若手の会自由集会報告
モガニ属をめぐる分類学的・生態学的研究から見えてきた
沿岸岩礁域生態系の多様性
Taxonomic and ecological revision of the genus Pugettia suggesting cryptic diversity of
rocky reef ecosystem (symposium abstract)
大土直哉
Naoya Ohtsuchi
はじめに 現在900種を超えるクモガニ類(クモガニ上科 Majoidea)のうち,底生生活期の一定期間に海藻群 落や海草群落を主要な生息場とする種は,「ケルプ クラブ(kelp crab)」と呼ばれる.我が国において ケルプクラブに相当する語は「モガニ(藻蟹)」で ある.どちらも慣習的に使用されてきた「総称」 で,特定の分類群に対応するものではないが,近年 ではモガニ科Epialtidaeモガニ亜科Epialtinaeの一部 の種に対して使われることが多いようである. ケルプクラブ類やモガニ類のなかで最も研究され ているのが,筆者が研究対象とするモガニ属Pugettia Dana, 1851である.本属には,これまでに北太平洋 の 東 西 両 岸 お よ び タ ス マ ニ ア か ら22種が知られ (Wicksten & Stachowicz, 2013; Ohtsuchi et al., 2014; Lee et al., 2017; Ohtsuchi & Kawamura, in press),そのうち 14種が日本を含む西岸に,7種がアラスカからバ ハ・カリフォルニアまでの東岸に分布する.両方の 海域にまたがって分布する種は知られていない.北 東アジア(日本,台湾,中国北部,朝鮮半島,ロシ ア極東域)の沿岸域は,12種のモガニ属が分布する, 特に種多様性の高い海域である.本稿では以下,北 太平洋東岸の浅海性種を「ケルプクラブ類」,西岸 の浅海性種を「モガニ類」と呼称する. 北太平洋東岸に分布するモガニ属7種のうちケル プクラブ類は4種であり,それらのすべてについて 何らかの生態学的研究の事例がある(Hines, 1982; Aris et al., 1983; Hultgren & Stachowicz, 2008; Daly & Konar, 2010など).ケルプクラブ類は,アラスカ沿 岸やモントレー湾,ボデガ湾において,ウニ類や ラッコなどとともに岩礁藻場生態系(特に Macro-cystisやNereocystisなどのジャイアントケルプから 成る海中林生態系)の重要な構成種として,遅くと も1960年代には生態学的知見が集積され始めた. 食物網のなかでの位置も把握されており,藻・草体 や種子,小型巻貝類を摂食し (Holbrook et al., 2000; Dobknowski, 2017), 大型魚類やタコ類,ラッコなど に捕食される(Hines, 1982など). 一方,北太平洋西岸に分布する本属15種のうち, 11種が北東アジア域に限産し,そのうちモガニ類は5 種である.これらの種もまた,岩礁藻場生態系の重 要構成種と推察されるが,生態に関する研究事例は 少なく,その対象はヨツハモガニP. quadridens (De Haan, 1837) に限られる.また,モガニ類は様々な海 藻群落に生息するとされるが (Sakai, 1976),群落の主 要構成種について言及されることは稀で,彼らがケ ルプクラブ類のように特定の海藻群落に生息するか は明らかではない.ヨツハモガニの食性に関する知 見は断片的で,しかも東北地方・北海道とその他の 東京大学大気海洋研究所国際沿岸海洋研究センター 〒028–1102 岩手県上閉伊郡大槌町赤浜1–19–8 International Coastal Research Center, Atmosphere andOcean Research Center, The University of Tokyo, 1–19–8 Akahama, Otsuchi, Iwate 028–1102, Japan
