broadly recognizes a variety of fungi, including Trichophyton,
Aspergillus, Cladosporium, Candida and Malassezia species (Figure 1). Interestingly,
only Malassezia species represented an overlapping target for both receptors. We
confirmed that Malassezia could not activate reporter cells expressing FcR alone (data
not shown).
Mincle and Dectin-2 recognize Malassezia by distinct mechanisms
In order to identify the ligand(s) for Mincle and Dectin-2 in Malassezia, we
tried to extract active fraction with various aqueous-organic solvents (Figure 2A). These
extracts and solvent-treated fungal cells were then tested to determine their ability to
8
stimulate NFAT reporter cells. We found that M. pachydermatis treated with
chloroform:methanol (C:M) selectively lost their Mincle-stimulating activity (Figure
2B). Simultaneously, we analyzed the activity of extracted fractions in plate-coated
form, and found that only the C:M phase after C:M extraction showed strong
stimulatory activity (Figure 2C). These findings suggested that Mincle recognizes some
lipophilic component(s) in Malassezia.
On the other hand, the activity for Dectin-2 was efficiently extracted into the
water phase, indicating that the Dectin-2 recognizes hydrophilic component(s) in
Malassezia (Figures 2D and 2E). These results suggest that Malassezia may possess
distinct ligand components that are recognized by two different CLRs, Mincle and
Dectin-2.
Identification of two glycolipids as Mincle ligands
We first tried to identify the Mincle ligand(s) in Malassezia. The C:M-soluble
fraction was separated into 49 fractions by silica gel column chromatography. Extracts
from these fractions showed strong ligand activity that peaked at fractions #44-45
9
(Figure 3A, top). A thin layer chromatography (TLC) analysis demonstrated that these
fractions (#44-45) contain several spots that were considered to be candidates for
Mincle ligands (Figure 3A, bottom). We further analyzed fraction #44 by means of high
performance thin layer chromatography (HPTLC) and separated it into 20 subfractions
to identify the active lipid components. Fraction #44 contained two peaks of ligand
activity, 44-1 and 44-2, corresponding to the position of two purple-red spots detected
by orcinol staining. These results showed that 44-1 and 44-2 contain glycolipids that are
different from TDM (Figure 3B). We further purified 44-1 and 44-2 from fraction #44
by reversed-phase column chromatography and HPLC and their chemical structures
were determined on the basis of the chemical and spectroscopic evidence using
FAB-MS, ESI-TOFMS, 1H NMR, 13C NMR, and GC-MS.
We first analyzed 44-1 structure (Figure S2A). The negative FAB-MS
spectrum of 44-1 showed a pseudo-molecular ion peaks [M-H]– at m/z 919. The
fragment ion peaks due to fatty acid anions were observed at m/z 297 [C18H37COO–]
and 241 [C14H29COO–] as shown in Figure 3C. The molecular formula of 44-1 was
further determined to be C49H92O15 by the ESI-TOFMS [m/z 943.6362, calcd. 943.6328
10
(M+Na)+] (Figure S3A). The 1H-NMR spectrum of 44-1 exhibited a strong broad signal
due to the aliphatic methylenes at H 1.22, a terminal methyl signal at H 0.80 and
several multiplets between H 3.80 and 4.80 due to oxygenated methine and methylene
protons. Two typical anomeric proton signals were also detected at H 4.97 (1H, d, J =
7.9 Hz) and H 4.78 (1H, d, J = 7.6 Hz), suggesting two -linked monosaccharides
(Figure S3B). The 13C-NMR spectrum exhibited two terminal methyl signals at C 11.3 and 19.1, aliphatic methylenes at C 25-35, fourteen oxygenated methylenes, methines
at C 62-78, two anomeric carbon signals at C 104.4 and 104.8, and two estercarbonyl
signals at C 173.3 and 173.4. These results indicated that 44-1-2 was a
glyceroglycolipid (Figure S3C).
The structure of the sugar and glycerol moiety was determined as follows.
The 1H-1H COSY and TOCSY spectra revealed the two independent correlations from
H-1 to H-6 of -glucopyranoses, and H-1 to H-3 of a glycerol. The connectivity of two
glucopyranoses and a glycerol was determined based on the HMBC correlations
between the H-1” (H 4.97) of -Glcp and C-6’ (C 69.6) of -Glcp, and between H-1’
(H 4.78) of Glcp and C-3 (C 68.1) of glycerol. The terminal structure of 3 branched
11
fatty acid was confirmed by the chemical shift value and HSQC correlations of terminal
methyl signals (Figures S3D-S3G) (Pan et al., 2010). Composition of the neutral sugar
and fatty acids was conducted by GC-MS analysis following methanolysis (Figures
S3H-S3J).
Taken together, the less polar glycolipid (44-1) was a glyceroglycolipid
having one glycerol, one gentiobiose (6-O--D-glucopyranosyl-D-glucopyranose), and
anteiso-fatty acids (C14 and C18), which are attached via ester bonds to the hydroxyl
groups of the glycerol backbone (Figure 3D and Figure S3K). In addition to 44-1, small
amount of related compounds that are similar to 44-1, except for the length of acyl
chains, were identified. We confirmed that all these compounds possess comparable
Mincle ligand activity (Figures S4A and S4B).
Interestingly, 44-1 has a structural similarity to the membrane anchor moiety
of lipoteichoic acid (LTA), a bacterial component recognized by TLR2 (Schwandner et
al., 1999). However, LTA did not act as a Mincle ligand (Figures S4C-S4E).
We next analyzed the structure of 44-2 (Figure S2B). The positive
ESI-TOFMS spectrum of 44-2 showed a pseudo-molecular ion peaks at m/z
12
1700.0501[M+Na]+ and gave molecular formula as C84H156O32Na (calcd. for
1700.0472) (Figure 3E). The 1H-NMR spectrum of 44-2 exhibited a typical spectrum
feature due to glycolipid (Figure S3L). Three anomeric proton signals at δH 4.96 (1H,
brs), 5.00 (2H, brs), and 5.43 (1H, brs) were assignable to -mannopyranose from the
correlations of 1H-1H COSY, TOCSY, NOESY and HSQC spectra (Figures S3M-S3P).
Because the molecular of 44-2 was so large to elucidate by only spectroscopic analysis,
the chemical conversion methods were used for structure elucidation. GC-MS analysis
following methanolysis revealed that 44-2 consists of D-mannose, mannitol and
10-hydroxystearic acid clearly (Figures S3Q-S3T). 1H-NMR, TOCSY, HSQC and
HMBC spectra of 44-2 at lower temperature clarified that the three acyl moieties were
linked to the position 1, 3, 4 on mannitol (Figures S3U-S3X). The treatment of 44-2
with 0.5 M NaOMe gave two glycosyl fatty acids and L-mannitol (Figures S4F-S4U).
