Ja panese Psycbological Research 1495, Vol.37, No.4, 210-224
Effects
of negative
contingency
upon
conditioned
sup-pression
of licking
in rats:
Systematic
manipulations
of session
length
and number
of shocks
KATSUYA
KITAGUCHI
and HIROSHI
IMADA1
Department
of Psychology,
School of Humanities,
Kwansei Gakuin University,
Nishinomiya,
Hyogo
662
The present experiments
were designed
to test the effects of negative
contingencies
of CS and
US on conditioned
suppression
of licking in rats under conditions
where P (USICS)-P
(USlnoCS)
pairs were .50-.50,
.25-.50,
and 00-.50. The contingencies
were manipulated
by matching
the
ses-sion length
in a Time-Matched
(TM) condition,
and by matching
the number
of USs in a
Number-Matched
(NM) condition.
To assess conditioned
inhibition,
the retardation
technique
was used in
Experiment
I and the summation
technique
was used in Experiment
2. The two methods
of
manip-ulating
contingencies
did not affect conditioning
differently.
Additionally,
clear evidence
of
condi-tioning of excitation
was found under zero contingency
(.50-.50). However,
.25-.50 conditions
yielded
no evidence
of conditioning,
and the inhibition
was conditioned
under 00-.50 conditions.
The results
are discussed
with reference
to possible
procedural
artifacts
and the Rescorla-Wagner
model.
Key words: negative
contingency,
conditioned
inhibition,
number
of shocks,
rat, conditioned
sup-pression
of licking
The Rescorla's contingency view of clas-sical conditioning asserts that the contingen-cy between conditioned stimulus (CS) and unconditioned stimulus (US) is critical in classical conditioning (Rescorla, 1966, 1967, 1968: Rescorla & LoLordo, 1965). Contingency is operationally defined in terms of the relationship between the probability of occurrence of US in the presence of CS (P USICS)) and in the absence of CS (P(US InoCS)). In the case of positive contingency between CS and US, or in the case where P (USICS) is greater than P (USInoCS), excita-tion is assumed to be condiexcita-tioned to the CS. On the other hand, in the case of negative contingency, or in the case where P (USInoCS) is greater than P (USICS), inhibi-tion is assumed to be condiinhibi-tioned to the CS. In either case, the amount of conditioning
should be an increasing function of the dif-ference of the two probabilities. In the case where the two probabilities are the same, there would be no conditioning.
A considerable number of experiments supported this contingency view of classical conditioning. An elaborated experiment was conducted by Rescorla (1968) under condi-tions where P (USICS) was higher than P
(USInoCS), with these probabilities varied systematically among 10 groups of rats. The probability pairs or P (USICS)-P (USInoCS)
for each of these 10 groups were .4-.4, .4-.2, .4-.1, .4-0, .2-.2, .2-.1, .2-0, .1-.1, .1-0, and 0-0. It was shown that, with a fixed P (USICS), increasing P (USInoCS) systematically atten-uated conditioned excitation. The above find-ing by Rescorla (1968), usfind-ing a technique of conditioned suppression of bar-pressing in rats, was confirmed in our laboratory by Bando (1981) using a method of the condi-tioned suppression of licking in rats.
The effects of negative contingency on conditioning were also examined by Rescorla (1969a) with the probability pairs of .4-.4, 0-' We would like to express our appreciation to Dr.
J. Bruce Overmier, Sadahiko Nakajima and Nobuyuki Kawai for their helpful advice on an earlier version of this article.
CS-US negativecontingency 211 .4, .1 -.1, and 0-.1, and by Witcher and Ayres
(1980) with probability pairs of 0, .1, 0-.8. .1-.8, .2-.8, and .6-0-.8. These studies, both using the method of conditioned suppression of bar-pressing in rats, demonstrated that higher negative contingencies between the CS and US led to a greater conditioning of inhibition.
