Tech Bull Fac Agr Kagawa Univ Vol 35, No 2 61-68, 1984
Peroxidase Isozymic Variation in Some Faba Bean Strains
(Vzcia
faba
L.)
Kiyoshi
YAMAMOTO
In order to clarify the phylogenetic and genecological relationship between some strains of faba bean (Vicia faba L.) which were introduced from different countries, variation of peroxidase isozymic band patterns were examined.
A total of twenty six strains used for this experiment were introduced from Japan, Spain and Finland. These strains were classified into five according to their native country and their seed size.
Peroxidase isozyme band patterns were detected by isoelectroforcusing method.
Other enzyme systems of Esterase, GOT and IPO were also detected in this experiment, but no difference in their isozyme band patterns was observed between the all strains used
A total of sixteen peroxidase bands were detected, of these, eight bands were in minus zone and another eight were in plus zone. Similarity index values were culculated between all strains experimented This values were high between Japanese strains regardless in their seed size and low in Spanish and Finnish strains. From the result, it is deduced that Japanese strains were more genetically homologous than those of the Spanish and Finnish strains It was also deduced that Japanese faba bean strains experimented were more specialized phylogenetically than those of Spanish and Finnish ones This inclination was also correspond to the speciality of their morphological characters observed.
./ 4 . 7 ~ & @ D & @ t * 4 k D e i a . % # % ? 6 f : & , El*, Y k j k . d 7 4 Y ? Y b?5iG%ALf:&SI26%%CZ
->~,\-c,
~A%t@FDk$!S;5iG ~ T D % % % I C ~ ~ % L ~ : ~ r h ~ ~ ~ ~ r r - > b ~ - c / ~ - j s ; f v ~ - - f i " ~ 7 4 ~ 9A 4,*
,
.
p' %,sw Z 2 3D%ti4ib~@!4L%~djln"c?r'T%%Lf:o13i1zlr:s.z.~4.-.+i', GOT B L d IPO ~ 7 4 ' / 4 4 ~ / * ~ b?~&ifl%.6-,f:5s, & % , % ~ 3 . ~ f : ( $ $ W h s l b 5 GhfJ5z.3 f:.
)q,-$;f Y9',-+'D/*Y Fbi&%%T6316&&ifl S h , + h G D
5
% C c i i % A % % L d @ T D A 3 S CCL 633%RT'T1@?&D%%l& G h 6 / T Y b'&& 9 f:o S GGf&tlj $ hf:26%%?<TCZ->L\T, 2 TD%,%D$ii&?CCL6/*Y F
~ 9 9
-
YDEM%,Z?
Similarity index (S. I.) %,$&, SG
Cr:
r
~ l 6 5+
hQhD%,%% t*t
Ir:
3t
&~%,i'glB%,3@
L, . k @ L f:o' 8 t L l n 5 ,
El$D%,%DPdTli@FDhB St
RMRGT:
S. I. D 4 6 b i ~ . * 4 YD%%Pd%Ld74 7 . 7 Y F D % % P ~ T D ~ E L
9
%f:aAi, ~ 4~ k j & d 7 4 4 Y ? Y b ? g % , % ~ a ~ P d ~ r i + ~ l E 5 f 4 &( ,
t
<
1 r : ~ l 2 i x % , ~ t x 4 4 ~ k ; ~ u a $ % % 2 7 4 7 5 7b ? r ~ q ~
s..I.
Dutig&l&fii.,f:o7 4 YY.4 A/*? b?D%{ZF3&?&gz?6 2 , B$D%,@?dTCiz44 Yk;L6$7 4 7 7 Y ~ ? D ~ & , B P B ~ L
9
&a
iZLrcl%,@I&iSrS<
,
;b~->&@~fi1C;hrk9 B h ~ b \ 6
t%Er?: h f z or
~fiW11-d: ~ l ~ % , % ~ f i % % f g F ~ & & I 4 , t&I#
&Y& 6 .A
,5
IC,E\hh6 0Introduction
62 Tech Bull Fac Agr Kagawa Univ Vol 35, No 2 (1984)
animal foods This cultivars, large variations in their seed size are observed. Of these, large seed sized varieties are so called V faba major"'. This varieties are commonly cultivated in Japan for vegetable@) On the other hand, middle (so called V. faba equina) and small sized varieties ( K faba mznor) are widely cultivated in Europian, Meditarranean, African and American countries both used for human foodand animal food(1,Z,5.9). The faba bean is considered to have its orgin in the Neolithic Age and was well known to and many varieties a r e derived and its phylogenetic differentiation is supposed ~omplicate'~' Not so many studies on this problem were carried out until now. On the evolution of the faba bean, some studies were done'2~4~9~19.20', but not yet clear in the present time. From this standpoint, it is important to clarify the ecogenetic and phylogenetic relationships between the cultivated strains introduced from different countries.