The structure of two glycosyl fatty acids were determined to be
10-O--D-mannopyranosyl stearic acid methyl ester and
10-O--D-mannopyranosyl-(1->2)--D-mannopyranosyl stearic acid methyl ester using
Mass and NMR analysis, respectively. Taken together, the analysis on native form
13
revealed that the polar glycolipid (44-2) has a mannitol backbone, which is attached to
two 10-O--D-mannopyranosyl-10-hydroxy-stearic acids, and one
10-O-[-D-mannopyranosyl-(1-2)--D-mannopyranosyl]-10-hydroxy-stearic acid via
ester bonds (Figure 3F).
Thus, 44-2 is a Mincle ligand with unique structure that have not been
previously reported in nature or synthesized in the laboratory. Interestingly, only weak
activity was detected in glycoside components of 44-2 obtained by alkaline hydrolysis
(Figures S4F-S4U), suggesting that intact form of 44-2 structure is required for the
potent ligand activity.
Mincle is necessary and sufficient for the recognition of Malassezia glycolipid
ligands
The ligand activity of these glycolipids was verified using Mincle-expressing
reporter cells. 44-1 and 44-2 had a Mincle ligand activity as potent as TDM (Figure 4A).
We further confirmed that Mincle directly binds to these glycolipids by using soluble
Mincle-Ig protein (Figure 4B). These results indicate that Malassezia fungus possess
14 two Mincle ligands with unique structures.
To examine the contribution of endogenous Mincle as a receptor for these
glycolipids, we tested the ability of 44-1 and 44-2 to activate dendritic cells. Wild-type
bone marrow-derived dendritic cells (BMDCs) were able to secrete TNF in response to
44-1 and 44-2 (Figure 4C). This TNF production was almost completely suppressed in
Mincle–/– DCs, indicating that Mincle is an essential receptor for 44-1 and 44-2 in DCs
(Figure 4C). This finding also confirmed that the observed DC activation is not due to
possible contaminating TLR ligands in these fractions, since TLR signaling is intact in
Mincle–/– mice (Ishikawa et al., 2009). Dectin-2 was dispensable for the recognition of
these glycolipids, which is consistent with the results of reporter cells. FcR, a signaling
subunit of Mincle, was also essential for the response to the glycolipids, whereas
zymosan induced a similar response in all these cells (Figure 4D). We therefore
concluded that Mincle is an essential receptor for the cytokine production induced by
two glycolipids derived from Malassezia.
Dectin-2 recognizes Malassezia through -1,2-linked mannose
15
Dectin-2 contains a glutamic acid-proline-asparagine (EPN) motif, which is
known to preferentially bind to mannose (Drickamer, 1992). To assess whether the
recognition of Malassezia cells by Dectin-2 requires this motif, we substituted residues
of the EPN motif of Dectin-2 to create a glutamine-proline-asparagic acid (QPD) motif,
which is known as galactose-binding motif (Drickamer, 1992). M. furfur failed to
activate reporter cells expressing Dectin-2 mutant (Dectin-2QPD) (Figure 5A).
To determine the saccharide through which Dectin-2 recognizes Malassezia,
we tried to block the recognition with various kinds of monosaccharides. An excessive
amount of mannose was able to block the NFAT-GFP reporter activity induced by
Malassezia cells, whereas glucose and galactose did not show any blocking activity
(Figure 5B). These results suggest that Dectin-2 may recognize Malassezia through
mannose related structure.
A hydrophilic component has Dectin-2 ligand activity
We next searched for the Dectin-2 ligand in Malassezia. The Malassezia
water-soluble fraction (hereafter referred to as MWS) had Dectin-2 ligand activity
16
(Figure 5C). The MWS could activate cells expressing Dectin-2 in amounts as low as 50
ng (Figure 5C), whereas it had no effect on Mincle-expressing cells, even in amounts as
high as 5 g (data not shown). We have previously reported that the cell wall matrix glycoprotein of Malassezia contains cell wall -glucan (Shibata et al., 2009). To enrich
the Dectin-2 ligands, we digested -glucan in MWS with westase (-1,6-glucanase)
(Shibata et al., 2009), because it has not been reported that Dectin-2 recognize -glucan.
We confirmed that the westase digestion of MWS did not impair ligand activity (data
not shown).
The reaction product was fractionated by gel filtration chromatography to
give four fractions, W1-W4, separated on the basis of their molecular mass (Figure 5D).
W1 activated reporter cells expressing Dectin-2, whereas W2, W3 and W4 did not show
strong Dectin-2 ligand activity (Figure 5E).
Identification of O-linked manno-protein as a Dectin-2 ligand
Dectin-2 recognizes several fungi that possess N-linked mannan on their
surface (McGreal et al., 2006; Sato et al., 2006). However, the 1H NMR analysis of W1
17
showed only two signals at the H-1 region (Figure 5F and Table 1), suggesting that W1
may possess structure distinct from N-linked mannan. The chemical shifts at 5.04 ppm
of W1 indicated the presence of a non-reducing terminal -1,2-linked mannose residue
(Shibata et al., 2007).
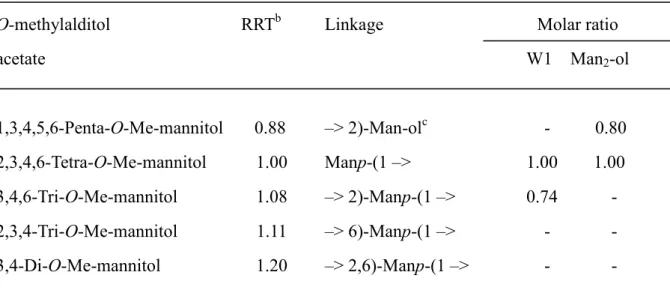
We further analyzed the structure of W1 by methylation analysis. W1 has only
a non-reducing terminal mannose residue and a 2-O-substituted mannose residue in the
molar ratio of 1:1 (Table S1). These data suggest that W1 may be a manno-protein
possessing predominantly O-linked manno-oligosaccharides, though such a cell wall
matrix glycoprotein has not been identified in yeasts and fungi.
To test this idea, we treated W1 with 0.1 M NaOH to induce -elimination,
which selectively releases O-linked oligosaccharides connected to serine and/or
threonine (Ser/Thr) residues. Bio-Gel P-2 column chromatography of the reaction
product showed that about 90% of the carbohydrate was released and eluted only in the
disaccharide fraction (Figure 5G). Sugar composition analysis revealed that the eluted
disaccharide consists of only mannose (Figure S7A). This result is significantly
different from that of the -elimination of the mannans from C. albicans, the amount of
18
the released oligosaccharides from which comprise only about 3-5% of the total
carbohydrate (data not shown).
The 1H NMR and methylation analyses of the released biose fraction
indicated that the O-linked oligosaccharide was an -1,2-linked mannobiose,
Man1-2Man (Table 1, Figures S7B, S7C and Table S1). To study the molar ratio of
Ser/Thr residues, we investigated the amino acid composition of the protein moiety of
mannosyl W1, and found that W1 was abundant in serine (32.9%, Table S2).