In these previous studies, the contingen-cies were manipulated by varying the number of USs in the presence or absence of CSs. For example, in Experiment 2 of Rescorla (1968), the mean number of USs per 2-h ses-sion for condition .4-.4 was 24, whereas that for condition .4-0 was 4.8. Also in Experiment 1 of Rescorla (1969a), the mean numbers of USs per session for 0-1 and 0-4 probability pairs were 4.8 and 19.2, respec-tively. Thus, the effect of total number of
USs was inevitably confounded with the effect of contingency per se in these studies. This was also true with Witcher and Ayres (1980). The only exception was the study conducted by Hearst, Bottjer, and Walker (1980): They isolated these two variables in an experiment investigating the effects of positive (.4-0, .4-.2), negative (0-.4, .2 -.4), and no contingencies (.4-.4) on pigeons' con-ditioned approach-withdrawal behavior, with key-light as CS and food as US. They added two control groups in which the contingen-cies were the same as .4-0 or .4-.2 conditions but the total number of USs was the same as
that of .4-.4 condition. It was done by elon-gating the session length and by increasing the total number of CS trials. They demon-strated the effect of contingency unconfound-ed with the effect of total number of USs. However, the effect of difference in the total number of CS trials was still a confounding variable in assessing the effect of contingen-cy per se upon conditioning.
In the present study, in its Number-Matched (NM) condition, the effect of nega-tive CS-US contingencies upon conditioning
was assessed under the condition where both
the number of USs and that of CSs were
matched across groups of different
contin-gencies. Two experiments were conducted
with rats in a situation of the conditioned suppression of licking. The probability pairs used in the present study were 00-.50, .25
-.50, and .50-.50. The effects of the same
probability pairs were also studied using a
conventional way of manipulating CS-US
contingency; the session time was the same
for all groups and the CS-US contingencies
were manipulated by varying the number of
USs. This condition was called the
Time-Matched (TM) condition, and served as the
control for the NM condition. Experiments 1
and 2 differed only in the technique of mea-suring inhibitory tendencies; the retardation test was used in Experiment 1 and the sum-mation test was used in Experiment 2.
In an ordinary conditioned suppression
experiment, 'excitation' is indexed by the
suppression of a baseline response, such as
rats' bar-pressing or licking, by the presenta-tion of the CS paired with US, or by an exci-tor. 'Inhibition,' however, cannot be detected by this ordinary method because there should be no reason for a putative inhibitor to affect the baseline response, thus the effect is indis-tinguishable from that of a neutral stimulus. The retardation and summation tests to assess conditioned inhibition overcome this difficul-ty in the following way. The retardation test
stands on the assumption that, when a
puta-tive inhibitor is used as the CS for excitatory conditioning and paired with a US, the condi-tioning to this CS would be retarded in pro-portion to the amount of inhibition having been conditioned to that CS. The logic of the summation test is that a putative inhibitor,
when presented in compound with an excitor
of known intensity, would attenuate the
amount of conditioned response elicited by
212 K. Kitaguchi and H. Imada Experiment 1
Method
Subjects. The subjects were 50 experi-mentally naive male Wistar albino rats. The mean body weight was 341 g (range: 290-396 g) at the start of the experiment. Preexperimental handling was given to all rats for 5 min/day for 5 days.
Apparatus. The apparatus consisted of five drinking boxes (cf. Imada, Shuku, & Moriya, 1983; Nageishi & Imada, 1974), each of which was 20 cm long x 10 cm wide x 15 cm high (inside dimensions) with a grid floor composed of 5 mm stainless-steel rods spaced 13 mm from center to center. In the lower part of the wall at one narrow end of the box was a hole (12 mm diameter) through which the rat could gain access to a drinking tube. When a rat made contact with the drinking tube, a low-current circuit between the tube and the grid floor of the box was completed, which activated a contact relay located in the adjoining room. The boxes, each having a transparent ceiling, were placed on a table and each box was separated from the adjacent two by an equal distance. The drinking-hole wall of each box faced toward the center of the table. A fan below the table provided a masking noise of approximately 65 dB (C). The CS was a 20-s tone (1 000 Hz, approximately 85 dB) deliv-ered through a speaker located 200 cm above the center of the table. The US was a scram-bled electric shock of 0.7-s duration deliv-ered from an 240-V AC shock source to the
grid floor through a 250-kQ2 current-limiting resistor in series with the rat.
During drinking training sessions, a ses-sion started with the opening of the shutter attached to the outside surface of the drink-ing-hole wall and ended with its closing. During these sessions the inside walls of the box were white and the room light was on. The entire apparatus was illuminated by
indi-rect lighting, which shed light of approxi-mately 8.2-1x into each box. This environ-mental arrangement was referred to as the white-and-light context. During conditioning
sessions, black acrylic plates were placed on the inside surface of the walls of each box, and a rat was not allowed to drink water. The room light was off during these sessions. This arrangement was referred to as the black-and-dark context.