Many phylogenetic and genecological studies by the isozyme band patterns were carried o ~ t ( ~ . ~ ~ ~ ~ ~ . ~ ~ ~ ~ ~ . 1 6 , 1 8 3 1 9 3 2 0 ) . But not so many studies on this problem in the faba bean cultivars were done. So, this study
of isozymic band patterns in these faba bean strains has been aimed to clarify the phylogeneticand genecological relationship.
Materials and Methods
Seed sources of the twenty six strains used for this experiment are listed up in Table 1. Of these, nine Table 1. Seed source of faba bean strains used for experiment
Strain No Strain name Seed source
1 Sanuki-nagasaya-1 Nagata Seed Co. Kagawa 1976
2 Kawachi-isun // /I Osaka //
3 Rikusei-isun Kowa Seed Co Tokyo N
4 Kawachi-isun Tokushima Pref E x p S t a N
5 Newly breeded strain // N N U 0
6 Sanuki-nagasaya-2 Hashimoto Seed Co Kagawa 1975
7 INIA 02 NO RTO 109 Mo-sagaua Spain 1976
8 INIA 04 NO RTO 99 M-E Spain 1976
9 INIA 05 NO RTO 115 Sevillana Spain //
10 INIA 06 NO RTO 143 M-E Spain //
11 NO RTO 649 Wseis BEAGLE N n
12 NO RTO 650 Waris BLAZE 11 //
13 NO RTO 172 d a Coaq porlorue t o grand //
Pirhonen (Horse bean) Old Finnish vareity Finland
Hja 60182 (horse bean) //
N 60189 // N
// 70001 // local strain 0 Nvijamas
0 70005 f l local vareity N Savitaipale
// 70006 N local strain 0 Savitaipale
N 70011 // local strain N Parikkak
N 70021 //
N 70025 N local strain 0 Parikkak
N 72001 0 local strain 0 Vihti
24 Otaf u k u bean
25 Gaeru-mame (from China)
26 Hajiki-mame
Hashimoto Seed Co. 1977
Nagata Seed Co. 0
Kiyoshi YAMAMOTO : Peroxidase Isozymic Variation in Faba Bean Strains 63 Table 2. Methods of isozyme detection
Enzymes Substrate and staining dye
Esterase Hz0
0.12 phosphate buffer pH 6.1 1% a-naphtyl acetate First blue RR salt
IPO* 0 03M Tris-HCI buffer pH 8.0 1M MgClz
Phenazin methosulf ate MTT tetrazolium
GOT** 0.1M Tris-HC1 buffer pH 8.0 Pyridoxal-5-phosphate L-aspartic acid a-ketoglutar ic acid First blue BB salt Peroxidase 0-dianisidene CH3COOH Hz0 0.2 Acetate buffer pH 4.8 0.06% H202-Acetate buffer 60m1
/
a t 3 7 0 ~ for 45 min 2ml l m l(
a t room ternp for 1.5 hr lOmg
I
0.84mg
70ml a t room t e m p for 10 min lomg
J
*
IPO : Indophenol oxidase**
GOT : Glutamic-Oxaloacetic transaminaselOOml '
5mq 100mg lOOmg 150mg
,
strains from No. 1 to 6 and No 24 to 26 were introduced from Japan, Of these strains only one of No 25 was Chinese origin Seven strains from No. 7 to 13 were introduced from Spain Another 10 strains from No. 14 to 23 were from Finland a s shown in the table.