On the basis of these results, we concluded that the structure of W1 was a
mucin-like serine-rich glycoprotein and the O-linked oligosaccharide was
predominantly -1,2-linked mannobiose (Figure 5H). Thus, although Dectin-2
recognizes Malassezia as well as C. albicans, the ligand structure of Malassezia was
quite different from that of C. albicans.
Dectin-2 is essential for the cytokine production induced by W1
Next, to assess whether Dectin-2 is an essential receptor for the Malassezia
ligand in DCs, WT, Mincle–/–,Dectin-2–/– and FcR–/– BMDCs were stimulated with
19
MWS and W1. MWS and W1 were capable to activate WT DCs to produce TNF.
However, the TNF production was almost completely suppressed in Dectin-2–/– and
FcR–/– cells, but not in Mincle–/– DCs (Figure 6A). These findings demonstrated that
Dectin-2 is an essential receptor for the Malassezia-derived O-mannobiose-rich protein
which can directly activate DCs to produce inflammatory cytokines.
Malassezia-derived ligands induces host immune responses
We then examined the effect of these Malassezia-derived ligands on immune
responses. First, 44-2 and W1 were intraperitoneally injected in mice to assess innate
immune responses against these ligands. 44-2 and W1 are capable of inducing
neutrophil infiltration into peritoneal cavities (Figure 6B). Furthermore, whole
Malassezia cells and 44-2 showed adjuvanticity toward acquired immune responses,
such as IFN production, in response to recall antigen stimulation. Although Mincle
ligand is capable of driving Th17 responses (Werninghaus et al., 2009), IL-17 was not
detected in our limited condition (Figure S6).
20
Mincle and Dectin-2 contribute to cytokine production in response to whole
Malassezia cells
Finally, we investigated the contribution of the two CLRs to the recognition of
whole Malassezia cells. The production of TNF in response to M. furfur was decreased
in the absence of Mincle or Dectin-2, thus suggesting that both Mincle and Dectin-2 can
mediate DC activation in response to M. furfur (Figure 6C). In line with these
observations, the TNF production was severely impaired in DCs lacking FcR, a
common subunit of Mincle and Dectin-2. We found that M. furfur also induces IL-10
production, which was also dependent on Mincle and Dectin-2 (Figure 6D). These
results suggest that Mincle and Dectin-2 cooperatively contribute to cytokine
production in response to the Malassezia species.
21 DISCUSSION
Here we have shown that the identification of distinct ligands for Mincle and
Dectin-2 in Malassezia fungus.
The general principle of the Mincle ligand structure has not yet been clearly
defined. Mincle recognizes TDM and its analogue trehalose dibehenate (TDB), but not
mycolate or trehalose alone (Ishikawa et al., 2009; Schoenen et al., 2010). Trehalose is a
disaccharide formed by two glucose with an ,-1,1 linkage. In this study, we found
that Mincle ligand 44-1 has one gentiobiose, a disaccharide composed of two units of
glucose with a -1,6 linkage, and two fatty acids, which are attached via ester bonds to
the hydroxyl groups of the glycerol backbone. In contrast, 44-2 has two mannosylated
fatty acids, and one -1,2-linked mannobiosylated fatty acid, which are attached to the
mannitol backbone, although there was minor structural variability such as number of
mannose or hydroxyl fatty acid residues. A bipolar glycolipid with disaccharide
composed of glucose or mannose attached to fatty acids may represent a potential
minimal ligand structure for Mincle. Furthermore, comparisons of the fatty acids
moieties of 44-1, 44-2, TDM and TDB suggest that the length of the fatty acids may not
22 critically influence the Mincle ligand activity.
The reason why Mincle selectively recognizes Malassezia among the various
fungi remains unclear. Given that Malassezia uniquely requires lipid for their growth
(Schmidt, 1997), the uptake of exogenous lipid as nutrition may be required for the
biosynthesis of long-chain fatty acid moiety of Mincle ligands in fungus. Currently, the
roles of 44-1 and 44-2 in the physiology and pathogenesis of Malassezia still remain
unclear, although it is possible that other glycolipids are synthesized in other Malassezia
strains.
On the other hand, it has been reported that Dectin-2 recognizes the terminal
mannose of N-linked glycan (McGreal et al., 2006). In the present study, we
demonstrated that O-linked manno-protein could be a Dectin-2 ligand. -1,2-mannosyl
residues of W1 were necessary and sufficient for the recognition by Dectin-2 (Figures
S7D and S7E). Although we detected similar O-linked manno-protein in M.
pachydermatis and M. sympodialis (Figure S1B and S1C), other Malassezia strains may
contain different forms of Dectin-2 ligands. Taken together, active Dectin-2 ligand
could be defined as a high density of terminal -1,2-mannose attached to glycans,
23
proteins, and presumably any kind of scaffold. In line with this hypothesis, C.
guillermondii and S. cerevisiae were not recognized by Dectin-2 (Figure 1), most likely
because the cell wall of these fungi contains -1,2-mannose masked with
-1,2-mannose and -1,3-mannose, respectively (Romero et al., 1999; Shibata et al.,
1996).
Protein mannosylation is an important process in fungal physiology. It has
been reported that protein O-mannosyl transferases (PMTs) and -1,2-mannosyl
transferases (MNTs) mediate O-mannosylation of proteins in several fungi (Deshpande
et al., 2008). Deficiency of these enzymes in several fungi results in an attenuation in
their virulence, adherence to host cells, biofilm formation, and cell interaction during
mating (Munro et al., 2005; Timpel et al., 1998). Therefore, the O-mannosylated
products would be one of the appropriate pathogen-associated molecular patterns
(PAMPs) for the host to induce an immune response against the fungus.
Characterization of W1 core protein by SDS-PAGE suggested that apparent molecular
mass of protein moiety is approximately 10 kDa (Figure S5). The identification of
mannosyl transferase in Malassezia may help to clarify the mechanism regulating the
24 biosynthesis of such PAMPs.
Several reports have suggested that Malassezia species are associated with
atopic dermatitis (Ashbee and Evans, 2002; Scheynius et al., 2002). Many kinds of
antigens in Malassezia have been demonstrated to react with patient IgE (Ashbee and
Evans, 2002). However, the precise molecular mechanisms underlying the pathogenesis
remain unclear. The reactivity of the O-mannosyl protein to patient IgE would be an
intriguing issue to be addressed.
Malassezia is known to alter the antigens expressed throughout their growth
cycle and culture conditions (Ashbee and Evans, 2002; Shibata et al., 2009). Indeed, It
is therefore possible that the relative amount, localization and structure of
Mincle/Dectin-2 ligands in Malassezia may also vary according to the life cycle,
nutritional status, temperature or substrains. Taken together, the acquisition of two
CLRs recognizing different ligands in the same fungus would enable the host to exert
stable immune responses against variable pathogens.