地域とでは異なる.東北地方・北海道では,本種が 稚ウニや稚アワビ,ヤドカリ類などを捕食すること が室内実験によって確認されている(白石,1997; 干 川,2003; 松尾ら,2015など).一方,その他の地域 では,ヨツハモガニが植食性であることが示唆され ている.例えば,紀伊白浜番所崎の潮間帯で行なわ れた研究では,本種の消化管内容物がホンダワラ科 褐 藻 に 大 き く 偏 る こ と が 示 さ れ た (Sato & Wada, 2000). 不思議なことに,これらの研究は互いに参照 されず,筆者の知る限り,ヨツハモガニを雑食性と 明言した文献も,海域間での食性の不一致に疑義を 呈した文献もない.ヨツハモガニの捕食者について は,おそらく大型魚類やタコ類などと推察されるが, 具体的な研究事例はない. このように,北太平洋の東岸と西岸とでは,分布 するモガニ属浅海性種の数は同じでも,それらの生 態についての理解度には大きな差がある.ケルプク ラブ類は沿岸海洋生態系の教科書にも登場するが (Schiel & Foster, 2015; Carr & Reed, 2016),北東アジ ア沿岸の藻場生態系はこれまで「モガニ類抜き」で 説明されてきた.北東アジア域において,これほど までにモガニ類の研究が進展しなかった背景には 様々な事情があるが,ヨツハモガニ1種しか生態学 的研究の対象とならなかったのは,ヨツハモガニと 近縁種の正確な判別が困難であったからと言って良 いだろう.本稿では,筆者が研究を開始する以前の 「ヨツハモガニ観」を整理し,それらがどのように 訂正されたのか,分類学的研究と生態学的研究のそ れぞれについて解説する. ヨツハモガニの分類学的再検討 1)研究開始前の「ヨツハモガニ」 標準和名「ヨツハモガニ」に対応する種は,1837 年,『日本動物誌 甲殻類編』の図版にPisa (Halimus) quadridensの名で掲載され,その2年後に出版され た本編にてPisa (Menaethius) quadridensの名で記載 された.その後,亜属Menaethius A. Milne-Edwards, 1834が属に昇格したため,本種の学名はM. qua-dridensとなるが,程なくして,当時北米のみから知 られていたPugettiaに移動させられた. 19世紀末になると,複数の研究者によってP. qua-dridensに種内変異が多いことが指摘され始め,1930 年代後半には本種の同定が “matter of difficulty” と表 現されるに至った.例えばSakai (1938, 1976) は,岩 手県産や道東厚岸産の標本が,伊豆半島下田など, より南の海域に産する “typical form” の標本より大 型であり,甲背表面に分布する顆粒の明瞭さ,眼後 歯と肝域歯の癒合の程度,ハサミの形態など,多く の形態的特徴に差異があることを指摘した. 1930年代には,ヨツハモガニの亜種が相次いで 記載された.まず静岡県の御前崎からコヨツハモガ ニP. quadridens pellucens Rathbun, 1932が記載され, そ の 後, 伊 豆 半 島 下 田 よ り ヨ ツ ハ モ ド キP. qua-dridens intermedia Sakai, 1938が記載された.しばら