Isozyme detection was done using the polyacrylamide slab isoelectroforcusing method for peroxidase and disc electrophoretic method was done for Esterase, GOT (Gultamic-oxaloacetic transaminase) and IPO (Indophenol oxidase). Staining methods were applied from B r e ~ e r ' ~ ) . The leaves used isozymic screening in detail were taken a s reported p r e v i o ~ s l y ( ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ . Leaf extructs were made by grinding a mixture of 2 grams of leaflets with the same weight of buffer solution (pH 7.54). Substrate and staining dyes used for each enzyme systems are listed in Table 2.
) a t 37OC for 30 min
Results and Discussion
lOOmg )
I Seed size and some characters
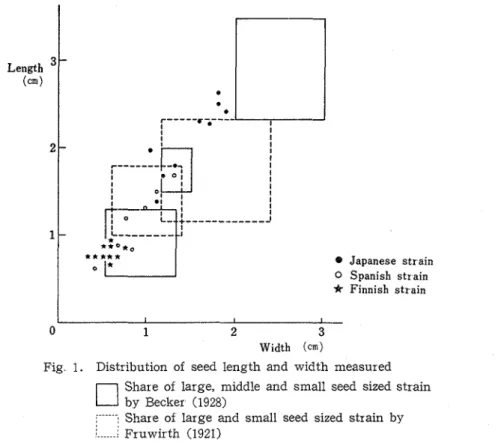
Much variation of seed size among the strains was observed in this experiment. Distribution of seed length (in vertical) and width (in horizontal) in each strains measured a r e shown in Figure 1. Additionally, the range of seed length and width classified to large ( V faba major), middle (% faba equzna) and small ( V faba r n z n ~ r ) ' ~ ~ ~ ~ ~ ) are also indicated in the same figure As shown in the same figure, clear segregation was observed among the strains and classified into large, middle and small sized strains in their seeds, however, some differences were demonstrated in the share of seedsize between the strains in this measurement and the share indicated by BECKER'~' and FRUWIRTH'~'. From this classification, 5 strains were large sized
(V
faba major), 4 were middle ( V faba equina) in Japanese strain. In Spanish strain, 4 were middle and 3 were small ( K faba mznor) On the other hand, in Finnish strain, all of the 10 strains were small sized.Tech. Bull Eac Agr Kagawa Univ. Vol. 35, No 2 (1984) e J a p a n e s e s t r a i n 0 S p a n i s h s t r a i n
I
*
~ i n n i s h s t r a i n 0 1 2 3 Width (cm)Fig. 1. Distribution of seed length and width measured Share of large, middle and small seed sized strain by Becker (1928)
i----'-: Share of large and small seed sized strain by 8 <
:..--..1 Fruwisth (1921)
varied from large to small Minus correlation between seed length and number of node in Japanese, Spanish and Finnish strains was observed In general, the number of node was few in Japanese strain and many in Finnish strain. In Spanish strain, i t was intermediate. Pod size was also varied and small in Finnish strain On the other hand, it was large in Japanese one The number of seeds per pod was many in Finnish strain but in Japanese strain, it was few. The color of spot in wings of flower varied from gray to purple, stem was reddish in almost Japanese strain In spanish strain, they were more variable between the strains than those of the Japanese and Finnish strains. In Finnish strain, i t has waved in the edge of leaflet. From the seed size and morphological characters above mentioned, Japanese strain was different from Spanish and Finnish strains, and so it is presumed that Japanese strains are specific cultivars among the faba bean vareities.
I1 Peroxidase isozymic variation
Presence of peroxidase isozymic bands detected in 26 strains are shown in Table 3.. As shown in the table, a total of 16 bands detected, of these, 8 were in minus zone (each bands were named from -1 to -8 according to their Rf value) and another 8 bands were detected in plus zone (which is also named from
+
1 to+
8). Of these 16 bands, No.-
3 and -5 were common in all strains examined, and band No. .-I, -7, + 3 and + 6 were also detected in almost all the strains. Band No. -2 was observed in all ,Japanese strains. But it was also observed in small sized Spanish and almost Finnish strains. Band No. -8 was only detected in .Japanese strains, on the other hand, No. +1 was .Japanese and almost all of Spanish strains. Characteristic isozyme bands in the related species of the genusVicza
were also reported by a ~ t h o r ( ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ) . But in this experiment, identification of strains by isozymic bands was not so easy. For clarify the difference of band patterns between the strains, similarity index values among all strains combined were culculated according to the next formula.Kiyoshi YAMAMOTO : Peroxidase Isozymic Variation i n Faba Bean Strains 65 Table 3. Peroxidase isozyme bands detected
Similarity index value between the strains.. (S. I.) =
-band +band
Strain ~ ~ ~ " ! 1 ! 2 ~ 3 ~ 4 / 5 ~ 6 ~ 7 ~ 8 ~ 1 ~ 2 ~ 3 ~ 4 / 5 ~ 6 ~ 7 ~ 8
-- Number of common bands in both strains combined
Total number of common and non common bands in both strains combined. Japanese strain Spanish strain Finnish strain Note
+
:These values are shown in Table 4. On the other hand, the mean values of similarity index classified according to their seed source and seed size are also indicated in Table 5.
