Elucidation of the molecular mechanisms for
overwintering phenotype of Lotus japonicus
controlled by natural variation.
著者
Yusdar Mustamin
号
18
学位授与機関
Tohoku University
学位授与番号
生博第415号
氏 名 ( 本 籍 地 ) 学 位 の 種 類 学 位 記 番 号 学 位 授 与 年 月 日 学 位 授 与 の 要 件 研 究 科 , 専 攻 論 文 題 目 博士論文審査委員 ユスダル ムスタミン Yusdar Mustamin 博士(生命科学) 生博第 415 号 令和2年9月25日 学位規則第4条第1項該当 東北大学大学院生命科学研究科 (博士課程)生態システム生命科学専攻
Elucidation of the molecular mechanisms for
overwintering phenotype of Lotus japonicus controlled by natural variation. (自然変異によるミヤコグサ越冬性 の分子メカニズムの解明)
(主査)教 授 東谷 篤志 教 授 佐藤 修正 教 授 経塚 淳子
論文内容の要旨
As sessile organisms, plants must cope with their environmental conditions and thus have evolved a set of perception and signalling mechanisms to respond or adapt to adverse environmental conditions. Colonization of the new habitats is expected to require modification of these mechanisms to overcome challenges in new environmental factors. Temperature is one of the environmental factors that affect plant growth and development. Thus, adaptation to the low temperature is essential for long term evolutionary success of plants, especially for those in temperate region1. For perennial plants, the cold tolerance, in other words, winter-hardiness is fundamental mechanism for surviving during winter season.
In general, cold tolerance in many plant species is controlled by multiple genes and complex biological processes2. The process of gaining cold tolerance by low temperature stress that involved in both biochemical and physiological changes is known as cold acclimation. In the state of cold acclimation, stabilization of cell wall and membrane are induced by modification of metabolic processes, such as carbohydrate and lipid, through changing the regulation of sets of genes3,4,5. Among the gene expression regulation pathways involved in cold acclimation, ICE-CBF-COR pathway is the most characterized pathways mainly analyzed in Arabidopsis. Although the components of ICE-CBF-COR pathway are conserved in perennial plants, the response to long term cold treatment has shown to be different between perennial plants and Arabidopsis. Thus, the detailed analysis of responses against low temperature in perennial plant, especially overwintering phenotype as its characteristic trait, will provide the new insights of cold stress responses in plants. Lotus japonicus is a perennial legume plant, which have been used as experimental model of legume plants with a variety of material resources, including a collection of Japanese wild accessions facilitated to carry out GWAS analysis. Based on the accumulated SNPs information, it was speculated that wild accessions of L. japonicus were colonized in Japan around 25,000 years ago in Kyushu island, then divided into three subpopulation and gradually spread dispersed to high latitude area along the end of glacial era (~10,000 years ago)6. Considering this estimation, wild accessions of L. japonicus is considered to be a suitable for the analysis on local adaptation based on overwintering trait, and field experiment in Tohoku University field would also fit for the evaluation of overwintering phenotype. With the aim to elucidate the molecular mechanism of overwintering trait in perennial plant with a relation to local adaptation, I carried out molecular genetic approaches by using wild accessions of L. japonicus.
