糖 タンパ ク質糖鎖 の構造 と機能
木
幡
陽 *
Structures
and
Functions
of the Sugar
Chains
of Glycoproteins
Akira KOBATA *
Most proteins within living organisms contain sugar chains. Recent advancement of cell biology has revealed
that many of these sugar chains play important roles as signals for cell surface recognition phenomena in
mul-ti-cellular organisms. Based on an idea to elucidate the biological informations included in the sugar chains and
introduce them as knowledge of biology , a novel scientific field called "glycobiology" was established. This
re-view will give an outline of the structural characteristics and biosynthetic mechanism of the Asn-linked sugar
chains of glycoproteins. Based on these knowledges , functional aspects of the sugar chains in the immune
sys-tem of mammals, and of those of human chorionic gonadotropin will be described in order to help readers to
ac-cess this new scientific field.
Key words : Glycoprotein ; Asn-linked sugar chains ; Microheterogeneity ; Concanavalin A ; Exoglycosidase ;
Bisecting GlcNAc ; Processing of sugar chains ; N-Acetylglucosaminyltransferases ;
Humancho-rionic gonadotropin ; Immunoglobulin G.
は じめ に
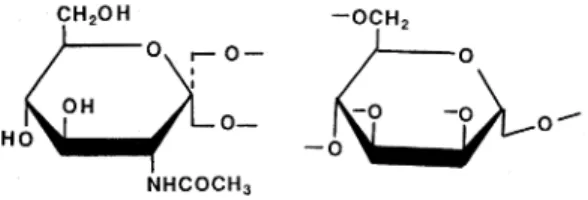
糖 タ ンパ ク 質 とは 単糖 の つ な が っ て で きた糖 鎖 が,タ ンパ ク質 に共 有 結 合 して い る もの の総 称 で あ る。 動 物 の 細 胞 が 作 り出 す タ ンパ ク質 の 多 くは糖 タ ンパ ク 質 と して 存 在 して い るが,そ の 生 理 機 能 を考 え る 上 で糖 鎖 の部 分 に も研 究 者 の 目が 向 け られ る よ う にな った の は そ う古 い こ とで は な い 。 こ れ はmRNAと い う鋳 型 の 介 在 に よっ て作 り出 され る タ ンパ ク質 と は異 な り,糖 鎖 部 分 に は構 造 上 の ミク ロ 不 均 一 性(microheterogeneity)が 存 在 す る た め に,そ の構 造 解 析 が きわ めて 困難 で あ った こ と と, ア ミ ノ酸 が一 本 の 糸 と してつ なが っ て い る タ ンパ ク質 と 違 って糖 鎖 に は枝 分 れ構 造 が あ り,比 較 的少 ない 数 の 単 糖 ユ ニ ッ トで作 り出 され る構 造 が理 論 的 に き わめ て 多 様 で あ った た め で あ る。 今,鎖 の最 少 単 位 で あ る二 つ の ユ ニ ッ トの 結 合(A-B)を と り上 げ て み よ う。 タ ンパ ク質 で はAを ア ラニ ン,Bを ロ イ シ ン とい うふ う に決 め る と, そ の構 造 は 一 種 類 しか な い。 とこ ろ が糖 鎖 の場 合 は, A をN-ア セ チ ル グ ル コサ ミ ン,Bを マ ン ノ ー ス と決 め る と,図1に 示 す よ う にN-ア セ チ ル グ ル コ サ ミ ン は マ ン ノ ー ス のC-2, C-3, C-4, C-6位 と4つ の 位 置 に 結 合 し う る か ら,4種 の 異 性 体 が 可 能 と な る。 ま た,N- ア セ チ ル グ ル コ サ ミ ン の 結 合 に は α と β の 二 つ の ア ノ マ ー構 造 が 可 能 で あ る の で,作 り出 され る構 造 は 4×2 =8種 類 とな る。 さ ら にN-ア セ チ ル グ ル コ サ ミン は 図 1に 示 した ピ ラ ノー ス 型 以外 に フ ラ ノー ス構 造 も と り う る の で,GlcNAc→Manと い う2糖 に は合 計16種 の 異 性 体 が 存 在 しう る こ とに な るわ け で あ る 。 ユ ニ ッ トの 数 を さ らに 三 つ,四 つ と増 や して い く と,タ ンパ ク質 の 場 合 は直 鎖 で あ るた め 相 変 らず1種 類 の構 造 しか作 り出 さ れ ない が,糖 鎖 の 場 合 は 枝 分 れ が 可 能 で あ る た め に,異 性 体 の 数 は幾 何 級 数 的 に増 加 して い く こ とに な る。 この よ う な構 造 的 特 徴 の 違 い が 糖 鎖 の構 造 解析 をお くらせ て き た ので あ った 。* 東 京 大学 医科 学研 究 所細 胞生 物化 学研 究部
* Department of Biochemistry, the Institute of Medical Science , the University of Tokyo
Fig. 1 Construction of sugar chains
452 有機 合 成 化 学 第50巻 第5号 (1992)
( 76 )