A synthetic LTA anchor, which has structural similarity to 44-1, has been
reported to induce TNF production from macrophages in a TLR-independent manner
25
(Morath et al., 2002). Mincle might be a responsible receptor for this response.
Although Malassezia is a pathogenic fungus, it is usually a harmless
commensal found in healthy skin. It is important to examine whether the
expression/function of Mincle/Dectin-2 in langerhans cells or dermal DCs is
downregulated in healthy skin. Alternatively, some inhibitory receptors recognizing
Malassezia may be expressed in such dermal cells to prevent unnecessary DC activation.
IL-10, an anti-inflammatory cytokine induced by Malassezia, may also play a role in
regulating host immune responses to Malassezia.
It has recently been revealed that several CLRs recognize the
damage-associated molecular patterns (DAMPs) derived from damaged tissue (Aragane
et al., 2003; Nauta et al., 2003; Ogden et al., 2001; Oka et al., 1998; Yamasaki et al.,
2008; Yuita et al., 2005). On the other hand, terminal mannose residues of glycoproteins
become exposed upon inflammation and stresses (Franz et al., 2006), although they are
normally masked with complex branched sugars during protein maturation in
vertebrates (Green et al., 2007). It is therefore possible that Dectin-2 may recognize
damaged-self through terminal mannose residues of self protein. The identification of
26
endogenous ligand for Mincle, Dectin-2 and other CLRs may help to elucidate the
immune responses to damaged-tissue through DAMPs-PRRs interaction.
27 EXPERIMENTAL PROCEDURES
Mice. Dectin-2-deficient mice and FcR-deficient mice on the C57BL/6 background
were described previously (Park et al., 1998; Saijo et al., 2010). Mincle-deficient mice,
described previously (Yamasaki et al., 2009), were backcrossed for at least nine
generations with C57BL/6 mice. All mice were maintained in a filtered-air laminar-flow
enclosure and given standard laboratory food and water ad libitum. All animal protocols
were approved by the committee of Ethics on Animal Experiment, Faculty of Medical
Sciences, Kyushu University.
Fungi. M. pachydermatis (IFM No. 48586) was grown on agar plates (Wako) or liquid
medium with potato dextrose broth (Difco Laboratories) for 5 days at 32°C. M. furfur
(IFM No. 52635) was grown in potato dextrose agar supplemented with 100 l olive oil
(Figure 1), or grown in potato dextrose liquid medium supplemented with 1%
Tween-80 (Nacalai tesque) (Figures 6C and 6D) for 5 days at 32°C.
28
Reagents. TDM, D-glucose, D-mannose and D-galactose were purchased from Nacalai
tesque. Zymosan (Z4250), LTA (L4015), OVA (A5503) and -mannosidase (M7257)
were purchased from Sigma-Aldrich. Westase (9095) and Candida albicans cell wall
mannan (MG001) were obtained from Takara Bio. Other reagents used for chemical
analyses were described in Supplemental Information.
In vitro stimulation. To stimulate the cells, TDM was dissolved in C:M (2:1) at 1
mg/ml in isopropanol. Then, these extracts from fungi were added to 96-well plates at
20 l/well, followed by evaporation of the solvent, as described previously (Ishikawa et
al., 2009).
In vivo stimulation. For innate immune responses, mice were intraperitoneally injected
with 44-2 or W1 ligands in oil-in-water consisting of mineral oil (9%), Tween-80 (1%)
and PBS (90%), or in PBS alone, respectively. At 20 h after injection, peritoneal cells
were collected and stained with anti-CD11b and anti-Gr1 mAb and analyzed by flow
cytometry. For acquired immune responses, mice were immunized with 4 x 107 M.
29
furfur (i.p.) or 200 g ovalbumin (OVA) together with 200 g Malassezia-derived
ligands (s.c.). At 7 days after immunization, splenocytes or inguinal lymph node cells
were collected and cultured at 3 x 105 cells/200 l with Malassezia antigen or OVA for
72 h. Cytokine concentrations in the culture supernatants were determined by ELISA.
Cells. 2B4-NFAT-GFP reporter cells expressing Mincle, Dectin-2 and Dectin-2QPD
mutant (E168Q/N170D) were prepared as previously described (Yamasaki et al., 2008).
For BMDC preparation, BM cells were suspended in RPMI 1640 medium
supplemented with 10% (vol/vol) FCS and -mercaptethanol, and were plated at a
density of 5 ×106 cells/ml in the presence of culture supernatant of MGM-5 (provided
by Dr. S. Nagata) as a sourse of GM-CSF, and were cultured for 6 days at 37°C. For
BMDMs, L929-conditioned medium were used as a source of M-CSF, and adherent
cells were used for the in vitro experiments. ELISA kit for TNF, IL-10, IL-4, IFN and
IL-17 were purchased from BD Biosciences or R&D Systems.
30
Preparation of Malassezia lipophilic fraction. M. pachydermatis was treated with
C:M (2:1; vol/vol), hexane, acetone, 1-butanol (BuOH), or distilled water. The insoluble
fractions were collected. The soluble fractions were further partitioned by C:M:W
(8:4:3; vol/vol) into a lower organic phase (C:M) and upper aqueous phase (M:W). The
upper aqueous phase (M:W) was further partitioned by 1-butanol:water (1:1; vol/vol)
into an upper butanol phase (BuOH) and a lower aqueous phase (water). Each fraction
was resuspended in a volume of 2-propanol relative to the original cell pellet weight,
and tested as lipid extracts (Morita et al., 2005).
Preparation of Malassezia water-soluble fraction (MWS). M. furfur cells were
washed with deionized water and dehydrated with acetone. The crude cell surface
matrix glycoproteins were extracted with deionized water at 120°C for 2 h. After
centrifugation, the soluble extract was dialyzed against running tap water for 2 days,
then were evaporated and lyophilized and used as Malassezia water-soluble fraction
(MWS).
31
Preparation of W1. MWS was dissolved in 25 ml of 100 mM McIlvain buffer (pH 6.0)
and 50 units of westase were added and incubated at 37°C for 24 h. The enzyme was
inactivated by heating at 100°C for 5 min, and the supernatant was separated by
centrifugation at 3,000 rpm for 10 min, followed by evaporation. The westase reaction
product was applied onto a column (4.0 x 40 cm) of Sephacryl S-100 and eluted with
deionized water to yield four fractions, W1, W2, W3, and W4. The amount of W1 was
approximately 0.5% of the dried cell mass.
Chemical analysis. Fast atom bombardment mass spectrometry (FAB-MS),
electrospray ionization-time of flight mass spectrometry (ESI-TOFMS), gas
chromatography-mass spectrometry (GC-MS) and Nuclear magnetic resonance
spectroscopy (1H NMR and 13C NMR) were performed as described in Supplemental
Information.