Kashimadai, Miyagi prefecture. I found significant differences of overwintering phenotypes among the wild accessions with strong latitude dependency as well as subpopulation dependency. Furthermore, the northern accessions displayed higher winter survival rates than southern accessions, and accessions classified in subpopulation 3 displayed higher winter survival rates than the other subpopulations. As a result of GWAS using overwintering phenotype in 2014-2015, two genomic loci were displayed a highly significant association with the phenotypes, and members of receptor-like protein kinases (RLKs), LjFER and LjSRK, were identified in these genetic loci on chr1 and chr6, respectively. LjFER encodes a protein very similar to the Arabidopsis FERONIA receptor-like kinase, and LjSRK encodes a putative G-type lectin S-receptor like serine/threonine protein kinase. On LjFER, six SNPs are identified in the 5’UTR, which compose two haplotypes LjFER_Ref and LjFER_Alt in Japanese wild accessions of L. japonicus analysed so far. Both of the haplotypes were identified in three subpopulations, with LjFER_Ref is a majority in subpopulation 1 and subpopulation 2, while LjFER_Alt associate with higher survival rate (winter-hardiness) in the majority of subpopulation 3. On LjSRK, three SNPs are identified on coding region, two of which are non-synonymous. Three haplotypes,
LjSRK_Hap1, LjSRK_Hap2 and LjSRK_Hap3, are composed of these SNPs, of which LjSRK_Hap2 and LjSRK_Hap3 associated winter-hardiness are identified only in the
subpopulation 3, suggesting that mutations causing LjSRK_Hap2 and LjSRK_Hap3 occurred after subpopulations were established. Additionally, these detected genomic loci were identified as genome regions with high fixation index (FST) level in comparison of subpopulation 3
occupied northern Japan and other subpopulations. This overlap between top GWA SNPs for overwintering and genome-wide top FST signal indicates that winter-hardiness haplotypes in
LjFER and LjSRK provided adaptive roles during colonization to higher latitude in Japan that has
low temperature in winter.
To investigate the roles of LjFER and LjSRK in winter-hardiness, the functional analysis was carried out using retrotransposon LORE1 insertion tag lines. As the result, the phenotypes evaluation of homozygous LjSRK_LORE1 confirmed the gene function in winter-hardiness. For
LjFER, I could not obtain the homozygous insertion lines, however, I could test the phenotype
since significant reduction of gene expression level was detected among the heterozygous
LjFER_LORE1 lines. Thus, it was confirmed that each of LjFER and LjSRK contributed in
establishing of winter-hardiness in wild accessions of L. japonicus.
By the comparative analysis of the accessions with different combination of haplotypes of LjFER and LjSRK, the effect of each winter-hardiness haplotype was confirmed in Tohoku field and additive effect of two winter-hardiness haplotypes was also confirmed. Further analysis
0 0.2 0.4 0.6 0.8 1 S u rv iv a l ra te w1-w4 w2-w5 w3-w6 w4-w7 w5-w8 w6-w9 w7-w10 w8-w11 w9-12 w10-13 w11-w14 w12-w15 0 0.2 0.4 0.6 0.8 1 S u rv iv a l ra te w1-w4 w2-w5 w3-w6 w4-w7 w5-w8 w6-w9 w7-w10 w8-w11 w9-12 w10-13 w11-w14 w12-w15
of haplotype effect of LjFER and LjSRK in winter-hardiness was carried out in greenhouse by using 23 wild accessions selected by considering the haplotypes combination as well as geographical and subpopulation relationship. The greenhouse experiment repeated weekly from late December to the early April, in which 5 weeks-old plants grown in greenhouse were placed in natural winter environment for 4 weeks then returned to greenhouse for two weeks to evaluate the phenotype. The results of greenhouse experiment is summarized in Figure 1. These result indicated that winter-hardiness haplotypes of LjFER and LjSRK increase the survival rate especially in middle of experimental period (mid-winter). Moreover, the greenhouse experiment also revealed the effect of low temperature level, latitude distributions, and L. japonicus subpopulations to survival determination of L. japonicus.
To analyze the cause of functional differences depending on the haplotypes of LjFER and
LjSRK, expression analysis was performed using 23 selected wild accessions. Results of gene
expression analysis by real-time qPCR using the RNA samples collected from field growing accessions in five time points (August, October, December, February, and March) revealed a significant up-regulation of LjFER and LjSRK during winter time in the accessions with each winter-hardiness haplotypes. Moreover, the correlation between expression level in mid-winter (December and February) and winter-survival rate were detected in each gene. Thus the significance of winter-hardiness haplotypes of LjFER and LjSRK is considered to be their potential to induce the high expression in winter season. By the cold treatment test in the growth chamber, it was confirmed that the low temperature, not the photoperiod, triggered the up-regulation of LjFER and LjSRK in winter-hardiness haplotypes. Moreover, result from greenhouse experiment revealed that gene up-regulation of winter-hardiness haplotype of LjSRK basically detected during tested period, while up-regulation of winter-hardiness haplotype of
LjFER observed in middle of the tested period with low temperature dependency. This
observation resembled to the field time course analysis suggesting the different functional timing in contribution to winter-hardiness.