Procedure. Preliminary training of drinking (14 days). The subjects were trained to drink tap water in the drinking boxes for 5 min daily in the white-and-light context. The rats were then returned to their home cages, and 10 min later, they were allowed to drink water for another 25 min. The training was conducted for 14 days. The mean total number of licks was 1 461.08 on the last day of this training.
Negative-contingency conditioning (30 alternate days). After the completion of the preliminary training of drinking, the subjects were divided into five groups of 10 rats each. The groups were approximately matched in terms of the mean body weight and the mean total number of licks per session during the last 3 days of the above training. Conditioning took place in the black-and-dark context on alternate days and the ordi-nary drinking training in the white-and-light context intervened conditioning days. The groups differed in the CS-US contingencies; one group was trained with .50-.50 contin-gency, two groups with .25-.50 contincontin-gency, and two with 00-.50 contingency. One of the groups of each of the .25-.50 and 00-.50 con-ditions was assigned to the Time-Matched (TM) condition, where the session length was the same as, and the number of USs per ses-sion was different from, that of Group 50-50, respectively. The other group of each of these conditions was assigned to the Number-Matched (NM) condition, where the number of USs per session was the same as, and the
CS-US negative contingency 211
Table 1
Summary of the conditions for groups in each session
"8 CS segments plus 8 post-CS segments. See text for details.
session length was different from, that of Group 50-50, respectively. Thus there was a total of five groups; Groups 50-50, 25-50TM,
25-50NM, 00-50TM, and 00-50NM. Eight 20-s tone CSs were given per session for all groups.
The experimental conditions of five groups are summarized in Table 1. The ses-sion length for Group 50-50 was 640 s, which was regarded as consisting of 32 seg-ments of 20 s each. Of these 32 segseg-ments, 8 were the CS segments, 8 were the post-CS segments, and the remaining 16 were the no-CS segments. The post-CS segments, i.e., the segments immediately following the CS seg-ments, were physically the same as the no-CS segments. but were regarded functionally the same as the CS segments. This was because of a possible trace conditioning to the CS during the post-CS segment (Rescorla, 1969a). The P (USICS)2 of .50 was then accomplished by giving US in 4 of the 8 CS segments and in 4 of the 8 post-CS seg-ments. The P (USInoCS) of .50 was accom-plished by giving USs in 8 of the 16 no-CS segments. The US intensity was 120 V on the
first day of conditioning and was then increased daily in a 20-V step to 240 V. The estimated current flows, assuming the resis-tance of rats as 100 kQ, were 0.43 mA and 0.86 mA, respectively. Twelve different pre-determined sequences of CSs and USs pre-sentations were prepared and used in rota-tion.
In Groups 25-50TM and 00-50TM, ses-sion length and P (USInoCS) manipulation were the same as in Group 50-50. The P (USICS) of .25 and .00 were accomplished by giving US in 2 of the 8 CS segments and in 2 of the 8 post-CS segments in the former case, and in none of the CS and post-CS seg-ments in the latter case. Thus the number of USs per session in each of these groups dif-fered from that in Group 50-50 as shown in Table 1.
The conditions for Groups 25-50NM and 00-50NM are also summarized in Table 1. The number of USs for these groups was 16 per session as in Group 50-50. In Group 25-50NM, US was given in 2 of the 8 CS seg-ments and in 2 of the 8 post-CS segseg-ments as
in Group 25-50TM. In order to give the remaining 12 USs in no-CS segments and yet to meet the condition of P (USInoCS)=.50,
the session length was changed to 800 s in which there were 24 no-CS segments. For Group 00-50NM, the session length was changed to 960 s so as to assure 32 no-CS segments in which 16 USs were given. Thus the session length of Groups 25-50NM and 00-50NM were 1.25 times (800 s) and 1.5
One may argue that regarding post-CS segments as CS segments, as Rescorla (1969a, p.505) did, is problematic. If we regard post-CS segments as no-CS
segments, the group names, in the order of Table 1, should be changed to 00-40NM, 25-44NM, 50-50, 25-42TM, and 00-33TM. These changes, however, do
not deteriorate the value of the present study in any essential way.
