1.はじめに
萌芽による個体の再生は森林における樹木の更新様式の一 つであり,林冠層の更新・維持において重要な役割を果たし ている(山本,1981;Del Tredici, 2001;大久保,2002;伊藤,
2004)。また,萌芽再生能力は重要な生活史戦略の一つであ
り(Zimmerman et al.,1994;Kruger et al.,1997),高木性樹木 の生活史戦略として,萌芽特性は,伐採などの撹乱によって 生じた地上欠損部の修復と再生,栄養繁殖,および非攪乱下
における地上部の更新といった役割を果している(酒井,
1997)。
イヌブナは,主に太平洋側山地帯林に分布し,ブナ,ミズ * 連絡・別刷請求先(Corresponding Author):〒321―8505 栃木県宇都宮市峰町350 宇都宮大学農学部森林科学科:Department of Forest Science, Faculty
of Agriculture, Utsunomiya University, 350 Mine, Utsunomiya, Tochigi, 321―8505, Japan E-mail : aizawam@cc.utsunomiya-u.ac.jp
1 Department of Forest Science, Graduate School of Agriculture, Utsunomiya University 2 Department of Forest Science, Faculty of Agriculture, Utsunomiya University 3 Oji Forest and Products Co., Ltd.
4 Forestry and Forest Products Research Institute
† 現所属:三井物産フォレスト株式会社:Present Address : Mitsui Bussan Forest Co., Ltd. (2011年10月11日受付,2012年5月15日受理)
イヌブナ天然林皆伐後の針葉樹不成績造林地における
イヌブナの萌芽再生と更新木としての有用性
松本健太郎
1,†・逢 沢 峰 昭
2,*・菊 地 陽 太
3・松 本 陽 介
4・大久保達弘
2 1宇都宮大学大学院農学研究科森林科学専攻・2宇都宮大学農学部森林科学科・3王子木材緑化株式会社・ 4独立行政法人森林総合研究所 本研究は,栃木県日光市のイヌブナ天然林皆伐後に造成された針葉樹人工林のうち,激甚なクマ剥ぎ被害を受け たヒノキ人工林と植栽針葉樹が成林せず広葉樹二次林となった林分の2つの不成績造林地において,皆伐,下刈り, 除伐といった人為撹乱下におけるイヌブナの萌芽再生過程を明らかにし,イヌブナの天然更新木としての有用性を 評価することを目的として,林分構造調査および成長解析を行った。イヌブナはいずれの不成績造林地においても, 複数の萌芽幹からなる株構造を有していた。下刈りと複数回の除伐が行われたヒノキ人工林では,ヒノキが高木層 で,イヌブナが低木層で優占していた。下刈り後に除伐が行われなかった広葉樹二次林では,高木層は主として先 駆性のミズメ,ミズキおよびウリハダカエデが優占し,イヌブナは亜高木層から低木層で優占していた。広葉樹二 次林におけるイヌブナの直径成長は,周囲の先駆性の広葉樹と比べて小さいものの,いずれの萌芽幹も被圧林冠下 でゆっくりと成長していた。また,いずれの不成績造林地の亜高木・低木層においても,イヌブナに匹敵する優占 度をもつ遷移後期種はほとんどみられなかった。これらのことから,イヌブナは皆伐とその後の人為攪乱下におい ても個体群の維持が可能な萌芽再生能力を保持し,将来的にイヌブナは主要な林冠構成樹種の1つになると考えら れた。したがって,不成績造林地を針広混交林化あるいは広葉樹林化する場合には,イヌブナは天然更新木として 有用な樹種であると考えられた。 キーワード:イヌブナ,人為撹乱下,萌芽再生,針葉樹不成績造林地,広葉樹林Kentaro Matsumoto, Mineaki Aizawa, Yota Kikuchi, Yoosuke Matsumoto and Tatsuhiro Ohkubo:Sprout regen-eration of Japanese beech (Fagus japonica) and its usefulness as a regenerator in unsuccessful conifer plantations af-ter clear-cutting of natural Japanese beech forests. Japanese Journal of Forest Environment 54:81−92, 2012.
We aimed to delineate the regeneration process of Japanese beech (Fagus japonica) by sprouting under strong and fre-quent human-induced disturbances, such as clear-cutting, weeding, and salvage cutting, and to assess the usefulness of the species as a natural regenerating tree in unsuccessful conifer plantations after clear-cutting of natural Japanese beech forests in Nikko, Tochigi. We investigated the stand structures and analyzed the growth of Japanese beeches and their spatially neighboring trees, focusing on 2 types of unsuccessful conifer plantations, (1) the Hinoki cypress plantations where weed-ing and several salvage cuttweed-ings had been conducted, and which were degraded by the bark strippweed-ing by bears; and (2) sec-ondary broad-leaved forests resulting from the poor growth of conifers planted in locations where weeding had been con-ducted. Japanese beech exhibited stool structures with many stems resulting from sprouting under all the human-induced disturbances mentioned above. In the Hinoki cypress plantations, Japanese beech was the most dominant species under the tree layer of cypresses; in secondary broad-leaved forests early successional species, such as Betula grossa, were dominant in the tree layer, under which Japanese beech was dominant in the shade. Besides Japanese beech, almost no late succes-sional species was observed under the tree layer in all the types of plantations. Therefore, Japanese beech could maintain a strong ability to produce sprouts and survive in unsuccessful conifer plantations under human-induced disturbances, and might become a main tree-layer component in the future. If unsuccessful conifer plantations are converted to broad-leaved forests, Japanese beech would be the most useful natural regenerator among the surviving broad-leaved trees.