しか し1970年 ご ろ か ら急 速 に 進 展 しは じめ た細 胞 生 物 学 の 研 究 に よ って,間 接 的 な事 実 で は あ るが糖 タ ンパ ク質 の糖 鎖 が細 胞 間 識 別 の シ グ ナ ル の役 割 を果 して い る こ と を示 す 知見 が色 々 と報 告 さ れ は じめ ,糖 タンパ ク質 糖 鎖 の機 能解 明 を求 め る機 運 が 高 ま っ て きた。 こ う した 動 きに触 発 さ れ て,糖 タ ンパ ク質 糖 鎖 の構 造 を 的確 に解 明 す る た め の多 様 な分 析 技 術 が 次 々 と開 発 され た。 こ れ らの技 術 を駆 使 して 様 々 な糖 タ ンパ ク質 の 糖 鎖 構 造 が 具 体 的 に明 らか に な り,そ こ に ひそ む法 則 性 や 多 様 性が 解 明 され て い った 。 こ う した 知 見 を も とに して 糖 タ ンパ ク 質 の糖 鎖,特 にAsn-型 糖 鎖(後 述)の きわ め て 複 雑 か つ 特 異 な 生 合 成機構 が 一気 に解 明 され,糖 鎖 の 形成 調 節 の メ カ ニ ズ ム を考 え て い く土 台 が 確 立 され た の が1980 年 代 の 前 半 で あ る。 加 えて 血 清 糖 タ ンパ ク質 の ク リ ア ラ ンス の研 究 に 端 を 発 したAshwellら の 肝 細 胞 形 質 膜 の ガ ラ ク トー ス結 合 タンパ ク 質 の 発 見,Sly, Neufeld, Kornfeldら の リソ ソー
ム酵 素 の選 択 的 な細 胞 内輸 送 機 構 の 研 究 に よる Man- 6-Pレ セ プ タ ーの 発 見 な どに よっ て,細 胞 の 膜系 に結 合 し た 様 々 な レ クチ ン(糖 鎖 と結 合 す る タ ンパ ク質)が 存 在 す る こ とが 明 らか に な る に お よ び1),糖 鎖 を介 した 細 胞 表 面 で の 認 識 機構 が 仮 説 の 域 を脱 して きわ め て具 体 的 な話 とな って きた 。 さ らに 遺 伝 子 組換 え 技術 の 導 入 に よ って,そ れ まで 育 成 され た 生 物 を利 用 す る こ とに 限定 され て来 た発 酵 隼 産 技 術 が,生 物 を よ り高 度 に利 用 す る こ とが 可 能 に な り, そ れ に伴 ってバ イ オ イ ン ダス トリー とい う新 しい 産 業 分 野 が生 まれ つつ あ る 。 しか しす で に のべ た よ う に,動 物 の体 内 で作 り出 さ れ る タ ンパ ク質 の多 くは糖 タ ンパ ク質 と して存 在 して お り,こ れ を糖 鎖 付 加 能 力 の ない バ クテ リア な どに遺 伝 子 組 換 え技 術 で 作 らせ る と,し ば しば 予 想 した生 理 活 性 を示 さ な い こ とが あ る こ とか ら,改 め て 糖 タ ンパ ク質 糖 鎖 の機 能 的役 割 の 重 要 性 が 浮 上 して きて い る。 本 稿 で は,こ の 多 方 面 の要 請 か ら発 展 しつ つ あ る糖 タ ンパ ク質 糖 鎖 の 構 造 と機 能 相 関 の 問 題 を,そ の 背景 に あ る構 造 解 析 法 や 糖 鎖 の 生 合 成 機構 を含 め て 紹介 す る こ と にす る。
1. 糖 タ ンパ ク質 糖 鎖 の 構 造解 析 法 の発 展
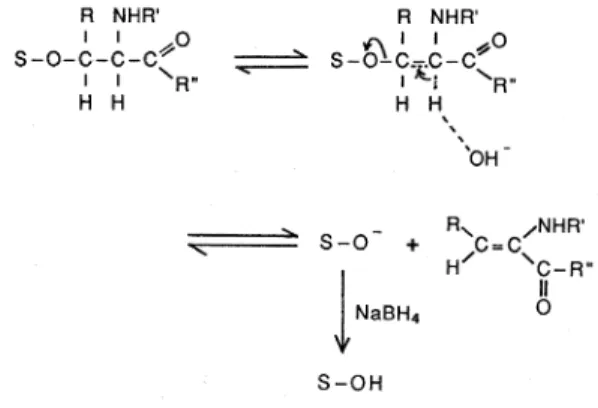
タ ンパ ク質 に 結 合 した糖 鎖 は,ポ リペ プ チ ドの ア スパ ラギ ン残 基 に結 合 して い るAsn-型 糖 鎖 と,セ リ ンや ト レ オ ニ ン残 基 に 結 合 して い る ム チ ン型 糖 鎖 に大 別 され る 。 ム チ ン型糖 鎖 に は 酵 母 の産 生 す る糖 タ ンパ ク質 の よ うな きわ め て少 数 の例 外 を除 い て マ ンノ ー スが 一 切 含 ま れ て い ない が,Asn-型 糖 鎖 には 多 か れ 少 な か れ マ ンノ ー ス が 必 ず 含 まれ て い る。 従 って,精 製 され た糖 タ ンパ ク 質 の 単糖 組 成分 析 を行 って マ ン ノー ス の有 無 を調 べ る こ とに よ り,い ず れ の糖 鎖 が 含 まれ て い る か を予 想 す る ζ とが で きる 。後 で の べ る よ う にAsn型 糖 鎖 とム チ ン型 糖 鎖 の構 造 研 究 法 は大 き く異 な る の で,こ のデ ー タ は糖 タ ンパ ク 質 の糖 鎖 構 造 の研 究 に とっ て き わめ て 重 要 で あ る。 糖 鎖 の構 造 決 定 の上 で 糖 タ ンパ ク質 が 糖 脂 質 と大 き く 異 な る点 は,後 者 が 分 子 中 に1本 の 糖 鎖 しか 含 まな い の に対 し,糖 タ ンパ ク質 の 多 くは複 数 の 糖 鎖 を持 って お り, しか もす で にの べ た よ う に糖 鎖構 造 に ミク ロ不 均 一性 が 存 在 して い る こ とで あ る。 この た め に,精 製 され た糖 タ ンパ ク質 は不 均 一 な糖 鎖 を含 ん で い る こ とに な り,構 造 決 定 の た め には まず 個 々 の 糖 鎖 を少糖 と して切 り離 し, これ を分 画 精 製 す る こ とが 必須 条件 とな る 。 ム チ ン型 糖 鎖 で は そ の 目的 の た め に古 くか らNaBH、 共存 下 で ア ル カ リ分 解 に か け て糖 鎖 を少 糖 ア ル コ ー ル と して 遊 離 す る β-脱離 反 応(β-elimination)2)が考 案 さ れ , 広 く使 わ れ て きた(図2)。 この 反 応 で はNaB3H4を 使 う こ とに よ り糖 鎖 を3H標 識 少 糖 ア ル コー ル に変 え る こ と が で き る の で,以 後 の 少 糖 の 検 出 が 楽 に な る 。 し か し β-脱離 反応 に は1Mと い う きわ め て高 濃 度 のNaBH、 が 必 要 な の で,NaB3H、 の使 用 は大 量 の ア イ ソ トー プ を 反 応 廃 棄 物 と して作 り出す こ と に な り,環 境 汚 染 の 面 か ら み て も問題 が あ る 。 最 近 天 野 ら3)はβ-脱離 反応 で遊 離 し た 少糖 ア ル コ ー ル を ヒ ドラ ジ ン と加 熱 す る こ と に よ りア ミノ糖 の ア セチ ル基 を除 き,こ れ を[3H]無 水 酢 酸 で 定 量 的 にN-ア セチ ル化 す る 方 法 を考 案 し,幾 つ か の 糖 タ ンFig. 2 ƒÀ-Elimination reaction of the mucin-type sugar
chains of a glycoprotein. S represents a sugar chain.
( 77 )
糖 タンパ ク質糖鎖 の構造 と機能
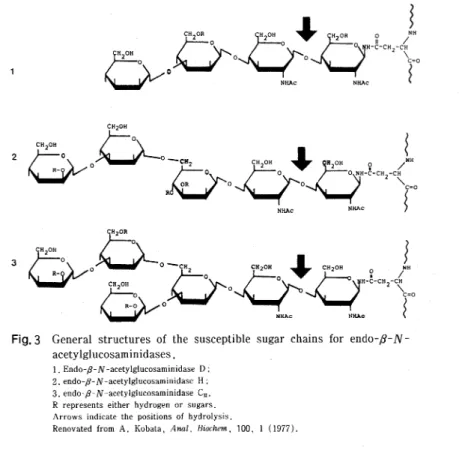