くしてから酒井(1986)は,これらの亜種を二名法 で表記したが,その根拠は示されなかった. ここまでに見てきたように,遅くとも1930年代 後半までにはヨツハモガニのアイデンティティに係 わる基本情報がすべて出揃った.加えて同時期に菊 池(1932)は,富山県のカニ類についてまとめた報 文の中で,P. quadridensについて「沿岸岩礁上ニ棲 息スル普通種ニテ(中略)海藻ヲ取リ中ニ蟹アレバ 大抵此ノ種ト云ヒツベシ」と記している.この時期 の研究者のあいだには,ヨツハモガニは「磯の海藻 群落に棲む普通種で,成体・幼生ともに非常に変異 が多く,特に北日本の個体は形態的特徴がやや異な り,大きい」という認識が既にあったことが窺い知 れる. その後の研究の歴史を辿ると,ヨツハモドキを種 に昇格させたGriffin & Tranter (1986) によって,ヨ ツハモガニの分類とその近縁種との判別に関する議 論は終結しているかのように見える.ところが, 「大型のヨツハモガニ」とヨツハモガニの関係,そ の他の本種の種内変異については,遂に詳細な検討 がなされることはなく,コヨツハモガニとヨツハモ ガニの関係も実質未決着のままだった.結果とし て,現在に至るまで,日中韓露の多くの研究者によ り,幼生期をも含めて様々な形態的特徴が撮影・描 画され,「ヨツハモガニ」として掲載され続けるこ とになった. 2)種内変異か,別種か 筆者らは,上述した「ヨツハモガニの様々な形
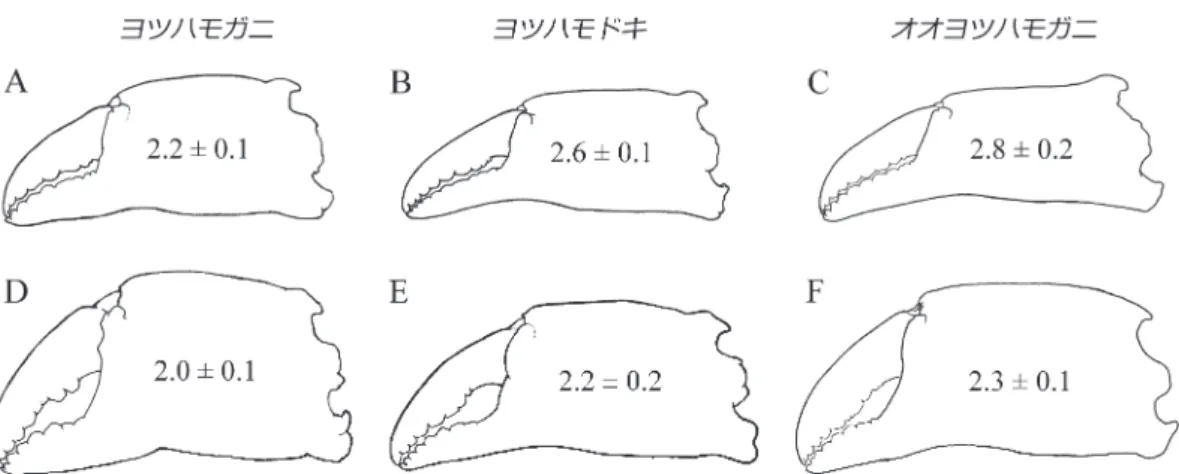
態」について,種内変異としてではなく,未記載種 が含まれている可能性も考慮して,改めて詳細に観 察する必要があると考え,まずは「大型のヨツハモ ガニ」と「typicalなヨツハモガニ」の形態的差異を 明らかにすることにした.まず,生態学的研究の調 査地であった牡鹿半島東岸と相模湾長井にて採集し た多数の標本を比較し,前者が岩手県や厚岸に産す る「大型のヨツハモガニ」 (Sakai, 1938: text fig. 28a) と,後者が下田などに産する「typicalなヨツハモガ ニ」(同:text fig. 28b) と 同 一 で あ る こ と,Sakai (1938, 1976) が指摘した部位の他にも,雄の第一腹 肢(生殖肢)を含む多くの形態的特徴に差異が認め られることを確認した. 次に,「真のヨツハモガニ」はどちらなのかを検 討した.Sakai (1938) は下田産の標本をtypicalと表 現したものの,それが「形態的特徴がタイプに一致 する」という意味なのかは判然としない.そこで Naturalis Biodiversity Center所蔵のPisa (Menaethius) quadridensのレクトタイプ標本を検討すると,ハサ ミの可動指に2, 3強歯があること,第二触角基節の 外側縁基部が眼窩後縁から隔絶されないことなどが 確認され,Sakai (1938, 1976) の「typicalなヨツハモ ガニ」こそが「真のヨツハモガニ」であることが明 らかになった. 最後に,2種類の「ヨツハモガニ」の形態的な差 異が種内変異の範囲と見なせるかについて検討し た.クモガニ類には,性的二型,二次性徴の発現, 成長に伴う形態変化,およびその他の変異など, 様々な種内変異が想定されるが,モガニ属の種同定 が難しいのは,そのような種内変異の詳細が各種に 図1. モガニ属Pugettia 3種の雄の発達段階.A–C, 未成熟個体;D–F, 最終脱皮前の成熟個体;最終脱皮後の成