Keywords : broad-leaved forest, Fagus japonica, under human-induced disturbances, sprout regeneration, unsuccessful conifer plantation
ナラ,モミ,ツガおよびカエデ類などとともに,林冠層を構 成する樹種である(遠山1965;Sasaki,1970;藤田,1986, 1987;野崎・奥富,1990;星,1999)。イヌブナ天然林の群 落構造,株構造,萌芽によるギャップ修復過程などに関する 研究によって,イヌブナは種特性として強い萌芽再生能力を 持ち,この特性によってイヌブナ林は非大規模撹乱下におい て永続的に維持されていることが明らかにされた(Ohkubo et al.,1988, 1996;Ohkubo, 1992;大久保,2002)。また,埼玉 県秩父山地の天然林皆伐後に放置された林分における萌芽再 生過程の調査から,イヌブナが,皆伐という強度の大規模な 撹乱後,即座に萌芽を発生させ,先駆性樹種の林冠下で成長 を続け,最終的に林冠を優占する可能性が示唆された(菊地 ら,2009)。しかし,イヌブナの萌芽再生過程に関して,針 葉樹人工林内のような下刈りや除伐といった強度・複数回の 人為撹乱下における研究例はみあたらない。長谷川(1998) は,スギ造林地において,下刈りおよび除伐が,ウダイカン バの消長に及ぼす影響について研究を行い,これらの施業に よってウダイカンバの密度は著しく低下することを示した。 したがって,針葉樹人工林地における下刈りや除伐といった 撹乱は,皆伐後放置といった1回の撹乱と比べて,その後 のイヌブナの萌芽再生に及ぼす影響は極めて大きいものと予 想される。 栃木県日光市の国有林には,1970∼80年代の拡大造林期 にイヌブナ・ブナの優占した天然林を皆伐し,その後にスギ, ヒノキ,アカマツ,カラマツなどの針葉樹が植栽された造林 地が広がっている(大久保,2002)。ここでは近年,激甚なク マの樹皮剥ぎ被害が発生し,植栽木の枯死が顕在化してきて いる(丸山,2004)。また,主として除伐不足が原因と思われ るが,植栽針葉樹が成林せずに広葉樹二次林となった林分が みられる。このような激甚な獣害被害林分や広葉樹二次林で は,本来目的とした木材生産機能が著しく低下していると考 えられることから,本研究では「不成績造林地」と呼ぶこと にする。このような不成績造林地では,本研究の予備踏査の 結果,イヌブナをはじめとする広葉樹が皆伐時の伐採株から 萌芽更新している様子がしばしば観察された。近年,人工林 においても森林生態系の保全・修復や種多様性という観点か ら,多様な機能の発揮が求められている(長池,2000)。また, 戦後に奥地まで拡大した針葉樹人工林の不成績地が問題化し ている(横井・山口,1998,2000;小谷,2009)。このよう な中,針葉樹人工林を広葉樹林へと誘導する広葉樹林化の推 進が図られ始めている(林野庁http://www.rinya.maff.go.jp/j/ kikaku/plan/pdf)。したがって,広葉樹天然林の皆伐前歴を もつ獣害被害地や不成績造林地の広葉樹林化も視野に入れ, 林内に残存しているイヌブナのような広葉樹の萌芽による無 性繁殖様式を利用することで,人為的に苗木などの造林材料 を外部からもち込むことなく,天然更新が可能であるかの検 討が必要である。 本研究は,栃木県日光市のイヌブナ天然林皆伐後のクマ剥 ぎ被害を受けたヒノキ人工林,植栽針葉樹が成林せず広葉樹 二次林となった林分の2つの不成績造林地において,林分 構造調査および成長解析を行い,皆伐,除伐などの人為撹乱 下におけるイヌブナの萌芽再生過程を明らかにし,広葉樹林 化を行う場合に,イヌブナが天然更新木として有用であるか を評価することを目的とした。 2.調査地と方法 調査地は,栃木県日光市(旧栗山村)の大滝沢流域の標高 880∼1,080mに位置する東明神岳国有林である(北緯36°54′, 東経139°37′;図―1)。この地域は,帝釈山脈に属し,主に新 第三紀火成岩,中生代火成岩などからなる(貝塚・武内, 1986)。土壌は,褐色森林土で,表層は部分的に関東ローム で覆われる。気候は,太平洋側気候区に属し,冬季の降水量 が少ない。約5km離れた土呂部(標高925m,年平均気温 7.2℃,年降水量1,736mm,日本海指数38°)の気象データか ら推定すると,暖かさの指数は46.8∼62.9℃・月で,冷温帯 中・上部に位置づけられる。植生は,ブナ―ミヤコザサ群集 に位置づけられている(栃木県自然環境調査研究会植物部会 編,2003)。 調査は,1979∼1982年にイヌブナ天然林を数ha規模で小 面積皆伐し,その後に針葉樹を植栽した4つの小班,およ びこれら小班の境界にある,皆伐・造林されずに保残帯とし て残されたイヌブナ天然林で2009∼2010年にかけて行った。 各小班を調査区(C1,C2,B1,B2,保残帯をR)とした(図― 1)。皆伐前の各調査区の林分組成については,旧前橋営林局 今市営林署(日陰担当区)が伐採収穫に当って作成した主産物 調査報告書の資料を基に推定した。日光森林管理署の小班沿 革簿を基に作成した各調査区の施業履歴の概要を表―1に示し た。C1およびC2は,いずれも27年生のヒノキ人工林で,下 刈り(筋刈り)が連続して6年間(6回)行われた後に,除伐 図―1.調査地の位置図 Fig. 1. Map of study site