As shown in both tables, similarity index values between Japanese strains varied from 0.67 to 0 91, and 0 77 in mean between the combination of large and large seed sized strain, from 0 67 to 0 90, and 0 76 in mean between middle and middle seed sized combination Both values were the highest in all strains combined in this experiment. The next highest similarity index value was the strain combinations between large and middle seed sized strains I t ranged from 0 58 to 0.90 and the mean value was 0 72. In Spanish strains, the range of this values in combination of middle and middle, small and small were from 0 50 to 0 75 and 0 58 to 0 75 respectively And their mean were 0 61 and 0 67 On the other hand, this values between middle and small were ranged from 0 45 to 0 69 and their mean value was 0 56
In Finnish strains, the range of this values of the strain combinations of small and small were ranged Large sized Middle sized Middle sized Small sized Small sized band detected, - 2 4 3 24 25 26 7 8 9 10 11 12 13 14 15 16 17 18 19 21 22 23 :
+
I +--
+
+
+
.+
+
+
+
+
+
,+
+
+.
+
+
2 0 + + + i-.+
+
band+
+
+
5 - + . +-+
6 . + + ++
-+.
+
- - -+,
.-I- detected+
.+
.+
+
+
+.
+
,+
+
+.
+
t+
+
+.
+,
+.
+
3.+
-+.
-
.- - - .--
, + + - I - + but+
+.
+
+
.t .f -t +.+
.+
t-
+
+
.+
+
+
+ + + . + +
+
+
+
.+
-
thin,.+
-+
+
+.
+
+
+ + -
+
intermediate -+ + -
- -- T+
-+
-+
+
-+
.+
+
+
+
-+
+
+.
-I-+
+
:-
.+
,+
+
+
+
+.
+
+
+
-T
.+
+
+
f+.
+.
+
.+
+
+
+
+.
+.
.t+
+
3.+.
+.
in+
+
+
+
+
+
+
+
thickness-
+
4- .t-
.+
,-I- --
+.
t.-
-+
+
+
+
.f+
+
+
+
+
+
+
f+
+
+
+
+
-
,++
-
+
+
+
.t+
+
-
-Table 4. Similarity Index values between the stralns
Kiyoshi YAMAMOTO : Peroxidase Isozymic Variation i n Faba Bean Strains
from 0.44 to 1 00 and was 0 66 in mean
In other combinations between Japanese, Spanish and Finnish strains, these similarity index values were all lower than those of the above mentioned Japanese and Japanese, Spanish and Spanish and Finnish and Finnish strain combinations. And the similarity index values were the lowest in both combinations of Japanese strains and Spanish, Japanese and Finnish strains regardless in their seed size..
On the other Esterase, GOT and IPO enzyme systems carried out in this experiment, no difference in their band patterns were observed among the all strains experimented, and so no discussion on these enzyme systems was done
Relationship between isozymic band pattern and genetic similarity were demonstrated by many previous
r e p ~ r t s ( ~ ~ , ~ ~ ~ ~ ~ . ~ ~ ~ ~ ~ . ~ ~ ) . In this experiment, genetic variation presumed by peroxidase isozymic bands were larger in Spanish and Finnish strains than that of the Japanese ones. On the other hand, similarity index values between large and large, middle and middle, small and small seed sized combinations were higher than those of the combinations between the strains which have different seed size. From the results, it is presumed that genetic affinity among Japanese strains a r e the highest This inclination is deduced from their morphological characters and it is also deduced that Japanese faba bean strains experimented are more specialized phylogenetically than those of Spanish and Finnish ones This inclination also cor- responded to the speciality of their morphological characters observed
Table 5. Mean and standard diviation of similarity index values between the strain groups classified according t o t h e seed source and seed size
Literature Cited
( 1 ) BOND, D. A.: The development of field beans a s a clop in B~itain, Proc Nutr Soc , 29 (I), 74-79 (1970)
( 2 ) BECKER-DILLINGEN, J : Handbuch des Hulsenf r uchterbaues und Futterbaues, (1928)
( 3 ) BREWER, G J : An introduction to isozyme techniques, Academic Press, New York (1970).