Release of O-linked oligosaccharides from W1 by -elimination. W1 (20 mg) was
dissolved in 0.5 M NaBH4/0.1 M NaOH and incubated at 25°C for 18 h. The reaction
32
mixture was neutralized with acetic acid and repeatedly evaporated with methanol to
remove boric acid. The reaction mixture was dissolved in 1 ml of water and was applied
onto a column (2.5 x 100 cm) of Bio-Gel P-2 and eluted with water. The released
oligosaccharide was analyzed by 1H NMR and the methylation analyses.
Carbohydrate composition analysis. For the analysis of the carbohydrate composition,
samples were hydrolyzed with 4 M trifluoroacetic acid (TFA) at 100°C for 3 h. The
resulting monosaccharide mixtures were reduced by treatment with NaBH4 and
acetylated by acetic anhydride/pyridine (1:1, v/v). The reagents were evaporated and
analyzed by GC/MS.
Monosaccharide linkage analysis. The methylation analysis was carried out according
to the method reported by Ciucanu and Kerek (Ciucanu and Kerek, 1984). The
manno-protein or oligosaccharide was dissolved in a NaOH/dimethylsulfoxide
suspension prepared using powdered NaOH. After stirring for 30 min, methyl iodide
was added, and the suspension was stirred for another 30 min. The methylated product
33
was extracted into chloroform and washed with water. The permethylated carbohydrates
were then hydrolyzed in 2 M trifluoroacetic acid at 110°C for 2 h. The partially
methylated monosaccharides were reduced with 1% NaBD4 at room temperature for 18
h. Following borate removal by drying from methanol, the partially methylated alditols
were acetylated by adding acetic anhydride/pyridine (1:1, v/v) and incubating them at
50°C for 3 h. The reagents were evaporated and analyzed by GC/MS.
34 ACKNOWLEDGEMENTS
We thank Y. Iwakura and S. Akira for providing mutant mice and Y. Nishi-Sanui for
secretary assistance. This work was supported by Grant-in-Aid for Young Scientists (S),
Funding Program for Next Generation World-Leading Researchers (NEXT Program),
Ono Medical Research Foundation (SY), and Grant-in-Aid for JSPS (TI). This work
was partly performed in the Cooperative Research Project Program of the Medical
Institute of Bioregulation, Kyushu University.
35 REFERENCES
Aragane, Y., Maeda, A., Schwarz, A., Tezuka, T., Ariizumi, K., and Schwarz, T. (2003).
Involvement of dectin-2 in ultraviolet radiation-induced tolerance. J Immunol. 171,
3801-3807.
Ashbee, H.R., and Evans, E.G. (2002). Immunology of diseases associated with
Malassezia species. Clin Microbiol Rev. 15, 21-57.
Ciucanu, I., and Kerek, F. (1984). A simple and rapid method for the permethylation of
carbohydrates. Carbohydrate Research. 131, 209-217.
Deshpande, N., Wilkins, M.R., Packer, N., and Nevalainen, H. (2008). Protein
glycosylation pathways in filamentous fungi. Glycobiology. 18, 626-637.
Drickamer, K. (1992). Engineering galactose-binding activity into a C-type
mannose-binding protein. Nature. 360, 183-186.
Franz, S., Frey, B., Sheriff, A., Gaipl, U.S., Beer, A., Voll, R.E., Kalden, J.R., and
Herrmann, M. (2006). Lectins detect changes of the glycosylation status of plasma
membrane constituents during late apoptosis. Cytometry A. 69, 230-239.
Gaitanis, G., Magiatis, P., Hantschke, M., Bassukas, I.D., and Velegraki, A. (2012). The
36
Malassezia genus in skin and systemic diseases. Clin Microbiol Rev. 25, 106-141.
Green, R.S., Stone, E.L., Tenno, M., Lehtonen, E., Farquhar, M.G., and Marth, J.D.
(2007). Mammalian N-glycan branching protects against innate immune
self-recognition and inflammation in autoimmune disease pathogenesis. Immunity. 27,
308-320.
Guillot, J., and Bond, R. (1999). Malassezia pachydermatis: a review. Med Mycol. 37,
295-306.
Hardison, S.E., and Brown, G.D. (2012). C-type lectin receptors orchestrate antifungal
immunity. Nat Immunol. 13, 817-822.
Hunter, R.L., Olsen, M.R., Jagannath, C., and Actor, J.K. (2006). Multiple roles of cord
factor in the pathogenesis of primary, secondary, and cavitary tuberculosis, including a
revised description of the pathology of secondary disease. Ann Clin Lab Sci. 36,
371-386.
Ishikawa, E., Ishikawa, T., Morita, Y.S., Toyonaga, K., Yamada, H., Takeuchi, O.,
Kinoshita, T., Akira, S., Yoshikai, Y., and Yamasaki, S. (2009). Direct recognition of the
mycobacterial glycolipid, trehalose dimycolate, by C-type lectin Mincle. J Exp Med.
37 206, 2879-2888.
Marcon, M.J., and Powell, D.A. (1992). Human infections due to Malassezia spp. Clin
Microbiol Rev. 5, 101-119.
Matsumoto, M., Tanaka, T., Kaisho, T., Sanjo, H., Copeland, N.G., Gilbert, D.J., Jenkins,
N.A., and Akira, S. (1999). A novel LPS-inducible C-type lectin is a transcriptional
target of NF-IL6 in macrophages. J Immunol. 163, 5039-5048.
McGreal, E.P., Rosas, M., Brown, G.D., Zamze, S., Wong, S.Y., Gordon, S.,
Martinez-Pomares, L., and Taylor, P.R. (2006). The carbohydrate-recognition domain of
Dectin-2 is a C-type lectin with specificity for high mannose. Glycobiology. 16,
422-430.
Morath, S., Stadelmaier, A., Geyer, A., Schmidt, R.R., and Hartung, T. (2002). Synthetic
lipoteichoic acid from Staphylococcus aureus is a potent stimulus of cytokine release. J
Exp Med. 195, 1635-1640.
Morita, Y.S., Velasquez, R., Taig, E., Waller, R.F., Patterson, J.H., Tull, D., Williams,
S.J., Billman-Jacobe, H., and McConville, M.J. (2005). Compartmentalization of lipid
biosynthesis in mycobacteria. J Biol Chem. 280, 21645-21652.
38
Munro, C.A., Bates, S., Buurman, E.T., Hughes, H.B., Maccallum, D.M., Bertram, G.,
Atrih, A., Ferguson, M.A., Bain, J.M., Brand, A., et al. (2005). Mnt1p and Mnt2p of
Candida albicans are partially redundant alpha-1,2-mannosyltransferases that participate
in O-linked mannosylation and are required for adhesion and virulence. J Biol Chem.
280, 1051-1060.
Nauta, A.J., Raaschou-Jensen, N., Roos, A., Daha, M.R., Madsen, H.O., Borrias-Essers,
M.C., Ryder, L.P., Koch, C., and Garred, P. (2003). Mannose-binding lectin engagement
with late apoptotic and necrotic cells. Eur J Immunol. 33, 2853-2863.