黒枠で囲われたグレーの区画は小面積皆伐された小班を,太線は 小班界を,点線は林道をそれぞれ表す。C1とC2はヒノキ人工林調 査区,B1とB2は広葉樹二次林調査区,Rは保残帯調査区である。 82
表 ―1 .調査区の施業履歴の概要 T a bl e 1 . O ut li ne a nd fo re st m a na g em ent hi st o ry o f st udy pl o ts *2 0 1 0 年時点の林齢を示す。 †「かぶり」は植栽木を覆う枝のみを刈る施業。 ‡除伐は行われなかったものとみなした。 83
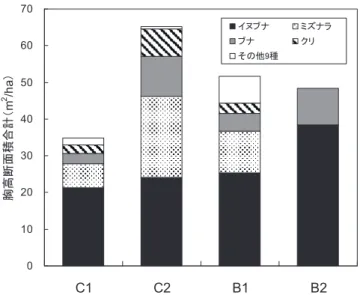
がそれぞれ2回と3回行われている。B1およびB2は,いず れも植栽されたアカマツ,ヒノキがほとんど枯死し,現在 30年生の広葉樹二次林となっている。B1は下刈り(筋刈り) が連続して3年間(3回)行われた後に,植栽木を覆う低木 のみを刈る「かぶり」とよばれる坪刈りの一種の下刈り施業 が連続して2回行われている。B2については,下刈り(筋刈 り)が連続して5年間(5回)行われた後,その翌年に「かぶ り」が1回行われている。B1は小班沿革簿では1991年に除 伐が1回行われたことになっているが,年輪解析の結果, その年にも調査区内に多くの広葉樹が生育していたと推定さ れたことから,本研究ではB1は除伐が行われなかった調査 区として扱った。なお,本研究で用いる林齢,樹齢は2010 年時点のものである。 ヒノキ人工林調査区(C1およびC2)では2010年時点で激甚 なクマ剥ぎ被害が発生している。植栽ヒノキの平均胸高直径 (以下,DBH)は14.8cm(N =83)であり,植栽ヒノキのうち DBHが11.1cm(平均DBH − 1 × 標準偏差)以上の個体につ いては79%が,平均DBH以上の個体については98%がクマ 剥ぎ被害を受けていた(松本ら,未発表)。 人為撹乱下における更新初期段階の林分構造を把握するた めに,各調査区内に等高線に平行して4m ×50mのベルト 枠を2010年に3∼4本設置した。各枠内の樹高1.3m以上の 籐本を除く高木性木本植物のDBH(cm)と樹高(m)を測定し, 樹高5m以下を低木層,樹高5∼10mを亜高木層,樹高10m 以上を高木層と定義した。なお,イヌブナのように株から多 数の萌芽幹を発生させている樹木においては,枝分かれと萌 芽の区別が困難な場合も多い。したがって,本研究では地上 0.5m以下で分岐していた幹をそれぞれ互いに1本の独立し た幹として調査した。また,ヒノキ人工林(C1およびC2)の ヒノキに関しては,ベルト枠1本あたり10本のDBHおよび 樹高を測定した。 各調査区で出現した高木性木本植物のDBHおよび萌芽に よる株構造の有無のデータを基にして,株当たりの幹本数, 胸高断面積合計(以下,BA)を求め,BAの相対優占度(%; 以下,RD)を求めた。なお,ヒノキに関してはベルト枠内の すべての立木を毎木調査していないため,測定したヒノキ生 存木の平均DBHおよび平均樹高を基に,当該プロットにお けるヒノキのBA,当該樹高階におけるヒノキの本数を推定 した。 伐採株から発生したイヌブナ萌芽幹と,その樹冠と重なる 範囲内(当該のイヌブナ株位置から半径約5m以内)に樹冠を もつ隣接広葉樹(以下,隣接木)の直径成長過程を把握するた め,円盤採取と年輪コアの採取を行った。円盤は可能な限り 地際から採取し,年輪コアは成長錐を用いて地際から50cm の位置で山側一方向から採取した。採取した円盤および年輪 コアは研究室に持ち帰り,年輪コアは木片上に木工用ボンド を用いてマウントした。円盤については,十分乾燥させた後, 電動ヤスリを用いて円盤の表面を平滑にした。さらに年輪を 読み易くするために,粗さが#400の紙やすりを用いて,年 輪に沿うようにして表面を研磨した。最後に,実体顕微鏡で 年輪数を数え,年輪測定器(Mitsutoyo AT111)を用いて年輪 幅を読み取った。そして,直径成長を累積半径成長量によっ て評価した。 3.結 果 3.1 皆伐前の林分構造 各調査区における皆伐時の収穫量調査記録を基に作成した, 皆伐時の林分構成種の胸高断面積合計(BA)を図―2に示した。 B2に関してはイヌブナとブナのみの記録しか残っていな かったため,両樹種のデータのみを示した。各種のBAはそ れぞれ異なるものの,皆伐前の各調査区では,イヌブナが最 も優占し,次いでミズナラ,ブナ,クリが優占する広葉樹天 然林であった。 