おいて全く把握されていないからであった.そこ で,クモガニ類の成長様式に関する多数の研究をヒ ン ト に, モ ガ ニ 類 雌 雄 の「発 達 段 階 (ontogenetic stage)」を区別し(図1),発達段階ごとに形態的特 徴の詳細な比較を行なった.その結果,第二触角基 節以外にも,性別や成長段階の別を問わない判別形 質が見い出され,同時に,二次性徴の発現や成長に 伴う形態変化の様式が2種類の「ヨツハモガニ」間 で明確に異なることも明らかになった(一例として 図2).このような差異は「真のヨツハモガニ」の 種内変異の範囲を超えていると考えられるが,筆者 らは,念のため両者の分布域を調べることにした. 3)分布域と生息場情報の更新 まず,国内外の博物館に保管されているモガニ類 の標本調査と文献調査を行ない,国内外からの多数 の標本記録を精査した.それらによってカバーでき ない地域については,自ら採集調査を行なった.そ の結果,かつての「ヨツハモガニ」の記録の大部分 が,同属8種および他属3種のクモガニ類と誤同定 あるいは混同されたものであることが明らかにな り,その分布域は大きく変更された(図3).「真の ヨツハモガニ」は,北東アジアの沿岸域に広く分布 し,日本では南西諸島を除くほぼ全国に分布する種 とされてきたが,「真のヨツハモガニ」は,太平洋 側では福島県いわき市以北,日本海側では富山湾よ り東の海域には分布せず,朝鮮半島とロシア極東部 に も 分 布 し な い こ と が 明 ら か に な っ た. 一 方, Sakai (1938, 1976) が岩手と厚岸から報告した「大型 のヨツハモガニ」は,北東アジア域に広く分布する 図2. モガニ属Pugettia 3種の雄における,ハサミの形態と相対長 (平均値±標準偏差) の成長に伴う変化.A–
C, 最終脱皮前の成熟個体;D–F, 最終脱皮後の成熟個体.Ohtsuchi & Kawamura (in press)から再構成.
図3. モガニ属Pugettia 3種の分布域.赤い星印は,それぞれの種のタイプ産地 (ヨツハモドキP. intermediaは
下田,オオヨツハモガニは大槌湾)を示す.ヨツハモガニP. quadridensのタイプ産地 (日本) は示してい ない.Ohtsuchi & Kawamura (in press)を一部改変.
ことが明らかになった. 筆者らが種同定を確認した標本記録のみに基づい て生息場情報を整理すると,「真のヨツハモガニ」 の生息場は潮間帯から水深8 mまでの浅海域の海藻 群落に限られた.一方「大型のヨツハモガニ」は, 最深で80 m(千葉県銚子)から,また,海藻群落 に限らず,転石帯,泥底やイガイ礁など様々な沿岸 域環境から採集されており,いわきから三浦半島に かけての沿岸域においては同じ海藻群落内からヨツ ハモガニとともに採集された. 第二触角基節と第一腹肢などの重要な形質の安定 的な相違と,その他の形態的特徴の成長に伴う変化 様式の明らかな相違,加えて分布域と生息場の重複 を根拠に,両者は明らかな別種と結論し,「大型の ヨツハモガニ」を新種オオヨツハモガニとして記載 した (Ohtsuchi & Kawamura, in press).東北地方・北 海道で磯根資源の捕食者として研究されてきたのは オオヨツハモガニであり,その他の地域で植食性と 推測されていたものが「真のヨツハモガニ」であっ た. 4)亜種関係の決着 コヨツハモガニとヨツハモドキについては,それ ぞれを再記載し,標本記録を再調査した結果,ヨツ ハモガニとは,第二触角基節や雄の第一腹肢を含む 多くの形態的特徴に安定的な差異があり,分布域が 重複することが明らかになったため,両者ともヨツ ハモガニとは別種と見なすべきと結論した (Ohtsu-chi et al., 2014; Ohtsu(Ohtsu-chi & Kawamura, in press).