453 バ ク 質 の ム チ ン型 糖 鎖 の構 造 研 究 に 応 用 して い る4,5)。 この 方 法 で は 使 用 す るア イ ソ トー プ の 量 が きわ め て 少 量 で す み,し か も比 活性 の 高 い標 識 少糖 ア ル コー ル を得 る こ とが で き る。 Asn-型 糖 鎖 の 遊 離 に は 当初 糖 鎖 の 根 本 に 共 通 に 存 在 す るN,N'-ジ ア セ チ ル キ ト ビ オ ー ス 部 分 を 水 解 す る 様 々 な エ ン ドーβ-N-ア セ チ ル グ ル コ サ ミニ ダ ー ゼ 群 が 発 見 精 製 され6∼8),糖鎖 の 構 造 研 究 に広 く使 われ て き た。 しか しこ れ ら の酵 素 群 に は図3に 示 す よ う な厳 密 な基 質 特 異 性 が存 在 す る た め に,そ の 応 用 範 囲 が 限 定 さ れ て い た 。 この 阻 路 を打 開 す る た め に 全 て のAsn-型 糖 鎖 を定 量 的 に 少 糖 と して 遊 離 す る 方 法 を 求 め て 研 究 が 進 め ら れ,ヒ ド ラ ジ ン 分 解 法9)が 開 発 さ れ た。 こ の 化 学 的 Asn一型 糖 鎖 の 遊 離 法 は,精 製 した 糖 タ ンパ ク 質 ば か り で な く,サ ンプ ル の 脱水 方法 さ え工 夫 す れ ば細 胞 や組 織 か ら も全 て のAsn型 糖 鎖 を少 糖 と して 遊 離 す る こ とが で き るの で,細 胞 の 分 化 や 形 態 形 成 に伴 な う糖 鎖 パ タ ー ンの 動 き を調 べ る上 で きわ め て 有 効 で あ る。 そ の 後Asn-型 糖 鎖 をGlcNAc→Asn結 合 部 位 で 切 り 離 し,最 終 的 に糖 鎖 を少 糖 と して 遊 離 す るグ リ コペ プ チ ダ ー ゼ(N-glycanase)10,11)が 様 々 な生 体 材 料 か ら発 見 さ れ,そ の一 部 は市 販 され る よ うに な った 。 これ らの 酵 素 は後 述 す るAsn-型 糖 鎖 の サ ブ グ ル ー プ に関 係 な く これ を遊 離 し,し か も タ ンパ ク質 部 分 を破 壊 しない の で 糖 鎖 の機 能 解 析 の上 で きわ め て 有 用 で あ る。 た だ し酵 素 で あ る の で,糖 鎖 の 全 構 造 解 析 に際 して はそ の 基 質 特 異 性 や タ ンパ ク 質部 分 の立 体 障 害 を含 め た応 用 限界 につ い て の 充 分 な検 討 が不 可 欠 と思 わ れ る。 以 上 に の べ た諸 技 術 に よ っ て遊 離 さ れ た少 糖 群 は全 て N-ア セ チ ル グ ル コ サ ミン残 基 を還 元 末 端 に持 っ て い る の で,こ れ をNaB3H4で 還 元 標 識 して 定 量 的 に3H標 識 少 糖 ア ル コ ー ル に変 え る12)こと に よ り,以 後 の分 画 や構 造研 究 にお け る検 出 を容 易 に す る こ とが で きる。 ま た還 元 末 端 を蛍 光 標 識 す る手 法13)も開発 さ れ て い る。 か く して 得 られ た サ ン プル は 標 識 少糖 の 混合 物 で あ る か ら,構 造 研 究 に先 立 って まず これ を分 画 す る必 要 が あ る。 こ の 目的 の た め に は,糖 鎖 に含 まれ るシ ア ル酸 や硫 酸 基,リ ン酸 基 な ど の荷 電 を利 用 した ろ紙 電 気 泳動 法 や イ オ ン交 換 ク ロマ トグ ラ フ ィ ー,お よ び糖 鎖 の サ イ ズ や 物 性 を利 用 した ゲ ル ろ 過 法14),HPLC15)な どが 広 く使 わ れ て きた。 こ れ ら に加 え て近 年 レクチ ンや 抗 体 を使 った 親 和 性 ク ロマ トグ ラ フ ィー(affinity chromatography)が きわ め て有 効 な少 糖 分 画 手 段 と して盛 ん に使 わ れ は じめ た16)。こ の方 法 は 将 来糖 鎖 の 自動 分 析 機 開 発 の 糸 口に な 1 2 3Fig. 3 General structures of the susceptible sugar chains for endo-ƒÀ-N-acetylglucosaminidases.
1. Endo-ƒÀ-N-acetylglucosaminidase D ; 2. endo-ƒÀ-N-acetylglucosaminidase H ; 3. endo-ƒÀ-N-acetylglucosaminidase C11. R represents either hydrogen or sugars. Arrows indicate the positions of hydrolysis.
454 有 機 合 成 化 学 第50巻 第5号 (1992)
( 78 )
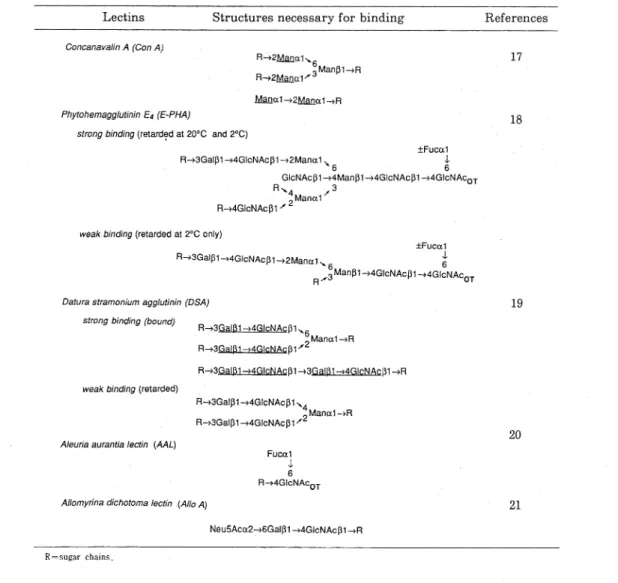
る と考 え られ るの で,少 し詳 細 に紹 介 して お く こ とに す る。 コ ンカ ナ バ リ ンA(Con A)は 図4に 示 す よ うな構 造 に 親和 性 を持 って い る 。 この構 造 は糖 タ ンパ ク質 の糖 鎖 を 構 築 す るMan α1→,GlcNAc α1→,Glc α1→ 残 基 に 含 まれ て い るが,尾 形 ら17)は不 溶 化 したConAの カ ラ ム に お け る様 々 な少 糖 お よび糖 ペ プ チ ドの挙 動 を調 べ て, ConAと 反 応 す る残 基 が 分 子 内 に一 つ しか ない もの は カ ラ ム か ら遅 れて 溶 出す るが 結 合 は しない こ と,同 残 基 を 二 つ 持 っ た も の は カ ラ ム に結 合 して100mMの α-メチ ル グ ル コ シ ドを含 む緩 衝 液 で溶 出 さ れ て くる こ と をみ い 出 し た。 こ の 発 見 が 糸 口 とな っ て,様 々 な形 の α-マン ノ シ ル 残 基 を 含 むAsn-型 糖 鎖 の 分 画 分 析 に, Con A-Sepharoseカ ラ ム を 用 い た 親和 性 ク ロマ トグ ラ フ ィ ー が 有効 な手 段 と して使 わ れ る よ うに な っ た。 ConA-Sepharoseカ ラ ム の 導 入 が 引 き金 とな って,そ の後 様 々 な不 溶 化 レク チ ンカ ラ ムが 糖 鎖 の 分 画 ・精 製 の 目的 で 開発 され て き たが,表1に そ の 結 合 特 異 性 が 明 確 に解 明 さ れ て い る もの につ いて ま とめ て お く。 少糖 群 の こ れ ら レクチ ン カ ラ ム にお け る挙 動 は,表1に 示 した構 造 要 求 性 さ え満 た され てお れ ば他 の 部 分 の 構 造 に は 殆 ん ど左 右 され ない の で,適 切 な レ クチ ン カ ラ ムの 組 み合 わ 17 18 19 20 21Fig. 4 The structure recognized by Con A.
Table 1 Binding specificities of immobilized lectin columns .