Ogden, C.A., deCathelineau, A., Hoffmann, P.R., Bratton, D., Ghebrehiwet, B., Fadok,
V.A., and Henson, P.M. (2001). C1q and mannose binding lectin engagement of cell
surface calreticulin and CD91 initiates macropinocytosis and uptake of apoptotic cells. J
Exp Med. 194, 781-795.
Oka, K., Sawamura, T., Kikuta, K., Itokawa, S., Kume, N., Kita, T., and Masaki, T.
(1998). Lectin-like oxidized low-density lipoprotein receptor 1 mediates phagocytosis
of aged/apoptotic cells in endothelial cells. Proc Natl Acad Sci U S A. 95, 9535-9540.
Pan, K., Inagaki, M., Ohno, N., Tanaka, C., Higuchi, R., and Miyamoto, T. (2010).
39
Identification of sixteen new galactocerebrosides from the starfish Protoreaster nodosus.
Chem Pharm Bull (Tokyo). 58, 470-474.
Park, S.Y., Ueda, S., Ohno, H., Hamano, Y., Tanaka, M., Shiratori, T., Yamazaki, T.,
Arase, H., Arase, N., Karasawa, A., et al. (1998). Resistance of Fc receptor- deficient
mice to fatal glomerulonephritis. J Clin Invest. 102, 1229-1238.
Redline, R.W., and Dahms, B.B. (1981). Malassezia pulmonary vasculitis in an infant
on long-term Intralipid therapy. N Engl J Med. 305, 1395-1398.
Ritter, M., Gross, O., Kays, S., Ruland, J., Nimmerjahn, F., Saijo, S., Tschopp, J.,
Layland, L.E., and Prazeres da Costa, C. (2010). Schistosoma mansoni triggers Dectin-2,
which activates the Nlrp3 inflammasome and alters adaptive immune responses. Proc
Natl Acad Sci U S A. 107, 20459-20464.
Robinson, M.J., Sancho, D., Slack, E.C., LeibundGut-Landmann, S., and Reis e Sousa,
C. (2006). Myeloid C-type lectins in innate immunity. Nat Immunol. 7, 1258-1265.
Romero, P.A., Lussier, M., Veronneau, S., Sdicu, A.M., Herscovics, A., and Bussey, H.
(1999). Mnt2p and Mnt3p of Saccharomyces cerevisiae are members of the Mnn1p
family of alpha-1,3-mannosyltransferases responsible for adding the terminal mannose
40
residues of O-linked oligosaccharides. Glycobiology. 9, 1045-1051.
Saijo, S., Ikeda, S., Yamabe, K., Kakuta, S., Ishigame, H., Akitsu, A., Fujikado, N.,
Kusaka, T., Kubo, S., Chung, S.H., et al. (2010). Dectin-2 recognition of alpha-mannans
and induction of Th17 cell differentiation is essential for host defense against Candida
albicans. Immunity. 32, 681-691.
Sato, K., Yang, X.L., Yudate, T., Chung, J.S., Wu, J., Luby-Phelps, K., Kimberly, R.P.,
Underhill, D., Cruz, P.D., Jr., and Ariizumi, K. (2006). Dectin-2 is a pattern recognition
receptor for fungi that couples with the Fc receptor gamma chain to induce innate
immune responses. J Biol Chem. 281, 38854-38866.
Scheynius, A., Johansson, C., Buentke, E., Zargari, A., and Linder, M.T. (2002). Atopic
eczema/dermatitis syndrome and Malassezia. Int Arch Allergy Immunol. 127, 161-169.
Schmidt, A. (1997). Malassezia furfur: a fungus belonging to the physiological skin
flora and its relevance in skin disorders. Cutis. 59, 21-24.
Schoenen, H., Bodendorfer, B., Hitchens, K., Manzanero, S., Werninghaus, K.,
Nimmerjahn, F., Agger, E.M., Stenger, S., Andersen, P., Ruland, J., et al. (2010). Cutting
edge: Mincle is essential for recognition and adjuvanticity of the mycobacterial cord
41
factor and its synthetic analog trehalose-dibehenate. J Immunol. 184, 2756-2760.
Schwandner, R., Dziarski, R., Wesche, H., Rothe, M., and Kirschning, C.J. (1999).
Peptidoglycan- and lipoteichoic acid-induced cell activation is mediated by toll-like
receptor 2. J Biol Chem. 274, 17406-17409.
Shibata, N., Akagi, R., Hosoya, T., Kawahara, K., Suzuki, A., Ikuta, K., Kobayashi, H.,
Hisamichi, K., Okawa, Y., and Suzuki, S. (1996). Existence of novel branched side
chains containing beta-1,2 and alpha-1,6 linkages corresponding to antigenic factor 9 in
the mannan of Candida guilliermondii. J Biol Chem. 271, 9259-9266.
Shibata, N., Saitoh, T., Tadokoro, Y., and Okawa, Y. (2009). The cell wall
galactomannan antigen from Malassezia furfur and Malassezia pachydermatis contains
beta-1,6-linked linear galactofuranosyl residues and its detection has diagnostic
potential. Microbiology. 155, 3420-3429.
Shibata, N., Suzuki, A., Kobayashi, H., and Okawa, Y. (2007). Chemical structure of the
cell-wall mannan of Candida albicans serotype A and its difference in yeast and hyphal
forms. Biochem J. 404, 365-372.
Takeuchi, O., and Akira, S. (2010). Pattern recognition receptors and inflammation. Cell.
42 140, 805-820.
Taylor, P.R., Reid, D.M., Heinsbroek, S.E., Brown, G.D., Gordon, S., and Wong, S.Y.
(2005). Dectin-2 is predominantly myeloid restricted and exhibits unique
activation-dependent expression on maturing inflammatory monocytes elicited in vivo.
Eur J Immunol. 35, 2163-2174.
Timpel, C., Strahl-Bolsinger, S., Ziegelbauer, K., and Ernst, J.F. (1998). Multiple
functions of Pmt1p-mediated protein O-mannosylation in the fungal pathogen Candida
albicans. J Biol Chem. 273, 20837-20846.
Werninghaus, K., Babiak, A., Gross, O., Holscher, C., Dietrich, H., Agger, E.M., Mages,
J., Mocsai, A., Schoenen, H., Finger, K., et al. (2009). Adjuvanticity of a synthetic cord
factor analogue for subunit Mycobacterium tuberculosis vaccination requires
FcRgamma-Syk-Card9-dependent innate immune activation. J Exp Med. 206, 89-97.
Yamasaki, S., Ishikawa, E., Sakuma, M., Hara, H., Ogata, K., and Saito, T. (2008).
Mincle is an ITAM-coupled activating receptor that senses damaged cells. Nat Immunol.
9, 1179-1188.