ちなみに,ヨツハモドキの分布域は,ヨツハモガ ニおよびオオヨツハモガニの分布域に重複するもの の,やや南方寄りであり,房総半島から台湾海峡ま でであった(図3).また,本種の生息場は他の2種 とは少し異なっていた.本種のタイプ産地は下田で あるが,小型個体を含むまとまった数の標本が得ら れているのは,大阪湾や舞鶴湾などの閉鎖的で護岸 整備された水域ばかりであったのだ.ただし,本種 の標本記録は他の2種に比べて少なく,今後も分布 域や生息場を詳細に調べる必要がある. モガニ属の生息場利用 筆者らは,相模湾長井沿岸の潮間帯から水深8 m までに存在する11種類の生息場において,底生動 物の定量採集調査を2年以上にわたって毎月行なっ た.各生息場において個体数密度に基づき優占種と 判断された十脚甲殻類種には,2種のモガニ類が含 まれた.一つはヨツハモガニであり,もう一つはモ ガニ属最小の種,アラサキモガニP. vulgaris Ohtsu-chi, Kawamura & Takeda, 2014であった.
ヨツハモガニの甲幅組成と発達段階組成の経月変 化からは,初夏~夏季にかけて水深1–4 mの紅藻群 落内に小型で未成熟の個体が急増すること,晩冬か ら初夏にかけて低潮線付近のヒジキ群落内に大型で 性的に成熟した個体が出現することが示された.本 種は,潮下帯の紅藻群落を初期生息場とし,成長に 伴ってヒジキ群落に移動すると考えられる (Ohtsu-chi et al., 2018). このような「成長に伴う生息場変化」 は,ケルプクラブ類では既に数例の報告があったが (Hines, 1982; Hultgren & Stachowicz, 2008など),モ ガニ類では初めて確認された.一方,同所的に出現 するアラサキモガニは,生活史を通じて潮下帯の有 節サンゴモ群落内やテングサ群落内に生息すること が示された (Ohtsuchi et al., 2016).相模湾長井では, ヨツハモガニが成長に伴い生息場を変えることに よって,近縁2種の同所的分布が実現していると考 えられる. 筆者らによる研究以前には,「ヨツハモガニ」の 出現動態や生活史に関連する3例の研究があった.
これらは館山坂田 (Fuseya & Watanabe, 1993),韓国 光陽湾 (Huh & An, 1998),紀伊白浜番所崎 (Sato & Wada, 2000) で行なわれた研究であるが,いずれの 海域にも複数のモガニ類が分布しているため,相模 湾長井での「真のヨツハモガニ」に基づく研究と安 易に比較することはできない.結局,筆者らは白浜 番所崎での研究のみを「真のヨツハモガニ」に関す る先行知見と見なすことにした.『動物行動の映像 データベース』(http://www.momo-p.com/) 上に,佐 藤氏が京大瀬戸臨海実験所内で撮影した動画4編が 登録されており,それらがいずれも「真のヨツハモ ガニ」を映したものと確認されたからである.
ハモガニが「冬季にのみ」低潮線付近に高密度に出 現し,その出現動態や分布様式がイソモクの季節消 長様式や分布様式とほぼ同じ傾向を示すとした.こ の研究では,ヨツハモガニが「夏季にどこにいるの か」 に つ い て は 言 及 さ れ な か っ た た め, 筆 者 は 2015年の夏季に京大瀬戸臨海実験所を訪れてシュ ノーケリングを行い,潮下帯に有力な初期生息場が ないか調査した.その結果,潮下帯に小規模のマク サ群落があり,そのなかにヨツハモガニ未成熟個体 が高密度に生息することが確認された (Ohtsuchi et al., 2018). 潮下帯の小型紅藻群落から低潮線付近の ホンダワラ科褐藻類群落への「成長に伴う生息場変 化」は,相模湾長井以外の生息地においても起こっ ているものと考えられる. 一方,同じ相模湾で長井よりやや北東に位置する 剱崎では,雌雄のすべての発達段階のヨツハモガニ が紅藻群落内から採集された.これらの磯にもヒジ キ群落と紅藻群落は生育するが,長井とは異なり岩 場の勾配が急であるため,2種類の海藻群落は水平 方向よりもむしろ垂直方向に隔てられている.その ため,晩冬から初夏の干潮時には,ヒジキ群落は完 全に干出し,低潮線付近には紅藻類が広範な群落を 形成する.このような磯では,ヨツハモガニの生息 場変化は起こらない,あるいは長井よりもかなり不 明瞭になるだろうと推察される. その他にも,筆者らはヨツハモガニの食性,藻体 選好性,マイクロハビタット選好性,デコレーティ ング・マテリアル選好性,およびそれらの成長に伴 う変化について,室内実験によって検討し,彼らの 生息場利用が,特定の海藻群落との関連から決まる のではなく,季節消長様式の異なる多様な海藻群落 から構成される「藻場全体」と様々な空間スケール で密接に関連して決定していることを明らかにしつ つある.