(79)
糖 タンパク質糖鎖 の構造 と機能
455 せ に よ って個 々 の 少糖 を単 一 に まで精 製 す る こ と も可 能 で あ る22)。 分 画 され た糖 鎖 の構 造 解析 に は エ キ ソグ リコ シ ダ ーゼ 群 を利 用 した 逐 次 消 化 法23)が広 く使 わ れ て い る。 エ キ ソ グ リ コ シ ダー ゼ は糖 鎖 の 非 還 元 末 端 か ら単糖 を遊 離 す る の で,標 識 に 使 わ れ た3Hや 蛍 光 原 子 団 が 最 後 まで 少 糖 側 に 残 る こ とに な る。 従 って 同 じサ ンプ ル を繰 り返 し使 用 で き る点 が きわ め て 有 利 で あ る 。 また,エ キ ソグ リコ シ ダー ゼ 群 に は様 々 な異 な るア グ リ コ ン特異 性 を持 っ た もの が 発 見 され て い る の で,こ れ を う ま く使 い わ けれ ば 遊 離 さ れ る単 糖 の結 合 位 置 を含 め て構 造 を解 明 す る こ と も可 能 とな る。 図5に 示 す 二 つ の5糖 異 性 体 を例 に と っ て 説 明 して み よ う。 この 二 つ の 少糖 は 人 乳 か ら単 離 され た もの で あ るが,通 常 の 少 糖 の 分 画 手段 で は相 互 に分 離 す る こ とがで き な い。 こ の両 者 の 混 合 物 を ア ーモ ン ドエ ム ル シ ン(Almondemulsin)か ら精 製 した α-フコ シ ダ ー ゼ1で 消 化 す る と,と も に フ コ ー ス が 外 れ た Gal β1→ 3GlcNAcβ1→3Galβ1→4GlcoT (LNT) と Gal β1→ 4GlcNAcβ1→3Galβ1→4GlcOT (LNnT)の 混 合 物 と な る 。 ア ー モ ン ド の α-フ コ シ ダ ー ゼ1はFuc α1→ 3GlcNAc結 合 とFucα1→4GlcNAc結 合 を 水 解 す る が, 他 の α-フ コ シ ル 結 合 は 切 る こ と が で き な い24)。 次 に Gal β1→4GlcNAc結 合 の み を 水 解 す る 肺 炎 双 球 菌 の β-ガ ラ ク ト シ ダ ー ゼ25)で 消 化 す る と,LNnTの み が 水 解 さ れ てGlcNAcβ1→3Galβ1→4GlcOTと な り,LNTの 方 は そ の ま ま 残 る 。 最 後 にGalβ1→3GlcNAc結 合 と Gal β1 →4GlcNAc結 合 の 両 方 を 水 解 す る ナ タ マ メ の β-ガ ラ ク ト シ ダ ー ゼ で 消 化 す る と,残 っ た4糖 が GlcNAc β1→ 3Galβ1→4GlcOTに ま で 分 解 さ れ る 。 こ の 結 果 を つ な ぎ 合 わ せ る と,図5の 二 つ の 異 性 体 の 構 造 と そ の モ ル 比 が 明 ら か に な る わ け で あ る 。 少 糖 サ ン プ ル が 比 較 的 潤 沢 に 使 え る 場 合 に は Vliegen-thartら26)に よ っ て 開 発 さ れ たNMR法 を 使 っ た 糖 鎖 の 構 造 解 析 法 が 有 効 に 応 用 で き る 。 2. 糖 タ ン パ ク 質 のAsn一 結 合 糖 鎖 構 造 に 含 ま れ る 法 則 性 と そ の 生 合 成 機 構 前 項 に の べ た 様 々 な 分 析 手 法 を 駆 使 し て,多 様 な 糖 タ ン パ ク 質 糖 鎖 の 構 造 研 究 が 進 め ら れ た 結 果,Asn-型 糖 鎖 に は ム チ ン型 に は み ら れ な い よ う な 構 造 上 の 法 則 性 が 存 在 す る こ と が 明 ら か に な っ た 。 即 ち,全 て のAsn-型 糖 鎖 に は,Manα1→6 (Manα1→3) Manβ1→ 4GlcNAc β1→4GlcNAcと い う 共 通 の 母 核 構 造 が 存 在 し,こ の 母 核 に つ い た 糖 鎖 の 構 造 の 違 い に よ っ て,図6に 示 す よ うFig. 5 Two isomeric pentasaccharides isolatod from
human milk.
456 有 機 合 成 化 学 第50巻 第5号 (1992)
(80 )
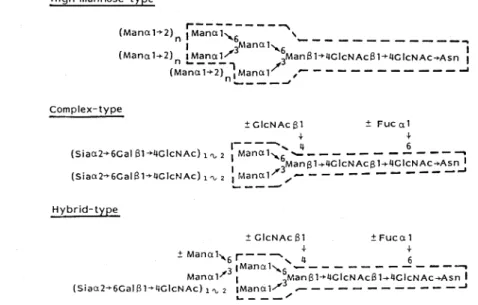
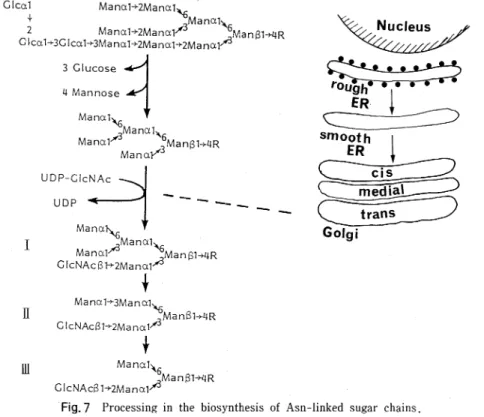
な 三 つ の サ ブ グ ル ー プ に 分 け る こ と が で き る の で あ る27)。 第1の グ ル ー プ は 母 核5糖 に α-マ ン ノ シ ル 残 基 の み が2∼6ヶ つ い た も の で,高 マ ン ノ ー ス 型 (high mannosetype)の 名 で 呼 ば れ て い る 。 こ の 群 の 糖 鎖 の 特 徴 は,Manα1→6 (Manα1→3) Manα1→6 (Man α1 → 3)Manβ1→4GlcNAcβ1→4GlcNAcと い う7糖 を 共 通 の 母 核 に 持 ち,そ の 非 還 元 末 端 に 位 置 す る 三 つ の α-マ ン ノ シ ル 残 基 に 結 合 し たManα1→2残 基 の 数 と 結 合 位 置 に よ っ て,糖 鎖 構 造 の 多 様 性 が 生 み 出 さ れ て い る 。 第 2の グ ル ー プ は 共 通5糖 母 核 以 外 に マ ン ノ ー ス を 持 た ず,母 核 の 二 つ の α一マ ン ノ シ ル 残 基 に 合 計 で 最 高5本 ま で の ガ ラ ク ト ー ス,NV一 ア セ チ ル グ ル コ サ ミ ン,フ コ ー ス,シ ア ル 酸 な ど か ら な る 側 鎖 が 結 合 し て お り,複 合 型 (complextype)の 名 で 総 称 さ れ て い る 。 第3の グ ル ー プ は 共 通 の5糖 母 核 のManα1→6腕 に は α-マ ン ン シ ル 残 基 が1∼2ヶ 結 合 し,Manα1→3腕 に は 複 合 型 と 同 様 の 側 鎖 が つ い た 丁 度 高 マ ン ノ ー ス 型 と複 合 型 の 混 成 体 の 形 を し て い る こ と か ら,混 成 型(hybridtype)の 名 で 呼 ば れ て い る 。 複 合 型 と混 成 型 の 糖 鎖 に は,糖 鎖 の 一 番 根 本 のN-ア セ チ ル グ ル コ サ ミ ン残 基 のC-6位 に つ い た α-フ コ シ ル 残 基 と5糖 母 核 の β一マ ン ノ シ ル 残 基 のC-4 位 に つ い た β-N-ア セ チ ル グ ル コ サ ミ ン 残 基(bisecting GlcNAcと 呼 ぶ)の 有 無 に よ っ て ,さ らに糖 鎖構造 の多 様 性 が 生 み 出 さ れ て い る。 Asn-型 糖 鎖 に 認 め られ る こ の 構 造 の 法 則 性 は,そ の き わ め て特 異 な 生 合 成 機 構28)に基 因 して い る。 ポ リペ プ チ ドの セ リ ン残 基 や トレ オニ ン残 基 に単 糖 が 一 つ ず つ 付 加 され て 作 り出 さ れ る ムチ ン型 糖 鎖 と は異 な り, Asn-型 糖 鎖 の 生 合 成 は ポ リ イ ソ プ レ ノ ール の 一 種 で あ る ドリ コ ー ル の ピロ リ ン酸 エ ス テ ル に結 合 したN-ア セ チ ル グ ル コサ ミン残基 に,単 糖 が 一 つ ず つ付 加 す る反 応 で 作 り 出 さ れ た14糖(Glc3・Mang・GlcNAc,)が 粗 面 小 胞 体 で 翻 訳 され つ つ あ る ポ リペ プ チ ド上 のAsn残 基 に移 し植 え られ る こ とに よ っ て 開始 さ れ る(図7)。 こ の際 に糖 鎖 の 付 くAsn残 基 は 必 ずAsn-X-Serま た はThrと い う トリ ペ プチ ド構 造 を と っ て い る。 この よ う に して 生 まれ た ば か りの タ ンパ ク 質 に結 合 したAsn-型 糖 鎖 は,次 に タ ン パ ク 質 が 滑 面 小 胞 体 ,cis-Golgiと 移行 して い く間に,3 個 の α-グル コ ース 残 基 と4個 のManα1→2残 基 が小 胞 体 の 膜 に 結 合 した2種 類 の α-グル コ シ ダ ー ゼ と1種 類 の α-マ ン ノ シ ダ ー ゼ に よ っ て水 解 除 去 され る。 タ ンパ ク質 がme4ial-Golgiに 移 る と,こ の 器官 の 膜 に 局 在 す る β-N-ア セ チ ル グ ル コサ ミ ン転 移 酵 素1(Gn-TI)が 働 い て,高 マ ン ノ ー ス型 の 母 核7糖 に1分 子 のN-ア セ チ ル I II II I
Fig. 7 Processing in the biosynthesis of Asn-linked sugar chains. R=GleNAc ƒÀ1 . 4GIcNAc (Asn-X-Ser or Thr) protein.