Yamasaki, S., Matsumoto, M., Takeuchi, O., Matsuzawa, T., Ishikawa, E., Sakuma, M.,
43
Tateno, H., Uno, J., Hirabayashi, J., Mikami, Y., et al. (2009). C-type lectin Mincle is an
activating receptor for pathogenic fungus, Malassezia. Proc Natl Acad Sci U S A. 106,
1897-1902.
Yuita, H., Tsuiji, M., Tajika, Y., Matsumoto, Y., Hirano, K., Suzuki, N., and Irimura, T.
(2005). Retardation of removal of radiation-induced apoptotic cells in developing neural
tubes in macrophage galactose-type C-type lectin-1-deficient mouse embryos.
Glycobiology. 15, 1368-1375.
44 FIGURE LEGENDS
Figure 1. Dectin-2 recognizes Malassezia species.
Screening of pathogenic fungi for Dectin-2 ligand activities. The reporter cell lines
expressing Dectin-2 + FcR were co-cultured with the indicated pathogenic fungi for 18
h. The NFAT-GFP induction was analyzed by flow cytometry (See also Figure S1).
Representative results from two independent experiments with similar results are
shown.
Figure 2. Isolation of Mincle and Dectin-2 ligands in Malassezia.
(A) A schematic diagram of the solvent-based fractionation of M. pachydermatis.
Solvent-treated fungal cells (ppt, gray boxes) and soluble extracts (sup, open boxes)
were subjected to reporter assays. C:M, chroloform:methanol.
(B-E) Solvent-treated fungal cells (B and D) and plated-coated soluble extracts (C and
E) of live M. pachydermatis were co-cultured with reporter cells expressing Mincle +
FcR (B-C) or Dectin-2 + FcR (D-E). The NFAT-GFP induction was analyzed by flow
cytometry (See also Figure S2).
45
All data (B-E) are the means ± SD for triplicate assays, and representative results from
three independent experiments with similar results are shown.
Figure 3. Identification of Mincle ligands in Malassezia.
(A) Silica gel column purification. C:M extract of M. pachydermatis was subjected to
silica gel column chromatography (Silica gel 60, Merck) and eluted with
CHCl3/MeOH/H2O (9/1/0.1 to 1/1/0, v/v/v) to give 49 fractions. Each fraction was
coated onto a plate to stimulate reporter cells expressing Mincle and FcR (top). Each
fraction was also separated by TLC followed by orcinol staining (bottom). The
arrowhead indicates Fraction #44.
(B) TLC separation. Fraction #44 (Fr. 44) was analyzed by HPTLC and divided into 20
subfractions. Each subfraction was coated onto a plate to stimulate reporter cells
expressing Mincle and FcR. The black arrowheads show the origin and solvent fronts.
The red arrowheads indicate spots corresponding to 44-1 and 44-2 by orcinol staining.
Purified TDM was used as a control.
46
(C) Mass spectrum of 44-1. The negative FAB-MS spectrum of 44-1 showed a
pseudo-molecular ion peaks [M-H]– at m/z: 919. The fragment ion peaks due to fatty
acid anions were observed at m/z: 297 [C19H37COO–] and 241 [C14H29COO–].
(D) The chemical structure of Mincle ligand 44-1. (See also Figure S3)
(E) Mass spectrum of 44-2. The molecular formula of 44-2 was determined to be
C84H156NaO32 by the ESI-TOFMS [m/z 1700.0501, calcd. 1700.0472 (M+Na)+].
(F) The chemical structure of Mincle ligand 44-2. (See also Figure S3)
Figure 4. Mincle is necessary and sufficient for the recognition of two glycolipids
from Malassezia.
(A) 44-1 and 44-2 derived from M. pachydermatis activated Mincle-expressing cells.
NFAT-GFP reporter cells expressing Mincle + FcR were co-cultured for 18 h with
plates coated with indicated amount of 44-1, 44-2 or TDM (0.27 µg/well) as a control
(See also Figure S4).
(B) Binding of Mincle-Ig fusion protein to 44-1 and 44-2. Serially diluted Ig control
(Ig) and Mincle-Ig fusion proteins were allowed to react with 44-1 and 44-2 coated on
47
ELISA plates (1 g/well). W1 and TDM were used as negative and positive control,
respectively. The bound Fc fusion proteins were detected using HRP-conjugated
anti-hIgG.
(C-D) Essential role of Mincle-FcR axis in cytokine production induced by 44-1 and
44-2. BMDCs from WT, FcR–/–, Dectin-2–/– and Mincle–/– mice were stimulated with
plates coated 44-1 or 44-2 (C) or zymosan (10 µg/ml) as a control (D). The culture
supernatants were collected at 48 h and their concentrations of TNF were determined by
ELISA.
The data (B-D) are the means ± SD for triplicate assays, and representative results from
two independent experiments with similar results are shown.
Figure 5. Identification of the Dectin-2 ligand in Malassezia.
(A) Role of mannose-binding motif of Dectin-2 in Malassezia recognition. NFAT-GFP
reporter cells expressing FcR together with Dectin-2WT or Dectin-2QPD were
co-cultured for 18 h with M. furfur.
48
(B) Blocking of Malassezia recognition by monosaccharides. NFAT-GFP reporter cells
expressing Dectin-2 + FcR were co-cultured with 3 106 of M. furfur in the presence
of glucose, mannose or galactose.
(C) Activation of Dectin-2-expressing cells by hydrophilic fraction of M. furfur.
NFAT-GFP reporter cells expressing Dectin-2 + FcR were co-cultured for 18 h with
plates coated MWS (0.05 – 5 µg/well) or Candida albicans cell wall mannan (C. a.
mannan; 0.1 µg/well) as a control.
(D) Size fractionation by gel filtration chromatography. Separation of MWS was
performed with a column (4.0 x 40 cm) of Sephacryl S-100, and the carbohydrate
content in each fraction was assayed by the phenol/sulfuric acid method. Four fractions,
W1-W4, were collected on the basis of their molecular mass and subjected to the
following assay.
(E) Detection of ligand activity in W1 fraction. NFAT-GFP reporter cells expressing
Dectin-2 or Mincle were co-cultured for 18 h with plates coated W1, W2, W3 and W4
(0.01 – 1 µg/well) or TDM (0.27 µg/ml/well) as a control.
(F) The 1H NMR analysis of M. furfur W1. W1 was dissolved in D2O. The 1H NMR
49
spectra were recorded by a JNM-LA600 spectrometer (JEOL) at 45C. (See also Table
S2)
(G) Gel filtration chromatography. The elution profiles of -elimination products of M.
furfur W1 were shown. Elution was performed with a column (2.5 x 100 cm) of
Bio-Gel P-2, and the carbohydrate content in each fraction was assayed by the
phenol/sulfuric acid method. The arrows indicate the fraction corresponding to
monosaccharide (mono), disaccharide (di), trisaccharide (tri) and polysaccharide (poly).
(See also Figure S5)
(H) Schematic representation of a possible structure of W1. X is any amino acid.