これらの生態学的研究は,モガニ類の生息 場利用が同一種内においても,生息海域における小 型紅藻群落とホンダワラ科褐藻群落それぞれの種組 成や分布状況(特に群落間の距離)に応じて可変的 であることを示唆している.相模湾長井で明らかに なったヨツハモガニの生息場利用様式は,おそらく 本種の可変的な生息場利用のなかの顕著な一例と解 釈すべきである.本種の生息場利用については,ま だまだケーススタディを積み重ねていく必要があ る. おわりに 分類学的研究からは,北東アジア沿岸域において 多様なモガニ類が生息場や水深によって棲み分けな がら重複分布していることが,生態学的研究から は,モガニ類の生息場利用様式は種ごとに異なり, そのうえ同一種内でも生息海域における小型紅藻群 落と褐藻群落の種組成や分布様式に応じて可変的で あることが示唆された.モガニ類の種組成や生息場 利用が海域や湾によって異なるのであれば,彼らを めぐる種間関係も海域や湾によって少しずつ異な り,沿岸岩礁域生態系の構造もやはり少しずつ異な ると考えるべきだろう.モガニ類の研究は再スター トを切ったばかりである.今後,モガニ類の生態学 的研究が進展すれば,我々が漠然と想像している 「沿岸岩礁域生態系の多様性」に具体的な形が与え られるようになるかもしれない. ケルプクラブ類・モガニ類には,藻場生態系の主 要構成種として,磯根資源の生活史初期の捕食者と して,さらにはカムフラージュ戦術をもつ動物とし て,研究すべきテーマが山のようにある.筆者は, 我が国の藻場生態系の教科書に「モガニ類」のペー ジを加えるために,今後も手法にこだわらずに研究 を続けていくが,意欲ある学生の登場にも期待した い. 謝 辞 修士課程から現在に至るまで,本研究についてご 指導いただき,また本稿の草稿をご高閲いただいた 東京大学大気海洋研究所の河村知彦教授と早川淳助 教に感謝申し上げます.また本研究を遂行するにあ たって,貴重なご協力・ご助言をいただいた多くの 先生方・学生諸氏にも改めて御礼申し上げます.紙 幅の都合上,一括して御礼申し上げることをお許し いただければ幸いです. 文 献
occur-rence of Pugettia richii (Crustacea: Decapoda) on Cys-toseira osmundacea follows a diel pattern. Bulletin of Marine Science, 32: 243–249.
Carr, M. H., & Reed, D.C. 2016. Shallow Rocky reefs and kelp forests. In: H. Mooney, & E. Zavaleta (eds.), Eco-systems of California. University of California Press, Oakland, California, pp. 311–336.
Daly, B., & Konar, B., 2010. Temporal trends of two spider crabs (Brachyura, Majoidea) in nearshore kelp habitats in Alaska, U.S.A. Crustaceana, 83: 659–669.
Dobkowski, K., 2017. The role of kelp crabs as consumers in bull kelp forests—Evidence from laboratory feeding trials and field enclosures. Peer J, 5: e3372.
Fuseya, R., & Watanabe, S. 1993. Growth and reproduction of the spider crab, Pugettia quadridens quadridens (De Haan) (Brachyura: Majidae). Crustacean Research, 22: 75–81.
Fuseya, R., Yokokura, K., & Watanabe, S., 2001. Morpholo-gy and distribution of setae used for decorating on the dorsal carapace of the spider crab, Pugettia quadridens quadridens (De Haan) (Brachyura: Majidae). Crusta-cean Research, 30: 154–159.
Griffin, D. J. G., & Tranter, H. A., 1986. The Decapoda Brachyura of the Siboga Expedition. Part VIII. Majidae. Siboga Expeditie Monografie, 39(c4): 1–335.