(81)
糖 タンパ ク質糖鎖の構 造 と機 能
457 グ ル コ サ ミンが 付 加 す る 。 こ のN-ア セ チ ル グ ル コサ ミ ンの付 加 に よ っ て,共 通5糖 母 核 のManα1→6腕 に結 合 した 二 つ の α-マン ノ シ ル 残 基 の 立 体 配 置 が 変 わ り, Golgiの 膜 に結 合 した α-マン ノ シ ダ ーゼ の働 きで 水 解 除 去 され る よ うに な る。 この 際 に残 りのManα1→6残 基 は酵 素 の働 きを受 け る こ とな く糖 鎖 上 に残 る。 以 上 の 一 連 の 反 応 がAsn-型 糖 鎖 の プ ロ セ シ ン グ と呼 ば れ る もの で あ る。 こ の プ ロ セ シ ング の 中 間体 で あ る1 とIIの 糖 鎖 に もう一 つ 別 の β-N-ア セ チ ル グ ル コサ ミ ン転 移 酵 素 皿(Gn-TImが 作 用 してbisecting GlcNAcが 付
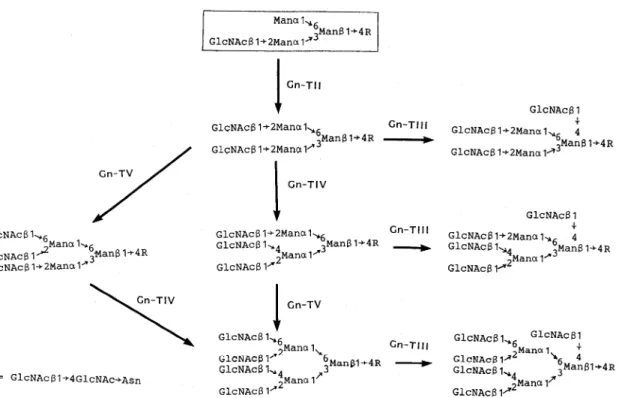
加 さ れ る と,Golgiの α-マ ン ノ シ ダー ゼ に よ る α-マ ン ノ シ ル残 基 の 除 去 が 起 こ らな くな る29)。この よ うな状 態 か ら後 で 述 べ る よ うな側 鎖 の成 熟 が起 こ って作 り出 され る一 連 の 糖 鎖 が 混 成 型糖 鎖 で あ る 。 さて,プ ロセ シ ン グの 最 終 産物 で あ る6糖(図7の 皿) は,次 に図8に 示 す よ うに様 々 な β-N-ア セ チ ル グル コ サ ミ ン転 移酵 素 群 に よ っ て色 々 なN-ア セ チ ル グ ル コサ ミ ン残 基 が付 加 され て,2本 鎖 か ら5本 鎖 まで の 複 合 型 糖 鎖 の 原 型 が 作 り出 さ れ る 。 一 方,根 本 のN- ア セ チ ル グ ル コ サ ミ ン残 基 のC-6位 に 結 合 す る フ コ ー ス が 付 加 す る た め に は,Gn-TIの 働 きで 作 り出 され る GlcNAc β1→2Manα1→3グ ル ー プ の 存 在 が 必 須 で あ り29),こ の た め に高 マ ン ノー ス 型糖 鎖群 に は こ の フ コー ス残 基 が 一 切 結 合 して い な い。 図8の 反 応 で 共 通5糖 母 核 に付 加 され た 一 連 の β- N-アセ チ ル グル コサ ミ ン残 基 に は,次 に 図9に 示 す よ うな 一 連 の 反 応 で 単 糖 が 逐 次 付 加 さ れ て 側 鎖 の成 熟 が 起 こ る。 この 際 にbisecting GlcNAcに は単 糖 の付 加 が 一 切 起 こ ら ない 。 この糖 鎖 の 成 熟 に関 与 す る糖 転 移 酵 素 群 は, 恐 ら くムチ ン型 の 糖 鎖 や糖 脂 質 の糖 鎖 の伸 長 に も関 与 し てい る もの と思 わ れ る。 これ らの 多様 な糖 転 移 酵 素 群 の 協 同 作 用 で 多 様 な 側 鎖 構 造 が 作 り出 され る わ け で あ る が,こ れ を図8に 示 した 枝 分 れ と組 み 合 わ せ る と実 に 多 様 な複 合 型 糖 鎖 が 生 み 出 され る こ とに な る。 生 体 内 で 演 じられ て い る識 別 現 象 に は,こ の 多様 な 複 合 型 糖 鎖 が シ グ ナ ル役 を担 って い る と考 えて よか ろ う。
3. Asn-型 糖 鎖 の 臓 器 並 び に種 特 異 性
前 項 で の べ た よ う に,複 合 型糖 鎖 の 側 鎖 を中 心 と した 可 変 部 位 の 構 造 は,糖 タ ンパ ク質 を作 り出 す 細 胞 に発 現 して い る 一 連 の糖 転 移 酵 素 の セ ッ トに よ っ て 決 定 され る。 こ の糖 転 移 酵 素 群 の セ ッ トは 分化 した 細 胞 で 異 な るFig. 8 Maturation of the Asn-linked sugar chains : formation of branching structures of complex
458 有 機 合 成 化 学 第50巻 第5号 (1992)
( 82 )
た め に,糖 タ ンパ ク 質 の糖 鎖 構 造 は臓 器 に よ って 異 な っ て い る。 ま た,同 一 臓 器 で も動 物 の種 に よ って 糖 転 移 酵 素 の発 現 パ タ ー ンが 異 な る た め に,糖 タ ンパ ク質 の糖 鎖 に は種 特 異 性 が み い 出 さ れて い る。 この 辺 の 事 情 を最 も 良 く研 究 さ れ て い る形 質 膜 結 合 酵 素 の 一 つ で あ る γ-グ ル タ ミル トラ ンス ペ プチ ダ ーゼ(γ-GTP)を 例 に とっ て 説 明 して み よ う。 図10に 様 々 な哺 乳 動 物 の 肝 臓 な らび に 腎臓 か ら精 製 さ れ た γ-GTPの 主 要 な糖 鎖 構 造 が示 し て あ る。 先 ず 腎 臓 の糖 鎖 を比 較 して み る と,ラ ッ トとウ シ の γ-GTPは 非 常 に よ く似 た 糖 鎖 を持 って い る が,マ ウ ス や ヒ トの も の はそ れぞ れ に異 な って い る。 一 方,肝 臓 の γ-GTPで み る と,ラ ッ トと ヒ トの 酵 素 は 良 く似 た 糖 鎖 を持 つ が,マ ウ ス の もの は大 き く異 な って い る 。 こ の よ うに γ-GTPの 糖 鎖 に は 腎臓,肝 臓 と も に顕 著 な動 物 種 差 が 存 在 す るの で あ る。 次 に マ ウ ス の γ-GTPの 糖 鎖 を 腎臓 と肝臓 で 比 較 して み る と,と も に2本 鎖 複 合 型 で は あ るが,前 者 の もの の 側 鎖 が フ コ ース を持 って シア ル 酸 を持 た な い の に対 し, 後 者 の 方 は逆 に シ アル 酸 を多 量 に 含 ん で お り,フ コ ー ス は持 っ て い な い 。 似 た よ うな違 い が ヒ トの γ-GTPの 糖 鎖 に もみ い 出 され る。 こ の よ うに γ-GTPの 糖 鎖 に は明 白 な臓 器 差 が存 在 して い るの で あ る 。糖 タ ンパ ク質 糖 鎖 の種 差 や臓 器 差 の存 在 は そ の後 リボ ヌ ク レア ーゼ で もみ い 出 され てお り36),糖 タ ンパ ク質 に普 遍 的 に み い 出 さ れ る 現 象 で あ ろ う と考 え られ て い る 。 と ころ で 図10の 糖 鎖 群 をみ る と,腎 臓 の γ-GTPの 糖 鎖 で は どの 哺 乳 類 で も共 通 にbisectingGlcNAcを 持 っ て い る の に対 し,肝 臓 の 酵 素 で は この 残 基 が 一 切 含 まれ て い な い こ とが わ か る。 実 は γ-GTPば か りで な く,我 々 の これ まで に調 べ た 様 々 な動 物 の 肝臓 で 作 られ る糖 タ ンパ ク質 の糖 鎖 に は bisectingGlcNAcを 持 っ た もの が 一 切 み い 出 さ れ て い な いの で あ る。 こ う した事 実 か ら哺 乳 動 物 の 肝 細 胞 は そ の 分 化 の 過 程 でGn-T皿 の 発 現 が完 全 に抑 え られ て い る も の と考 え られ る 。 これ に対 して 腎臓 を構 築 す る細 胞 で は こ の酵 素 が哺 乳 類 に 共 通 に発 現 さ れ て い る と い え よ う。4. Asn-型 糖 鎖 の部 位 特 異 的 な構 築