The data (A, B, C and E) are the means ± SD for triplicate assays, and representative
results from three independent experiments with similar results are shown.
Figure 6. Mincle and Dectin-2 mediate Malassezia-induced immune responses.
(A) BMDCs from WT, FcR–/–, Dectin-2–/– and Mincle–/– mice were stimulated with
plates coated W1 or MWS or zymosan (10 µg/ml) as a control. Culture supernatants
were collected at 48 h, and their concentrations of TNF were determined by ELISA.
50
(B) Mice were intraperitoneally injected with 200 g 44-2 in oil-in-water emulsion
consisting of mineral oil (9%), Tween-80 (1%) and PBS (90%). Mice were also injected
i.p. with 200 g W1 in PBS. At 20 h after injection, peritoneal cells were stained with
CD11b and Gr1 and analyzed by flow cytometry. Each symbol represents an individual
mouse. (See also Figure S6) *, P < 0.05.
(C-D) BMDCs from WT, FcR–/–, Dectin-2–/– and Mincle–/– mice were stimulated with
M. furfur or zymosan (10 µg/ml) as a control. After 48 h culture, the concentrations of
TNF (C) and IL-10 (D) were determined by ELISA.
The data (A) are the means ± SD for triplicate assays, and representative results from
three independent experiments with similar results are shown. The data (C-D) are the
means ± SD for triplicate assays, and representative results from two independent
experiments with similar results are shown.
Figure 1 !
Figure 2 !
A !
B ! C !
E !
D !
Figure 3 !
A ! B !
C ! D !
E ! F !
44-1!
44-2!
Figure 4 !
A ! B !
C ! D !
Figure 5 !
A ! B !
D !
C !
E!
F ! G !
H !
Figure 6 !
A !
D !
B ! C !
Table 1. 1H and 13C chemical shifts (!, ppm)a for W1b and mannobiose released from W1 by !-elimination.
Mannoprotein or
Oligosaccharide Residue H-1 H-2 H-3 H-4 H-5 H-6/H-6’
(JH1, H2) (JH2, H3) (JH3, H4)
mannoprotein
W1 !2Man"-O-Ser/Thr 5.123 3.960 3.783 3.636 -d - Man"1! 5.037 4.073 3.843 3.652 - -
!-elimination
Man2-ol !2Man-ole 3.926 3.783 4.021 3.690 3.768 3.877/3.679 Man"1! 5.001 3.989 3.871 3.668 3.799 3.908/-
(1.8)c (3.3) (9.6)
Residue C-1 C-2 C-3 C-4 C-5 C6
Man2-ol !2Man-ol 62.07 80.27 68.49 70.40 71.91 63.98 Man"1! 102.00 71.32 71.20 67.68 74.06 61.81
a Sample was dissolved in D2O. The 1H NMR spectra were recorded by a JNM-LA600 spectrometer (JEOL) at 45°C. The proton and carbon chemical shifts were referenced relative to the internal acetone at # 2.225 and 31.07, respectively.
b W1 was isolated from M. furfur.
c J = Hz.
d Not determined.
e Man-ol; mannitol
Supplemental Information
Identification of distinct ligands for Mincle and Dectin-2 in pathogenic fungus Malassezia
Tetsuaki Ishikawa, Fumie Itoh, Sayumi Yoshida, Shinobu Saijo, Tetsuhiro Matsuzawa, Tohru Gonoi, Takashi Saito, Yoshio Okawa, Nobuyuki Shibata,
Tomofumi Miyamoto and Sho Yamasaki
Figure S1 Recognition of Malassezia species by Mincle and Dectin-2 (related to Figure 1)
Figure S2 Isolation of 44-1 and 44-2 (related to Figure 2)
Figure S3 Structure determination of 44-1 and 44-2 (related to Figure 3) Figure S4 Structures and activities of 44-1- and 44-2-related compounds (related to Figure4)
Figure S5 Characterization of W1 (related to Figure 5)
Figure S6 Effect of 44-2 and W1 in acquired immunity (related to Figure 6) Figure S7 Carbohydrate analysis and activities of W1 (related to Table 1) Table S1 GC-MS analysis of W1 (related to Table 1)
Table S2 Amino acid composition of mannosyl-W1 (related to Figure 5)
Figure S1 A
Figure S1. Recognition of Malassezia species by Mincle. (related to Figure 1)
(A) Screening of pathogenic fungi for Mincle ligand activities. The reporter cell line expressing Mincle + FcR was co-cultured with the indicated pathogenic fungi for 18 h.
The NFAT-GFP induction was analyzed by flow cytometry. Representative results from two independent experiments with similar results are shown.
Figure S2
A
B
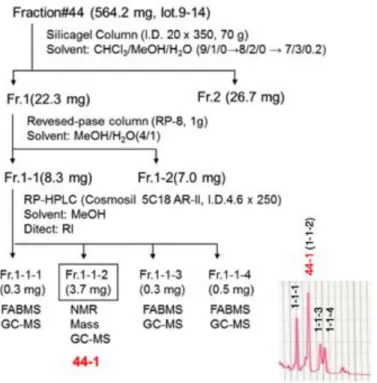
Figure S2. Isolation of 44-1 and 44-2. (related to Figure 2)
(A) Purification of 44-1. Fraction#44 (lot.9-14) was concentrated in vacuo to give a residue (564.2 mg), which was subjected to silica gel column chromatography (Silica gel 60, Merck, Darmstadt, Germany) eluted with CHCl3/MeOH/H2O (9/1/0 to 8/2/0 to 7/3/0.2, v/v/v) to give two fractions, Fraction 1 and 2 (11.7 mg). Fraction 1 (22.3 mg)
was further chromatographed on a reversed phase column chromatography (LiChroprep RP-8, Merck, Darmstadt, Germany), eluted with MeOH/H2O (4/1, v/v) to give two fractions, Fraction 1-1 and 1-2. Fraction 1-1 (8.3 mg) was further subjected to reversed phase HPLC (Cosmosil 5C18 AR-II, Nacalai Tesque, Tokyo, Japan), eluted with MeOH to give five fractions, Fraction 1-1-1 (0.3 mg), 1-1-2 (3.7 mg), 1-1-3 (0.3 mg), and 1-1-4 (0.5 mg). We named Fraction 1-1-2 as 44-1 (shown in red).
(B) Purification of 44-2. Fraction#44 (lot.20) was concentrated in vacuo to give a residue (71.0 mg), which was subjected to reversed-phase column chromatography (RP-8) eluted with MeOH/H2O (9/1/ to 9.5/1 to 1/0, v/v) to give three fractions.
Fraction 2 (7.4 mg) was further chromatographed on a silica gel column chromatography eluted with CHCl3/MeOH/H2O (7/3/0.2, v/v/v) to give three fractions.
We named Fraction 2-3 as 44-2 (shown in red).
A B
C D E
F G
H
I
J
K
L Figure S3
R
S
T
U
V W X
M N O
P
Q