Hines, A. H., 1982. Coexistence in a kelp forest: size, popu-lation dynamics, and resource partitioning in a guild of spider crabs (Brachyura, Majidae). Ecological Mono-graphs, 52: 179–198.
Holbrook, S. J., Reed, D. C., Hansen, K., & Blanchette, C. A., 2000. Spatial and temporal patterns of predation on seeds of the surfgrass Phyllospadix torreyi. Marine Bi-ology, 136: 739–747.
干川 裕,2003. エゾアワビ人工種苗に対するヒトデ
類3種およびヨツハモガニの捕食(室内試験).北
海道立水産試験場研究報告,64: 121–126.
Huh, S. H., & An, Y. R., 1998. Seasonal variation of crab (Crustacea: Decapoda) community in the eelgrass (Zos-tera marina) bed in Kwangyang Bay, Korea. Journal of the Korean Fisheries Society, 34: 535–544.
Hultgren, K. M., & Stachowicz, J. J., 2008. Alternative cam-ouflage strategies mediate predation risk among closely related co-occurring kelp crabs. Oecologia, 155: 519– 528.
菊池勘左エ門,1932. 富山湾,甲殻類目録(富山湾生
物調査目録V).富山教育,227: 1–23.
Lee, B-Y., Richer de Forges, B., & Ng, P. K. L., 2017. Deep-sea spider crabs of the families Epialtidae MacLeay, 1838 and Inachidae MacLeay, 1838, from the South China Sea, with descriptions of two new species (De-capoda, Brachyura, Majoidea). European Journal of Taxonomy, 358: 1–37.
松尾謙人・石原(安田)千晶・和田 哲,2015. 右鋏脚 の自切によるテナガホンヤドカリの捕食者回避行 動.Cancer, 24: 21–23.
Ohtsuchi, N., Kawamura, T., & Takeda, M., 2014. Rede-scription of a poorly known epialtid crab Pugettia pel-lucens Rathbun, 1932 (Crustacea: Decapoda: Brachy-ura: Majoidea) and description of a new species from Sagami Bay, Japan. Zootaxa, 3765: 557–570.
Ohtsuchi, N., Kawamura, T., Hayakawa, J., Kurogi, H., & Watanabe, Y., 2016. Growth patterns and population dynamics of the kelp crab Pugettia vulgaris (Decapoda, Brachyura, Epialtidae) on the coast of Sagami Bay, Ja-pan. Crustaceana, 89: 645–667.
Ohtsuchi, N., Kawamura, T., Hayakawa J., Kurogi, H., & Watanabe, Y., 2018. Ontogenetic habitat shift in Puget-tia quadridens on the coast of Sagami Bay, Japan. Fish-eries Science, 84: 211–255.
Ohtsuchi, N., & Kawamura, T., in press. Redescriptions of Pugettia quadridens (De Haan, 1837) and P. intermedia Sakai, 1938, with description of a new species. Sakai, T., 1938. Studies on the crabs of Japan III.
Brachyg-natha, Oxyrhyncha. Yokendo Co., Tokyo, pp. 193–364. Sakai, T., 1976. Crabs of Japan and the adjacent seas. Tokyo,
Kodansha Ltd., 773 pp., 251 plates.
酒井恒,1986. 珍奇なる日本産蟹類の属と種につい
て.甲殻類の研究,15: 1–4, 図版1–3, 扉絵. Sato, M., & Wada, K., 2000. Resource utilization for
deco-rating in three intertidal majid crabs (Brachyura: Maji-dae). Marine Biology, 137: 704–714.
Schiel, D. R., & Foster, M. S., 2015. The biology and ecolo-gy of giant kelp forests. University of California Press, Oakland, California, 395 pp.
白石一成,1997. 肉食動物のキタムラサキウニに対す る捕食及ぼす水温の影響.水産増殖,45: 321–235. Wicksten, W. K., & Stachowicz, J. J., 2013. Mimulus Stimp-son, 1860, a junior synonym of Pugettia Dana, 1851 (Decapoda: Brachyura: Majoidea: Epialtidae). Zootaxa, 3693: 358–364.