3.2 樹種組成およびサイズ構成 各調査区における幹密度(本/ha),株密度(株/ha),株当り の幹本数(本/株),BA(m2 /ha),およびRD(%)と,各調査区 における優占種の樹高階分布を表―2,図―3に示した。ヒノキ 人工林の調査区であるC1およびC2では,それぞれ25種と19 種の高木性木本植物が出現した。優占種として,RDの値が 最も高かった樹種はヒノキであり,それぞれ95.7%,94.3% であった。次いで,C1ではイヌブナ(0.7%)およびアサダ (0.6%),C2ではイヌブナ(0.9%)およびミズナラ(0.7%)が優 占していた。優占種の樹高階分布において,C1およびC2と もにヒノキが亜高木層から高木層まで優占しており,高木層 はほとんどヒノキで占められていた。また,亜高木層で出現 図―2.各調査区の皆伐前の優占種の胸高断面積合計
Fig. 2. Basal area of each dominant tree species in the study plots before clear-cutting obtained from harvest records
皆伐時の主産物調査報告書(旧今市営林署)を基に作成。B2に関して はイヌブナとブナのみの記録しか残っていなかった。なお,その他 9種には,コナラ,モミ,カエデ類,アサダ,ケヤキ,サワラ,ハ リギリ,トチノキ,その他が含まれる。各小班の胸高断面積合計を その小班の標準地面積で除してヘクタール換算した。なお,B1およ びB2については標準地面積の記録された資料が抜け落ちていたため, C1およびC1の小班面積と標準地面積の比(定数 =5.4)を基に推定し た標準地面積を使用した。また,本データは伐採木のデータである ため,皆伐を行っていない保残区(R)のデータは存在しない。 84
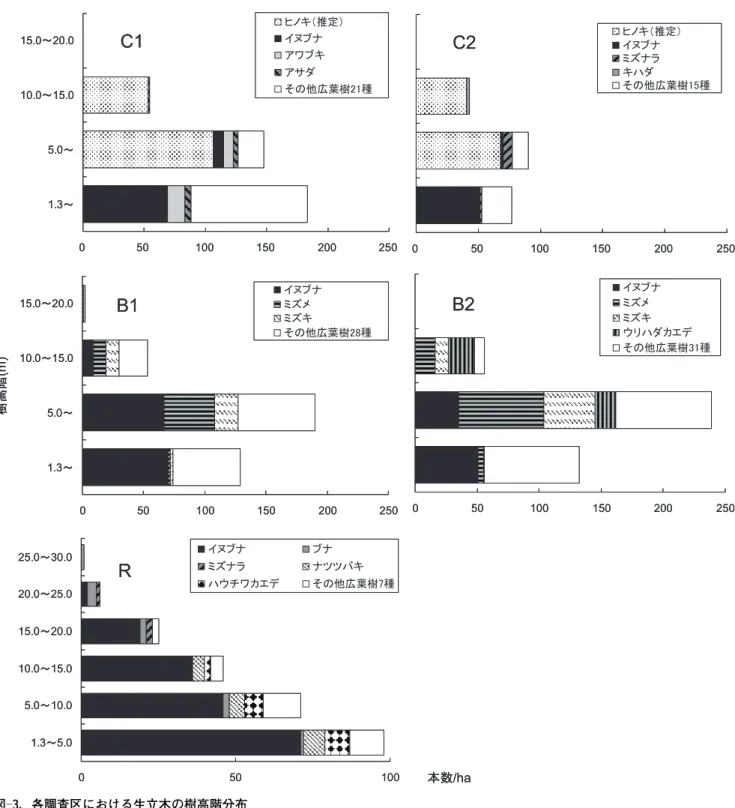
表 ―2 .各調査区における樹種構成 T a b le 2 . T ree sp ecies co m p o sit io n in th e st u d y p lo ts N ,幹密度 (本 / ha ) ; S ,株密度 (株 / ha ) ; N / S ,幹密度 / 株密度比; BA ,胸高断面積合計 ( m 2/ ha ) ; RD , BA 相対優占度 ( BA %) 85
した広葉樹はイヌブナ,アワブキ,アサダ,ミズナラおよび キハダなどであったが,その量はわずかであった。低木層で はイヌブナが最も優占しており,その他の広葉樹が20種前 後みられた。広葉樹二次林の調査区であるB1およびB2では, いずれも34種の高木性木本植物が出現した。優占種として, RDの値が最も高かった樹種は,B1ではイヌブナが22.4%, B2ではウリハダカエデが28.2%であった。次いで,B1では ミ ズ キ(16.7%)お よ び ミ ズ メ(15.0%),B2で は ミ ズ メ (18.7%)およびミズキ(17.7%)が優占していた。B1およびB2 では,ミズキ,ミズメおよびウリハダカエデは主として,亜 高木層から高木層にかけて優占しており,低木層ではあまり 見られなかった。イヌブナはB1でのみ高木層でみられた。 皆伐されていない保残帯調査区Rでは,12種の高木性木本植 物が出現した。優占種として,RDの値が最も高かった樹種 は,イヌブナ(58.0%)であった。次いで,ブナ(17.8%)およ びミズナラ(12.6%)が優占していた。Rではイヌブナが高木 層,亜高木層,低木層のすべての階層で最も優占しており, この他にブナおよびミズナラなどが高木層で見られた。 図―3.各調査区における生立木の樹高階分布
Fig. 3. Frequency distribution of height of living trees in the study plots.
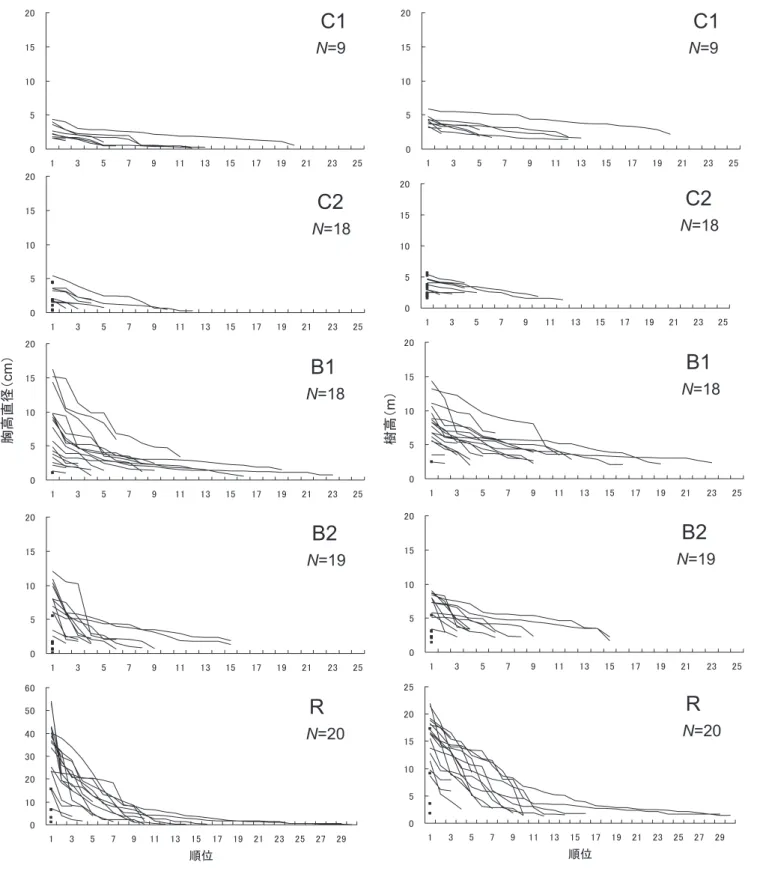
3.3 人工林および広葉樹二次林内におけるイヌブナ株 内の萌芽幹の構造 各調査区に出現したイヌブナ株内の萌芽幹のDBHおよび 樹高のサイズ構成を検討するため,各株内の萌芽幹の順位曲 線を図―4に示した。2種の順位曲線は各々の調査区では概 ね同様の曲線形を示した。C1およびC2では曲線が全体的に X軸に平行の直線型分布を示しており,株はDBH,樹高とも, ほぼ同サイズの萌芽幹で構成されていた。また,樹高10m 図―4.各調査区に出現したイヌブナ株内の萌芽幹の胸高直径(DBH)と樹高(H)の順位曲線
Fig. 4. Arrangement in the order of diameter at breast height (DBH) and height (H) of the stems in each stool of Japanese beech in the study plots