( 4 ) CUBERO J I : On the evolution of Vicza faba
Finnish strain Small sized
0.66h0.16
L., Theoretical and Applied Genetics, 45,
47-51 (1974)
.
( 5 ) EHARA, K.. : Forage Crops I, Yokendo, Tokyo (1950), (in Japanese).
( 6 ) FRUWIRTH, G.. : Handbuchs des Hulsenfrucht- baues, (1921).
( 7 ) KATAYAMA, T,., CHERN, J.. L. : Zymographic studies on diploid 0r:y.z~ punctata and its related species, ,Japan.
,I:
Breed, 23, 329-333 Seed source andSeed size
%
$
ag
s
a2
G 2iZ
Largesized Middle sized Middle sized Small sized Small sized ,Japanese strain Large sized(
Middle sized0.7710.07 0.72h0 09 0.49h0.12 0.5110.08 0.511.0.11 Spainish strain Middle sized
(
Small sized0.76ir0.07 0.5610.08 0.591.0.06 0.5510.11 0.61k0.08 0.5610.10 0.622~0.13 0.67k0.07 0.531. 0.09
68 Tech Bull Fac Agr Kagawa Univ Vol 35, No 2 (1984) (1973)
( 8 ) KUMAZAWA, S. : Vegetable Crops, Yokendo, Tokyo (1956), (in Japanese).
( 9 ) LADIZINSKY, G : On the origin of the broad bean, Israel J B o t , 24, 80-87 (1975) (10) NAKAGAWARA, M. : Genetic analysis for
isozymes in rice cultures, Japan J Breed, 27, 141-148 (1977)
(11) NAKAI, Y., TSUNEWAKI, K. : Isozyme variations in Aegzlops and Trzticum I. Esterase isozymes in Aegzlops studied using the gel isoelectro- forcusing method, Japan J Genetzcs 46,321- 336 (1971).
(12) NISHIKAWA, K t NOBUHARA, M. : Genetic studies of a-amylase isozymes in wheat, Japan J Genetzcs, 46, 345-353 (1971)
(13) PLITMANN, U : Biosystimatical study in the annual species of Vicza of the Middle East, PhD Thesis, The Hebrew University of Jerusalem (1967)
(14) SHAHI, B. B., CHU, Y E., OKA, H. : Analysis of genes contorolling peroxidase isozymes in Oryza satiua and 0. perennzs, Japan J Genetzcs, 44, 321-328 (1969)
(15) SHANON, L. M. : Plant Isozymes, A n n Rev
Plant Physzol, 19, 187-210 (1968)
(16) YAMAMOTO, K.: Estimation of genetic homogeneity by isozymes from interspecific hybrids of Vzcza I Amylase isozyme patterns in the hybrid progeneis between Vicia pzlosa and
V.
macrocarpa, Japan J Breed,25 (I), 60-64 (1975)
(17) - : Estimation of genetic homogeneity by isozymes from interspecific hybrids of Vzcia I1 Amylase isozyme patterns in the hybrid progenies between Vicza amphzcarpa, true and
V.
macrocarpa, Japan J Breed, 29 (I), 59- 65 (1979)(18) YAMAMOTO, K., PLITMANN, U.: Isozyme polymorphism in species of the genus Vzcza (Legumznosae), Japan J Genetzcs, 55, 151-164 (1980)
(19) YAMAMOTO, K., MORITANI, O., ANDO, A. :
Karyotypic and isozymatic polymorphism in species of the section Faba (genus Vzcza), Tech Bull Fac Agr Kagawa
U ,
34 (I), 1- 12 (1982)(20) ZOHARY, D.: Comments of the origin of cultivated broad bean, Vzcza faba L., Israel J B0t