さ て これ まで に の べ て きた よ うに,糖 タ ンパ ク質 の 糖 鎖 の構 造 は こ れ を産 生 す る細 胞 のGolgiに 具 わ った 糖 転 移 酵 素 群 の組 み 合 わ せ に よ って 決 定 さ れ る わ け で あ る が,同 一 の 細胞 が作 り出 す異 な った糖 タ ンパ ク質 が 違 っ た糖 鎖 を持 つ 例 も数 多 くみ い 出 され て い る。例 え ば血 清 中の トラ ンス フ ェ リ ン とセ ル ロ プ ラス ミンは と もに 肝 実 質 細 胞 に よ って作 り出 され る が,ト ラ ンス フ ェ リ ンの 糖 鎖 は フ コ ー ス を持 た な い2本 鎖 複 合 型 で あ る の に 対 し27),セ ル ロ プ ラス ミン に は3本 鎖 複 合 型 が 含 まれ て い る38)。従 って糖 鎖 の 成 熟 の段 階 で は,ポ リペ プ チ ド部 分 の 構 造 が何 らか の コ ン トロ ー ル の役 割 を果 して い る もの と思 わ れ る 。 こ の 問題 に 関連 して重 要 な知 見 が タ ンパ ク 質 の 部位 特 異 的 グ リコ シ レー シ ョ ンの存 在 で あ る 。例 え ば ヒ ト絨 毛 性 性 腺 刺 戟 ホ ル モ ン(hCG)は α,β二 つ の サ ブユ ニ ッ トか ら構築 さ れ てお り,両 サ ブユ ニ ッ トと もに 2本 ず つ のAsn-型 糖 鎖 が 含 ま れ て い るが,実 は この 2 本 の糖 鎖 の 構 造 が そ れ ぞ れ に異 な っ て い る の で あ る。 αFig. 9 Formation of various outer chain moieties
R represents the core pentasaccharide linked to a
( 83 )
糖 タンパ ク質糖鎖の構造 と機能
459 サ ブ ユ ニ ッ トに は 表2に 示 す よ うに シ ア ル酸 を持 った フ コー ス を含 ま な い1本 鎖 と2本 鎖 複 合 型 糖 鎖 が ほ ぼ1: 1の 比 率 で 含 ま れ て い る。 これ に対 し,β サ ブ ユ ニ ッ ト に は シ ア ル 酸 を持 っ た フ コ ー ス を含 む2本 鎖 と,含 まな い2本 鎖 が1:1の 比 率 で 含 まれ て い る39)。一 つ の糖 タ ンパ ク質 分 子 の 二 つ の 位 置 に 異 な っ たAsn-型 糖 鎖 が つ い て い る とい う事 実 は,糖 鎖 の 成 熟 に タ ンパ ク質 部 分 が 何 らか の 関与 を して い る こ と を よ り強 く示 す もの とい え る 。 最 近 我 々 は 遊 離 αサ ブ ユ ニ ッ トの 糖 鎖 の 構 造 研 究 を進 め て,こ の 問題 を説 明 す る有 力 な仮 説 を提 出 した40) 。 妊 婦 尿 中 に はhCGの1/100程 度 と微 量 で は あ る が 遊 離 の αサ ブ ユ ニ ッ トが 含 まれ て い る。 こ の サ ブ ユ ニ ッ ト は,hCGを 構 成 して い る αサ ブユ ニ ッ ト と同 じ ア ミ ノ 酸 配 列 を 持 つ が,な ぜ か β サ ブユ ニ ッ ト と会 合 す る こ とがで きな い。 我 々 は この 興 味 あ る現 象 の 分 子 的 背 景 を 明 らか に す る た め に,遊 離 αサ ブ ユ ニ ッ トの 糖 鎖 を詳 し く調 べ て,こ の サ ブユ ニ ッ トに はAsn-型 糖 鎖 が1本 しか 含 まれ て い な い こ と,そ の 構 造 は表2の β の 糖 鎖 と同 じで1本 鎖 複 合 型 は一 切 存 在 しな い こ とを明 らか に し た41)。この 知 見 を も とに,我 々 はhCGのAsn一 型 糖 鎖 の 生 合 成 機 構 を図11の よ う に 考 えて い る 。 即 ち胎 盤 の 絨 毛 細 胞 の 粗 面 小 胞 体 の 中 で 作 り出 さ れ る α と β の 二 つ の サ ブ ユ ニ ッ トに は,そ の翻 訳 の過 程 で ドリコ ー ル 中 間体 か ら2本 のGlc3・MaI19・GlcNAc,と い う14糖 が 付 加Fig. 10 Major sugar chain structures of y-GTPs from the
460 有機 合 成 化 学 第50巻 第5号 (1992)
( 84 )
さ れ 直 ち にMan9・GlcNAc,に 変 換 さ れ る が,α サ ブ ユ ニ ッ トの ご く一 部 に は糖 鎖 が1本 つ き損 う。 この 糖 鎖 の 不 完 全 な付 加 は,ポ リペ プ チ ドの折 りた たみ が 糖 鎖 の 付 加 部 分 で あ るAsn-X-Thrの 一 つ を隠 して しま うた め に 起 こ る ので あ ろ う。 この よ うな 糖 鎖 付 加 の 阻 害 機 構 は, 2ヶ 所 のAsn-X-Thr(Ser)を 持 つ 卵 白 ア ル ブ ミ ン糖 鎖 が 1本 しか つ い て い な い 例 をは じめ と して,多 くの 糖 タ ン パ ク質 で み られ る糖 鎖 の 欠損 現 象 を説 明 す る た め に 広 く 受 け 入 れ られ て い る 。 糖 鎖 が2本 つ い た αサ ブユ ニ ッ トは 直 ち に β サ ブユ ニ ッ トと小 胞 体 内で 会 合 し, medial Golgiに 到 達 す る まで に図7に 示 した 糖 鎖 皿 に ま で プ ロ セ シ ン グ を受 け る が,そ の後 のGolgiで の糖 鎖 の成 熟 に 際 して は,お そ ら く会 合 したサ ブユ ニ ッ ト同 志 の 立体 的 影 響 の た め に αサ ブユ ニ ッ トの 糖 鎖 の 一 つ は1本 鎖 の ま まで 止 ま り,も う一 つ は2本 鎖 に まで 成 長 す る が,と もに フ コ ー ス が付 加 され ない 。 