黒四角の点は単幹木(H >1.3m)を表す。
を超える萌芽幹はなく,樹高5mを超すものもわずかであっ た。C1とC2は同時期に皆伐と最近の除伐が行われたが,除 伐総数が2回のC1と3回のC2を比較すると,株内の萌芽幹 の最大DBHおよび最大樹高の差はあまりなかったが,C2の 方がC1より,株内の萌芽本数が若干少ない傾向がみられた。 特にC2では,樹高1.3m以上となった萌芽幹が1本のみのイ ヌブナ株が多く見られた。B1およびB2でのイヌブナ株内の 萌芽幹は右下がりの直線型もしくはL字型の分布を示し,樹 高10mを超える幹も存在した。このことは,株内には高木 層に達している萌芽幹も数本存在し,各イヌブナ株内では高 木層から低木層までサイズの異なる萌芽幹が連続的に分布し ていることを示す。Rは,皆伐された上述の調査区よりも DBHの順位曲線がより明瞭なL字型分布に近い曲線を示して いた。すなわち,株内には高木層に達する萌芽幹が数本存在 して,その下の亜高木層を占める萌芽幹は少なく,低木層に はサイズの小さい萌芽幹が多数存在していた。 3.4 ヒノキ人工林および広葉樹二次林内におけるイヌ ブナとその隣接木の成長過程 年輪解析によるイヌブナの各株の萌芽幹と隣接木の直径成 長過程を図―5に示した。C1およびC2では,イヌブナは複数 回の除伐後も,その年もしくは翌年から萌芽による更新を 行っていた。B1およびB2では,イヌブナ株内の萌芽幹とイ ヌブナ以外の隣接木の直径成長には大きな差がみられた。イ ヌブナや隣接木であるミズキ,ミズメ,ホオノキ,アワブキ, クマシデ,ウリハダカエデ,ヌルデ,ミズナラおよびクリの, すべての樹種が皆伐後に発生していた。これらイヌブナ以外 の樹種は皆伐後,イヌブナ株内の萌芽幹の成長よりも急速に 成長して,林冠層に到達していた。 4.考 察 4.1 ヒノキ人工林および広葉樹二次林内における広葉 樹の種組成とイヌブナの萌芽更新 イヌブナが最優占し,次いでミズナラ,ブナ,クリが優占 する広葉樹天然林を皆伐後に造成され,さらに複数回の広葉 樹の除伐がなされたヒノキ人工林調査区では,ヒノキ植栽木 を除いてイヌブナが最優占種として,特に低木層で優占して いた。除伐なしの広葉樹二次林調査区においては,イヌブナ が,亜高木層から低木層で優占し,種子の散布様式が鳥散布 型のミズキ(Masaki et al.,2004),風散布型のミズメおよびウ リハダカエデ(清野,2009;谷口,2005)といった先駆性の 高い樹種が高木層を中心に優占していた。これら3樹種は, 広葉樹二次林調査区を囲む保残帯内にはほとんどみられない ことから,鳥や風によって散布された種子が皆伐後に定着, 成長したものと考えられる。このように皆伐とその後の下刈 り・除伐によって種組成が大きく変化し,人為攪乱の強度の 高い順,すなわちヒノキ人工林,B1,B2の順に,イヌブナ の優占度が低下し,ウリハダカエデ,ミズメ,ミズキといっ た,風・鳥散布型種子をもち,先駆性を示す樹種の優占度が 高くなる傾向にあった。富山県の大辻山のスギ造林地におい て侵入した広葉樹を除伐した結果,ミズメは100%が枯死し, ウリハダカエデは43%が枯死したとの報告がある(長谷川, 1991)。陽樹で成長に十分な陽光を必要とするミズメは,樹 高成長が速い代わりに,幹の萌芽再生能力は早い時期に失わ れる(清野,2009)。また,長野県木曽地方の皆伐後にヒノキ を造林して2年間のみ下刈りを行い,その後下刈りを中止 した試験地において,ミズメと同属の先駆性樹種のウダイカ ンバは,2年間の下刈りによって主軸が切られ萌芽株を形 成したものの,そのほとんどが数年以内に枯死したとされる (原ら,1995)。したがって,ヒノキ人工林内にミズメなどの 高い先駆性をもつ樹種がほとんどみられなかったのは,ヒノ キ植栽後の下刈りや除伐といった複数回の人為撹乱のためと 考えられる。ウリハダカエデやミズキは先駆性をもつものの, 伐採(下刈り,除伐)されても萌芽再生し,2回の除伐によっ て発生が促進された例もある(長谷川,1991)。しかし,人為 攪乱頻度の高いヒノキ人工林内ではこれらの樹種がほとんど みられなかった。この理由として,ヒノキ人工林では3回 の除伐がなされたため,これらの樹種でさえ除伐後に生残で きなかった可能性が考えられる。また,ウリハダカエデはカ エデ類の中で耐陰性が低く(岩本・佐野,1998),ミズキは林 冠下で成長が悪い(正木・小谷,2009)とされる。したがって, 植栽ヒノキの林冠閉鎖による林内の光環境の悪化も,これら の樹種を消失させる原因になった可能性がある。以上のよう に,ヒノキ人工林調査区のように皆伐による強度の撹乱,ヒ ノキ植栽後の下刈りや除伐といった複数回の人為撹乱は,先 駆性を示し,萌芽再生能力の低い樹種には不利に働き,萌芽 再生能力の高いイヌブナ(大久保,2002)が結果的に生残した ものと考えられる。一方,筋刈りのような軽度の下刈りが行 われたものの,除伐が行われなかった広葉樹二次林調査区で は,萌芽再生能力の低い先駆性樹種も成長し,林冠層で優占 することができたものと考えられる。 4.2 ヒノキ人工林および広葉樹二次林内におけるイヌ ブナの成長と林分構造変化 イヌブナの株構造についてみると,ヒノキ人工林,広葉樹 二次林,保残帯の順で,萌芽幹の最大DBH,最大樹高が大 きくなる傾向を示すとともに,皆伐,下刈り,除伐といった 撹乱の有無にかかわらず,株からは常時多数の萌芽幹を発生 させた多幹構造を有していた(図―4)。このようにイヌブナは