Figure 1. Survival rates of 23 selected accessions in the greenhouse experiments. The greenhouse experiments
were repeated weekly from late Dec to early Apr. The five-week old plants grown in the greenhouse were treated with natural winter temperature for four weeks, then placed to greenhouse for two weeks for recovery. The graph indicates the survival rate of each week. The accessions were selected by considering the haplotypes combination as well as geographical and subpopulation relationship, these information was listed in the bottom part of the graph. Red lines indicates the accessions which has similar distribution location.
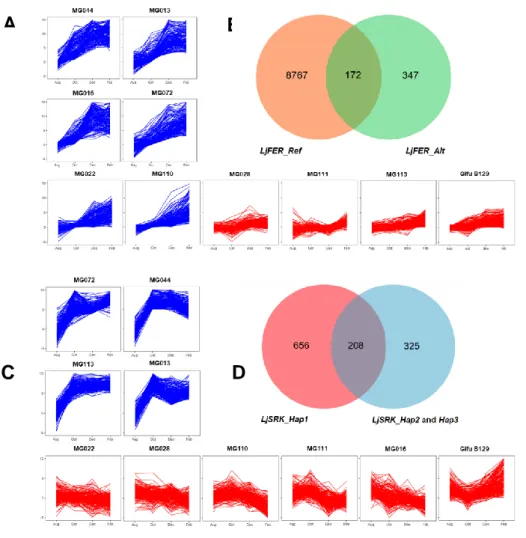
To elucidate the haplotype dependent effect of LjFER and LjSRK in the regulatory network, co-expression analysis was carried out using the RNA-seq data set on field samples collected from summer to mid-winter. A total of 172 and 208 genes were identified as co-expressed gene with LjFER and LjSRK, respectively (Figure 2). Although none of the co-expressed genes of LjFER and LjSRK were overlapped, GO term enrichment analysis revealed that ten GO terms, including membrane activity and cell wall metabolism, were commonly enriched in co-expressed genes in both LjFER and LjSRK. In addition, genes related to lipid metabolism were co-expressed with LjSRK, while genes related to carbohydrate metabolism as well as components of ICE1-CBF pathway were clustered in LjFER. These results suggested that upregulation of LjFER and LjSRK in winter season long depending on the winter-hardiness haplotypes lead to the up-regulation of the different set of genes that contribute to enhance the physical toughness against harsh winter.
In this study, I revealed the novel insights of molecular mechanisms in local adaptation of
L. japonicus to high latitude area with low temperature. By gaining winter-hardiness haplotypes
in two receptor kinase genes, LjFER and LjSRK, up-regulation of the set of genes that provide protection against harsh winter including cell wall and cell membrane metabolism, lipid metabolism, carbohydrate metabolism, together with the activity of ICE1-CBF regulon were established during the winter season long. For perennial plant such as L. japonicus, the enhancement of tolerance in terms of surviving to the harsh of winter climate could have been critical for local adaptation to high latitude area. Moreover, the identified receptor kinase genes,
LjFER and LjSRK may be considered as the master regulators of genes that contribute to the
enhancement of winter-hardiness. Further analysis of these genes will contribute to the understanding of the environmental adaptation in the perennial plants. In addition, orthologues of
LjFER and LjSRK will be good targets for improvement of temperature responses of
B
Figure 2. The co-expressed genes with LjFER and LjSRK haplotypes. The co-expressed genes were analyzed
using RNA-seq data based on the clustering analysis by comparing the gene expression trend against expression pattern of LjFER and LjSRK during qRT-PCR analysis. The line graphs (A: LjFER, C: LjSRK) indicate the expression level of co-expressed genes in ten accessions. Red graph indicates winter-susceptive haplotypes and blue indicate winter-hardiness haplotypes. In venn diagram (B: LjFER, D: LjSRK) each circle element represents number of conserved co-expression genes in the LjFER and LjSRK haplotypes and the overlapped area indicate the numbers of co-expressed genes of LjFER and LjSRK, respectively.