一 方,β サ ブ ユ ニ ッ トの 糖 鎖 は2本 鎖 に まで 成 長 す るが そ の1本 に は αサ ブユ ニ ッ トの 影 響 で フ コ ー ス の 付 加 が 起 こ らな い の で あ ろ う。 糖 鎖 が1本 しか 付 か な か った αサ ブユ ニ ッ トは β と会 合 で きず,Golgi内 で は β の 影 響 を受 け な い た め に, 糖 鎖 の 成 熟 は絨 毛 細 胞 の 持 つ 糖 鎖 合 成 能 力 が最 大 限 に発 揮 され て,フ コ ース を持 った2本 鎖 に まで成 長 す る の で あ ろ う。 5. Asn-型 糖 鎖 の 機 能 的 役 割 以 上 に のべ て きた よ うに糖 タ ンパ ク 質糖 鎖 の 形 成 機 構 の特 徴 が 解 明 され,か つ そ の構 造 が 的確 に解 析 で き る よ うに な る と,こ れ まで漠 然 と して い た糖 鎖 の 機 能 的 な 働 きが 分 子 レベ ル で 明 らか にな りは じめ た 。 現 在 まで に集 積 して き た知 見 を整 理 して み る と,ム チ ン型 糖 鎖 はポ リ ペ プ チ ド上 の 局所 に 親 水性 の グ ル ー プ を供 与 す る とい う 物 性 賦 与 の 面 で 重 要 な 役 割 を演 じて い る よ う に思 わ れ る 。 これ に対 して生 体 で営 ま れ て い る様 々 な認 識 現 象 に お い て は,Asn-型 糖 鎖 が シ グ ナ ル の 役 割 を担 っ て い る こ と を示 す事 実 が 次 々 と明 ら か に な りは じめ て い る。 本 項 で は紙 数 の 関係 もあ り,そ の 中か ら代 表 的 な 話 題 を二 つ 選 んで 紹 介 す る こ と に しよ う。 5.1. 免 疫 にお け る糖 鎖 の 働 き 近 年 の 免 疫 学 の進 歩 に よ り,細 胞 性 免 疫 反 応 と液 性 免 疫 反 応 が 相 互 に複 雑 な 免 疫 担 当 細 胞 相 互 作 用 の ネ ッ トワー クに よ って調 節 され て い る こ とが 明 らか に な って きた 。 しか も これ らの相 互 作 用 に糖 鎖 が 関 与 す る こ とが 次 々 に 報 告 され て お り, 糖 鎖 の 機 能 を考 えて い く上 で 恰 好 の 話 題 で あ る の で まず こ の 問 題 を と り上 げ る こ と にす る。 閉 鎖 され た 循 環 系 に は様 々 な血 液細 胞 が含 ま れ て い る が,そ の 中 で 免 疫担 当 細胞 だ け は 毛細 血 管 の 内皮 細 胞 を す り抜 け る 形 で 系 の外 側 に 出 る こ とが で き る。 Gesner とGinsburgは リ ンパ 球 が 内 皮 細 胞 を通 り抜 け る に 際 し て,そ の 形 質膜 糖 タ ンパ ク 質 の糖 鎖 がパ ス ポ ー トの 役 を 果 して い る可 能 性 を示 した42)。そ の 後,内 皮 細 胞 表 面 にELAM-1(Endothelial-Lymphocyte Adhesion Molecule)の
名 で 呼 ば れ る リ ンパ 球 接 着 分 子 が 発 見 され43),こ の 分 子 が リ ンパ 球 表 面 のNeu5Acα2→3Galβ1→3ま た は 4 (Fucα1→4ま た は3)GlcNAcと い う二 つ の 異 性 体4 糖 構 造 を認 識 す る レ クチ ンで あ る こ とが 判 明 した44∼46) 。 免 疫 担 当細 胞 間相 互 作 用 にお い て 主 要 な役 割 を果 して い る の は,染 色 体 のMHC(Major Histocompatibility Complexの 略)領 域 に含 ま れ る 遺 伝 子 に よっ て 作 り出 さ れ る形 質 膜 糖 タ ンパ ク質 群 で あ る。 同 種 異型 動 物 か ら採 取 した リ ンパ 球 を混 合 す る とT細 胞 の 増 殖 が 引 き起 こ
462 有 機 合 成 化 学 第50巻 第5号 (1992)
( 86 )
さ れ る 現 象 は,混 合 リ ンパ 球 反 応(Mixed Lymphocyte Reaction,略 してMLR)と 呼 ば れ,免 疫 担 当細 胞 間 相 互 作 用 を研 究 す る一 つ の モ デ ル と して 使 わ れ て い る。Hart 47 ) はMLRに 用 い る 同種 異 型 刺 戟 細 胞 を前 も って ツ ニ カ マ イ シ ン で 処 理 す る とT細 胞 の 増 殖 が 起 こ ら な くな る こ と を報 告 した。 ツ ニ カ マ イ シ ン は タ ンパ ク質 へ の Asn -型 糖 鎖 の付 加 を特 異 的 に抑 え るの で,こ の 結 果 は 刺戟 細 胞 の 形 質 膜 糖 タ ンパ ク 質 のAsn-型 糖 鎖 が リンパ 球 間相 互 作 用 の シ グナ ルの 役 を果 して い る こ と を示 す もの で あ る。 免疫 担 当細 胞 の相 互 作 用 に は細 胞 表 面 同志 の 情 報 伝 達 に 加 え て,多 様 な分 泌 糖 タ ンパ ク 質 が 液 性 メ デ ィエ ー ター と して 働 い て い る こ とが 判 明 して い る 。 こ れ ら メ デ ィエ ー タ ーの 糖 鎖 が その 機 能 発 現 の 上 で 何 らか の役 割 を 果 して い る こ と を示 す 事 実 は 未 だ な い が,逆 に IL-1 やIL-2が 特 定 の糖 鎖 と結 合 す る レク チ ン活 性 を持 つ こ とが判 明 し,改 め て 免 疫 担 当 細 胞 表 面 の 糖 鎖 の 重 要 性 が 浮 上 しは じめ て い る48)。 液 性 免 疫 の 主 役 を演 じて い るの はIgGで あ るが,こ の 糖 タ ンパ ク質 の 糖 鎖 か ら ガ ラ ク トー ス を酵 素 的 に除 去 す る と,そ のFc領 域 の 機 能 が 低 下 す る こ とが 判 明 した 49 ) 。 免 疫 グ ロ ブ リ ン は全 て 糖 タ ンパ ク質 で あ り,今 後 他 の サ ブ ク ラ スの 免 疫 グ ロブ リ ン糖 鎖 の 機 能研 究 に よ って, 液 性 免 疫 にお け る糖 鎖 機 能 の 全 貌 が 明 らか に され る 日 も近 いで あ ろ う。 5.2. 糖 タ ン パ ク 質 ホ ル モ ン 糖 鎖 の 機 能 1980年 代 の 半 ば,欧 米 の い くつ か の研 究 室 か らhCGの 糖 鎖 を化 学 的 あ るい は 酵 素 的 に 除 去 す る と,標 的細 胞 へ の 結 合 は む しろ上 昇 す る が ホ ル モ ン活性 が完 全 に 消 失 す る こ とが 報 告 さ れ た 。 