萌芽バンク(Ohkubo et al., 1996;Bellingham and Sparrow,
2000)を形成することで,株の生残率を高めているものと考
えられる。しかし,平均萌芽本数が多い樹種ほど最大樹高が 小さい傾向にある(Nakagoshi and Wada, 1990)ように,多数 の萌芽幹の存在は,複数の幹に資源が分散され,樹高成長が 抑えられるため,隣接木との空間獲得競争において不利に なってしまうと考えられる。そのため,株内に同サイズの萌 芽幹を複数もつ株が多いヒノキ人工林および広葉樹二次林内 のイヌブナ(図―4)は,資源的な制約により,先駆性樹種より 緩慢な成長を示す(図―5)ものと考えられる。 このような萌芽・成長特性をもつイヌブナを含むヒノキ人 工林および広葉樹二次林は将来的にどのような林分に移行し ていくのだろうか。ヒノキ人工林調査区では,針葉樹植栽木 88
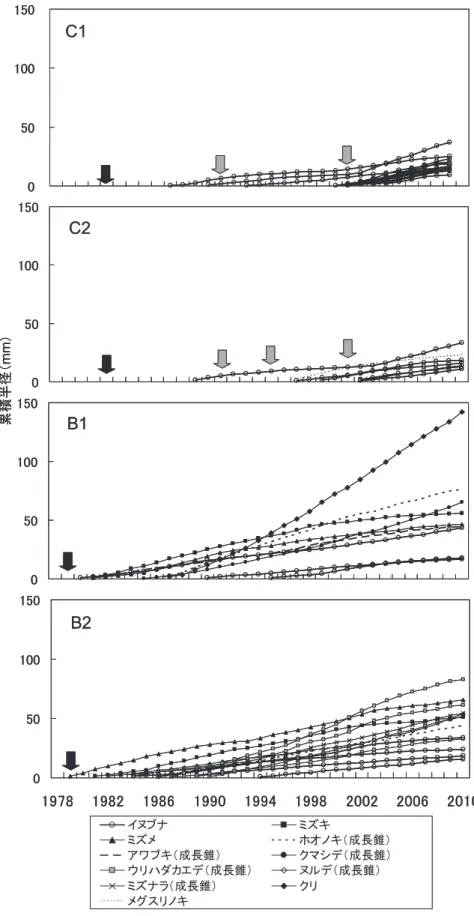
図―5.各調査区のイヌブナと隣接木樹種の累積半径成長曲線
Fig. 5. Cumulative radius growth curve of stems at 50 cm above ground of Japanese beeches and their spatially neighboring trees in the study plots
黒矢印は皆伐年を,灰色矢印は除伐年を表す。
であるヒノキの大部分がクマによる樹皮剥ぎ被害を受けてお り,被害木全体の約2割が枯死していた。ヒノキ人工林下 に残存するイヌブナの中には20年近く生残しているものも みられた(図―5)。したがって,今後,クマ剥ぎによりヒノキ が単木的に枯死することによって林冠が開けた場合は,広さ が100m2 程度のギャップ内であれば萌芽更新によって埋める ことが可能なイヌブナ(Ohkubo et al., 1988)が成長し,イヌ ブナ混交率が徐々に高まっていく可能性が考えられる。一方, 広葉樹二次林では,現在,林冠層はミズメ,ウリハダカエデ およびミズキなどの先駆樹種が優占していた。同様の傾向は, 埼玉県秩父山地のブナを交えたイヌブナ天然林の一部を皆伐 し,その後放置した林齢14年の林分においてもみられる(菊 地ら,2009)。ここでは,林冠層はウダイカンバ,ウリハダ カエデおよびミズメなどの先駆樹種が優占する一方で,下層 ではイヌブナが優占していたとされる。皆伐から31年経過 した本調査地では,イヌブナ幹の樹高は,亜高木層ないし一 部は高木層に達し,樹高10mに達する萌芽幹も数本みられ, 皆伐後14年の林分より樹高成長がみられた。このように, 時間の経過に伴い,イヌブナは,先駆性の高い広葉樹の樹冠 下で被圧に耐えて緩慢ながら樹高成長していく可能性が高い。 イヌブナは林冠を構成しうる遷移後期種とされる(大久保, 2002;澤田,2010)。現在の広葉樹二次林にはイヌブナに匹 敵する優占度をもつ遷移後期種はほとんどみられないことか ら,将来的に,イヌブナは主要な林冠構成種の1つとなる ものと推察される。 4.3 更新木としてのイヌブナの有用性 近年,人工林におけるクマ剥ぎ被害が全国的に問題化して きている(例えば,門脇ら,1997;八神,2000)。このよう な地域への被害対策としては,特に人手を入れにくい奥山地 域の不成績造林地においては針広混交林化または広葉樹林化 するなどの方法が検討され始めている。一般的に広葉樹林化 に適した広葉樹とは,寿命の長い遷移後期種や萌芽再生能力 をもつ二次林を構成する高木性樹種であるとされる(島田・ 野々田,2010)。イヌブナは遷移後期種であり,天然林内で 強い萌芽再生能力を有し,平均株直径がおよそ400cmに達 してさえも萌芽発生を繰り返しているという事例も知られて いる(Ohkubo et al., 1998;大久保,2002)。したがって,皆 伐前の林内においてイヌブナが存在すれば,強度・複数回の 人為攪乱を受けた林内においても,伐採後もイヌブナは萌芽 更新によって維持され,将来的にイヌブナが主要な林冠構成 種の1つとなる可能性が高いと考えられる。一方,ブナは 皆伐後ほとんど消失していた。これは,ブナは実生による更 新が中心(中静,1984;前田,1988)とされ,萌芽に関して は,樹齢が25年前後までに伐採すると発生するが(紙谷, 1986),樹齢50年程度以上で萌芽能力が著しく衰える(樫村 ら,1952)ためと考えられる。