References:
1. Körner, C. Plant adaptation to cold climates. F1000research. 5, 1-5 (2016).
2. Preston, J.C., & Sandve, S.R. Adaptation to seasonality and the winter freeze. Front. Plant Sci. 4, 1-17 (2013). 3. Wingler, A. Comparison of signalling interactions determining annual and perennial plant growth in response to
low temperature. Front. Plant Sci. 5, 1-9 (2015).
4. Guo, X., Liu, D., & Chong, K. Cold signalling in plants: Insight into mechanisms and regulation. J Integr Plant
Biol. 60, 745-756 (2018).
5. Liu, Y., Dang, P., Liu, L., & He, C. Cold acclimation by CBF-COR pathway in a changing climate: Lessons from
Arabidopsis thaliana. Plant Cell Rep. 38, 511-519 (2019).
6. Shah, N., Wakabayashi, T., Kawamura, Y., et al. Extreme genetic signatures of local adaptation during Lotus
japonicus colonization of Japan. Nat. Commun. 11, 1-15 (2020).
A
論文審査結果の要旨 植物は、周囲の環境を感知し順応するための機構を発達させており、分布域を広げる際 には、新たな環境要因に適応するために、それらの機構を変更してきた。植物の発生や生 育を阻害する要因となる低温はそのような環境要因の一つであり、特に。多年生植物にと っては環境適応の重要な要因となる。 Yusdar Mustamin 氏は、多年生植物が地域適応の過程で獲得した耐冬性の分子メカニズ ムを解明することを目的として、マメ科の多年生植物であるミヤコグサを用いた集団ゲノ ミクス、分子遺伝学解析を行なった。日本各地から採取した 120 種のミヤコグサ系統を東 北大鹿島台圃場で栽培することにより得た越冬性の表現型を用いたゲノムワイド関連解析 (GWAS)の結果、2 種類の受容体型キナーゼ、LjFERとLjSRKのハプロタイプと表現型と の相関を同定した。ミヤコグサ集団におけるこれら 2 遺伝子のハプロタイプの組み合わせ と越冬性、低温傷害耐性の関係を詳細に解析した結果、LjFERとLjSRKそれぞれの耐冬性 型ハプロタイプは相加的に耐冬性を上昇される効果があることを見出すとともに、高緯度 地域への適応に重要な役割を果たしたことを示唆する結果を得た。耐冬性型ハプロタイプ のLjFERとLjSRKの遺伝子発現への影響を調査した結果、各遺伝子の耐冬性型ハプロタ イプを持つ系統では、冬季の低温によりそれぞれの遺伝子の発現上昇が認められ、冬季期 間中高いレベルの遺伝子発現が維持されることを発見した。加えて、異なるハプロタイプ の組み合わせを持つ10 系統を用いて RNA-seq 解析を実施し、LjFERとLjSRKの遺伝子 発現変動と同じ変動パターンを示す遺伝子群を検索した結果、共に低温時の細胞保護のた めに機能する遺伝子群が共発現することを見出した。このように、一連の研究により、ミ ヤコグサの高緯度地域の適応に重要な役割を果たしたハプロタイプにより LjFER と LjSRKの冬季発現上昇がもたらされ、それが一連の耐冬性関連遺伝子の発現につながるこ とを明らかにした。 以上の研究成果は、博士論文提出者が自立して研究活動を行うのに必要な高度の研究能力 と学識を有することを示している。したがって、Yusdar Mustamin 氏提出の論文は,博士 (生命科学)の学位論文として合格と認める。