また,CalvoとRyan50)はhCGの αサ ブ ユ ニ ッ トを プロ ナ ー ゼ 消化 して得 た糖 ペ プ チ ドを 共存 させ る とhCGの ホ ル モ ン活 性 が 阻 害 さ れ る の に,β サ ブ ユ ニ ッ トか ら得 た糖 ペ プ チ ドで は こ の よ うな 阻害 が起 こ ら ない こ と を報 告 した。 一 方 天 野 ら51)は絨 毛性 疾 患 患 者 の 尿 か ら精 製 したhCGの ホ ル モ ン活 性 を健 常 妊 婦 の もの と比 較 分 析 して,Asn-型 糖 鎖 の構 造 変 化 が 大 きい もの 程 そ の 活性 が低 下 して い る こ とを報 告 して い る 。 こ れ ら の 知 見 はhCGが 標 的細 胞 に 作 用 す る 際 に,ホ ル モ ンの ペ プ チ ド部分 が細 胞 表 面 の レセ プ タ ー と結 合 す る と と も に,α サ ブ ユ ニ ッ トの糖 鎖 が 標 的 細 胞 形 質 膜 に含 ま れ る レ クチ ン と結 合 す る こ とがhCGの ホ ル モ ン活 性 発 現 に 必 須 で あ る こ と を示 す もの で あ る 。 近 年hCGと 構造 的 に酷 似 して い る こ とで 知 られ る脳 下 垂 体 前 葉 の 作 り出す 3つ の ホ ル モ ンLH,FSH,TSHの 糖 鎖 が 個 々 に 異 な っ て お り,ま た,hCGと も異 な る こ とが 明 ら か に され た。 従 っ て今 後 こ れ ら の ホ ルモ ンの 作 用 機 構 は糖 鎖 を抜 きに して は考 え られ な い と こ ろ まで きて い る。お わ り に
糖 鎖 に含 まれ る様 々 な生 物 情 報 を解 明 し,こ れ を生 物 学 の 知識 と して取 り入 れ て い こ う と い う考 え の も とに, 糖 鎖 生物 学 とい う新 しい学 問領 域 が 生 まれつ つ あ る 。糖 タ ンパ ク質糖 鎖 の構 造 と機 能 の問 題 は こ の糖 鎖 生 物 学 研 究 の重 要 な柱 の一 つ と な る もの で,現 在 ま で に実 に 多様 な 局面 で研 究 が 進 め ら れ てお り,本 稿 で は そ の ほ ん の 一 部 を紹 介 した に す ぎな い。 リ ンパ 球 と血 管 内皮 細 胞 の例 にみ る よ うな細 胞 と細 胞 の認 識 機構 は,受 精,細 胞 の 分 化 あ るい は 組織 形 成 とい っ た多 細 胞 生 物 で演 じ られ て い る基 本 的 な 生物 現 象 の基 盤 を なす もの で,こ れ らの現 象 に糖 鎖 が か らん で い る こ と は今 や 疑 い の な い事 実 と し て,そ の 実 態 の解 明 が待 た れ て い る 。 そ の 生 合 成 が鋳 型 に よっ て コ ン トロ ー ル され て い な い 糖 鎖 の構 造 は,当 然 の こ とな が ら タ ンパ ク 質 に く らべ て フ ァジ ーで あ り,細 胞 の 生 理 的 な状 況 の影 響 を受 け や す い。 こ う して ひ き起 こ され た 変化 が様 々 な 疾患 の 背 景 に あ る こ と は,糖 鎖 の癌 性 変化 と癌 の 悪性 度 の 関係 や慢 性 関 節 リ ウ マ チ 患 者 のIgGに み い 出 さ れ た 糖 鎖 の ガ ラ ク トー ス欠 損 現 象 等 で 明 白 に示 され て い る 。 一 方,糖 鎖 生 物 学 の 研 究 か ら得 られ る様 々 な知 見 を現 在 の バ イ オテ ク ノ ロ ジ ーの 分 野 で積 極 的 に応 用 して い こ う とい う考 えか ら,糖 鎖 工 学 とい う新 語 まで生 まれつ つ あ る。 この 小 稿 が 読 者 諸 氏 に この新 しい糖 質化 学 の潮 流 を考 え るい さ さか の 糸 口 と して お 役 に 立 て れ ば幸 い で あ る。 (平 成4年1月 14日 受 理)文
献
1) E. F. Neufeld, G. Ashwell
, "The Biochemistry of
Glycoproteins
& Proteoglycans"
, ed, by W. J.
Len-narz , Plenum Press, Inc. , New York
, 1980, p.241
2) D. M. Carlson, J. Biol
. Chem
. , 243, 616 (1968)
3) J. Amano, A. Kobata, Methods
Enzymol
. , 179
, 261
(1989)
4) J. Amano, R. Nishimura, M. Mochizuki, A.
Kobata, J. Biol
. Chem
. , 263, 1157
(1988)
5) J. Amano
, P. Staehl
, E. G. Berger , N. Kochibe
, A.
Kobata, ibid . , 266, 11461 (1991)
6) N. Koide, T. Muramatsu, ibid . , 249, 4897 (1974)
7) A. L. Tarentino, F. Maley, ibid
. , 249, 811 (1974)
8) S. Ito
, T . Muramatsu, A. Kobata, Arch
, Biochem
.
Biophys
. , 171
, 78 (1975)
( 87 )
糖 タ ンパ ク質糖 鎖 の構 造 と機 能 463Enzymol
. , 83, 263 (1982)
10) N. Takahashi, Biochem
. Biophys
. Res
. Commun
. ,
76, 1194 (1977)
11) T. H. Plummer, Jr. , J.H. Elder, S. Alexander,
A. W. Phelan, A.L. Tarentino, J. Biol
. Chem
. ,
259, 10700
(1984)
12) S. Takasaki, A. Kobata, Methods
Enzymol
. , 50, 50
(1978)
13) S. Yamamoto, S. Hase, S. Fukuda, O. Sano, T.
Ikenaka, J. Biochem
. , 105
, 547 (1989)
14) K. Yamashita, T. Mizuochi, A. Kobata, Methods
Enzymol
. , 83, 105 (1982)
15) 遠 藤 玉 夫,新 生 化 学 実 験 講 座3,糖 質I,糖 タ ン パ ク 質(上),東 京 化 学 同 人,東 京, 1989, p. 163 16) 木 幡 陽,新 生 化 学 実 験 講 座3,糖 質I,糖 タ ン
パ ク 質(上),東 京 化 学 同 人,東 京,1989, p. 197