同様の例は,先述の埼玉県秩 父山地のブナを交えたイヌブナ天然林(イヌブナRD72.03%, ブナRD =24.65%)の皆伐後放置した林齢14年の林分でもみ られ,ここでも,皆伐後ブナは完全に消失していた(菊地ら, 2009)。このように,ブナの更新は皆伐後ほとんど期待でき ない。ミズナラやクリは,C2で皆伐前優占してはいたが, 皆伐後のヒノキ人工林内では,ヒノキを除くとミズナラはイ ヌブナについで優占していたものの,クリはみられなかった。 C1や広葉樹二次林調査区ではミズナラはイヌブナに比べて 量的にかなり少なかった。皆伐前のミズナラのサイズに関す る記録は無いが,保残帯調査区RのミズナラのDBHは38∼ 72cmと比較的サイズは大きい。本調査地とほぼ同じ標高の 落葉広葉樹林内の調査で得られた,岐阜県におけるミズナラ のサイズと樹齢の関係(小見山,1989)から,40cm程度のミ ズナラで少なくとも70年以上の樹齢をもつものと推察され る。ミズナラは少なくとも約40年生までは樹齢とともに萌 芽能力が増大するとされる(紙谷,1986)が,一般的に高齢ほ ど萌芽能力が低下することを考えると,ミズナラは皆伐前の 林分ではイヌブナに比べると量的に多くはなかった上,萌芽 発生能力が減じた高齢木を皆伐したため,萌芽を発生させた 株が少なかったものと考えられる。ブナとミズナラについて は,いずれも母樹となりうる大径木が保残帯にみられる(図― 1,表―3)が,ヒノキ人工林および広葉樹二次林調査区いずれ においても,これら優占木の実生由来によると考えられる個 体はほとんどみられなかった。ブナ・ミズナラの天然下種更 新施業で発生した稚樹は,雑草木とともに刈り払われても, 更新の支障になるほど本数を減少することはなく,むしろ, 2∼3年間の刈り払いは,両樹種の更新にとって有効であ るとされる(長谷川,1998)。しかし,連続下刈り年数が連続 3年(3回)と最も少ないB1においてもブナ・ミズナラの実 生更新はほとんどみられなかった。したがって,ブナ・ミズ ナラの実生がみられないのは,重力散布型種子(浅野・桑原, 1990)であるために,種子の散布範囲が狭く,ヒノキ人工林 および広葉樹二次林に種子がほとんど散布されなかった可能 性が考えられる。 以上のように,イヌブナは,皆伐後も下刈りや除伐のよう な複数回の人為撹乱が行われたヒノキ人工林および広葉樹二 次林のいずれの不成績造林地においても,ミズナラ,ブナ, クリといった皆伐前に優占していた他樹種と異なり,皆伐後 の林内に残存して,萌芽更新により緩慢な成長ながら個体群 を維持していた。イヌブナ以外の遷移後期種はほとんどみら れないことから,先駆的な樹種が衰退した後に,イヌブナが 主要な林冠構成樹種の1つとなることが予想される。した がって,激甚なクマ剥ぎなどの被害を受けた不成績造林地を 広葉樹林へ誘導する場合に,自然の力に任せて森林の更新を 行うことが可能という点で,残存するイヌブナは天然更新木 として有用な樹種であると考えられる。ただし,東北地方の ブナ皆伐母樹保残法施業試験地における近年の更新状況調査 の結果でみられたように,更新樹のサイズが小さい段階では, その林分が目的樹種の再生林へ推移するかどうかの判断がで きないこともある(杉田ら,2006)。本調査地は皆伐後30年 程度が経過しているとはいえ,イヌブナの萌芽幹の大部分は 高木層に達しておらず,イヌブナの更新木としての有効性の 評価を下すことは時期尚早の可能性がある。さらに長期的な 継続調査が必要である。 90
5.謝 辞 本研究を行うにあたり,林野庁関東森林管理局日光森林管 理署より小班沿革簿の閲覧と林分調査について,また,旧今 市営林署(日陰担当区)より主産物調査報告書の使用について, それぞれご許可を頂いた。また,本論文を作成するにあたり, 宇都宮大学農学部の小林幹夫教授,同大学雑草科学研究セン ターの西尾孝佳准教授より,有益なアドバイスを頂いた。同 大学農学部石栗太准教授より年輪解析に関してご協力を賜っ た。2名の査読者の方より有益なご指摘を頂いた。現地調 査の際には同大学農学部森林科学科森林生態学・育林学研究 室の皆様に御協力を頂いた。以上の方々にこの場を借りして 厚く御礼申し上げます。 引 用 文 献 浅野貞夫・桑原義晴編(1990)日本山野草・樹木生態図鑑.664pp, 全国農村教育協会,東京.
Bellingham, P. and Sparrow, A D. (2000) Resprouting as a life history strategy in woody plant communities. Oikos 89:409―416.
Del Tredici, P. (2001) Sprouting in temperate trees: A morphological and ecological review. The Botanical Review 67:121―137.
藤田 昇(1986)日本海型ブナ林と太平洋型ブナ林の構造と組成. 種生物学研究10:1―12. 藤田 昇(1987)ブナ林構成種の太平洋型分布と日本海型分布. 植物分類・地理38:311―329. 原 光好・仙石鐵也・藤森隆郎(1995)ヒノキ造林地に侵入した ウダイカンバとヒノキの二段林の成立過程.森林立地37: 89―93. 長谷川幹夫(1991)スギ不成績造林地での下刈り,除伐が広葉樹 の定着に与える影響.日本林学会誌73:375―379. 長谷川幹夫(1998)多雪地のスギ造林地に侵入したウダイカンバ の消長に及ぼす下刈,除伐の影響.日本林学会誌80:223― 228. 星 直斗(1999)丹沢山地におけるイヌブナ林の種組成と分布. 植生学会誌16:13―24. 伊藤 哲(2004)無性生殖.(樹木生理生態学,小池孝良編,264 pp.,朝倉書店,東京).185―195. 岩本慎吾・佐野淳之(1998)落葉性広葉樹二次林におけるササ現 存量と稚樹の成育様式.日本林学会誌80:311―318. 門脇正史・遠藤 徹・和出昌典・滝浪 明・大坪輝夫(1997)筑 波大学井川演習林におけるツキノワグマの生態と森林被害に 関する研究¿―クマ剥ぎ被害の実態報告―.筑波大学演習林 報告13:73―102. 貝塚爽平・武内和彦(1986)地形と地質.(日本植生誌7,関東. 至文堂,東京).55―61. 紙谷智彦(1986)豪雪地帯におけるブナ二次林の再生過程に関す る研究½À 主要構成樹種の伐り株の樹齢と萌芽能力との関 係.日本林学会誌68:127―134. 樫村大助・斎藤久夫・貴田 忍(1952)ブナ萌芽林に関する研究 ½¿ 伐採後の萌芽状況¸.第61回日本林学会大会講演集. 117―119. 清野嘉之(2009)ミズメ.(日本樹木誌.日本樹木誌編集委員会, 辻潔,東京).669―679. 菊地陽太・梶 幹男・澤田晴雄・谷本丈夫・逢沢峰昭・大久保達 弘(2009)秩父山地における林冠の撹乱規模の異なるイヌブ ナ天然林の20年間の再生過程.森林立地51:39―48. 小見山章(1989)落葉広葉樹二次林の樹齢構成とその再生過程. 日本林学会誌71:374―379. 小谷二郎(2009)多雪地帯のスギ人工林皆伐跡地において標高の 違いが木本種の定着に与える影響.森林立地51:69―76.
Kruger, L.M., Midgley J J. and Cowling, R.M. (1997) Resprouters vs
reseeders in South African forest trees; a model based on forest canopy height. Functional Ecology 11:101―105.
前田禎三(1988)ブナの更新特性と天然更新技術に関する研究. 宇都宮大学農学部学術報告46:1―79. 丸山哲也(2004)栃木県における野生動物による林業被害発生状 況º.野生鳥獣研究紀要31:70―80. 正木 隆・小谷二郎(2009)ミズキ.(日本樹木誌.日本樹木誌編 集委員会,辻潔,東京).611―633.
Masaki, T., Ota, T., Sugita, H., Oohara, H., Otani, T., Nagaike, T. and Nakanuma S. (2004) Structure and dynamics of tree populations within unsuccessful conifer plantations near the Shirakami Mountains, a snowy region of Japan. Forest Ecology and Man-agement 194:389―401.
Nakagoshi, N. and Wada, S. (1990) Population structure and succes-sion in temperate forests of southwestern Japan. Vegetatio 87:73―
84. 長池卓男(2000)人工林生態系における植物種多様性.日本林学 会誌82:407―416. 中静 透(1984)ブナ林の更新.遺伝38:62―66. 野崎玲児・奥富 清(1990)東日本における中間温帯性自然林の 地理的分布とその森林帯的位置づけ.日本生態学会誌40: 57―69.
Ohkubo, T. (1992) Structure and dynamics of Japanese beech (Fagus
japonica Maxim.) stools and sprouts in the regeneration of the
natural forests. Vegetatio 101:65―80.
大久保達弘(2002)イヌブナの萌芽特性及びイヌブナ天然林の更
新に関する研究.宇都宮大学農学部演習林報告38:1―86.
Ohkubo, T., Kaji, M. and Hamaya, T. (1988) Structure of primary Japanese beech (Fagus japonica Maxim.) forests in the Chichibu mountains, central Japan, with special reference to regeneration processes. Ecological Research 3:101―116.
Ohkubo, T., Peters, R., Sawada, H. and Kaji, M. (1998) Fagus japonica stools in natural beech forests: age structure and growth dynamics of the sprouts. Journal of Sustainable Forestry 6:143―154.
Ohkubo, T , Tanimoto, T. and Peters, R. (1996) Response of Japanese beech (Fagus japonica Maxim.) sprouts to canopy gaps. Vegeta-tio 124:1―8.
林野庁http://www.rinya.maff go.jp/j/kikaku/plan/pdf(2010年1月に
閲覧)
酒井暁子(1997)高木性樹木における萌芽の生態学的意味.種生
物学研究21:1―12.
Sasaki, Y. (1970) Versuch zur systematischen und geographischen Gliederung der Japanischen Buchenwaldgesellschaften. Vegeta-tio 20:214―249. 澤田晴雄(2010)秩父山地イヌブナ―ブナ林における構成樹種の 空間分布・更新特性とブナ類2種の豊凶特性に関する研究. 東京大学大学院農学生命科学研究科博士論文. 島田博匡・野々田稔郎(2010)暖温帯域における広葉樹林化の可 能性.森林科学59:13―16. 杉田久志・金指達郎・正木 隆(2006)ブナ皆伐母樹保残法施業 試験地における33年,54年後の更新状況―東北地方の落葉低 木型林床ブナ林における事例―.日本森林学会誌88:456― 464. 谷口真吾(2005)ウリハダカエデ(Acer rufinerve)の更新形態―多 雪地帯における伏条繁殖の事例―.日本緑化工学会誌31: 183―186. 栃木県自然環境調査研究会植物部会編(2003)とちぎの植物¿. 534pp,栃木県林務部自然環境課,宇都宮. 遠山三樹夫(1965)大室山のイヌブナ林―富士山の森林植生第À 報―.日本生態学会誌15:139―142. 八神徳彦(2000)石川県におけるクマ剥ぎ被害の現状(第1報) ―クマ剥ぎ激害地における剥皮形態―.石川県林業試験報告 31:1―9. 山本進一(1981)極相林の維持機構―ギャップダイナミクスの視 点から―.生物科学3:8―16. 横井秀一・山口 清(1998)積雪地帯のスギ不成績造林地におけ 91
るスギと広葉樹の生育実態.森林立地40:91―96.
横井秀一・山口 清(2000)積雪地帯におけるスギ不成績造林地
の取扱い―スギと広葉樹の成長からみた施業案―.森林立地
42:1―7.
Zimmerman, J.S , Everham III. E.M., Waide, R.B., Lodge, D.J., Taylor,
C M. and Brokaw, N.V.L. (1994) Responses of tree species to hur-ricane winds in subtropical wet forest in Puerto Rico: implica-tions for tropical tree life histories. Journal of Ecology 82:911―
922.