214 K. Kitaguchi and Ft. Imada times (960 s) that of Group 50-50 (640 s),
respectively.
In Groups 25-50NM and 00-50NM, the temporal distribution of USs within any par-ticular session was adjusted in such a way that each inter-US interval was lengthened to 1.25 or 1.5 times longer, respectively, than that of Group 50-50. The temporal distribu-tion of CSs was then determined so as to
sat-isfy the respective contingency conditions. Twelve such sequences were used in rotation over 30 days of conditioning.
Retardation test (4 alternate days). After the completion of the negative-contin-gency conditioning, four recovery sessions
were given in the white-and-light context in the same way as in the preliminary training phase. At the end of the recovery sessions, the mean total number of licks per session recovered to 83.7% of that of the last day of the preliminary training phase.
The retardation test of inhibition was then conducted in the white-and-light context on four alternate days. During a 300-s test ses-sion, the 20-s tone CS was presented four times, two of these CSs, randomly deter-mined in each session, were followed imme-diately by a 0.7-s 120-V footshock. The trial
sequence was the same for all groups on any particular day. Ordinary drinking training sessions in the white-and-light context inter-vened conditioning sessions. If the CS had
acquired inhibitory property during the nega-tive-contingency conditioning phase, acquisi-tion of condiacquisi-tioned suppression should be retarded.
In the present study, all the statistical tests of significance were run with a rejection cri-terion of .05.
Results and Discussion
The overall suppression to each CS was indexed in terms of suppression ratios (SR) of the form B/ (A+B), where B represents the number of licks during the CS and A repre-sents the number of licks during an equally
long period immediately preceding the CS. The As and Bs were summed for all four test days, and a SR was obtained for each indi-vidual subject. The SR of 0 indicates com-plete suppression of responding during the CS, indicating strong excitation conditioned to the CS. The SR of .5 indicates no response suppression during the CS. The stronger the inhibition conditioned to the CS, the longer it would take for the CS to become an excitor. And this should be represented by slower development of conditioned suppression to the CS.
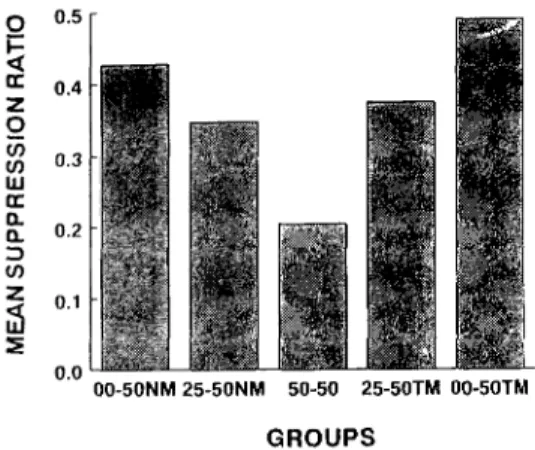
Figure 1 shows the mean SR for each group. Two tendencies are observed in Figure 1. First, there is no difference in the results between TM and NM conditions. Second, the amount of conditioned suppres-sion to the CS becomes less as the P (USICS) decreased from .50 to .00. Above two findings are consistent with the notion that the amount of classical conditioning is determined by the contingency difference as determined by P (USICS) and P (USInoCS).
A one-way ANOVA revealed a
signifi-cant difference among groups (F
(4,45)=2.84). Subsequent Fischer's LSD tests revealed that Groups 50NM, 00-50TM and 25-50NM differed significantly from Group 50-50.
Although the results shown in Figure 1
Figure 1. Mean suppression ratios as a function of negative contingency. A .50-.50 condition is common for both NM and TM conditions.
CS-US negative contingency 215
are not Inconsistent with the contingency view of classical conditioning, there are at least two problems which requires careful discussion.
The first is that, although the suppression becomes less as a function of negative con-tingency in Figure 1, this does not unequivo-cally mean that different amount of inhibition was conditioned to the CS: This result could also mean that the CSs have simply acquired different degrees of conditioned excitatory property during the negative-contingency conditioning phase. In other words, a system-atic variation of P (USICS) with a fixed value of P (USInoCS) may have resulted in a differ-ent amount of excitatory conditioning as in partial reinforcement experiments (e.g., Nageishi & Imada, 1974, Experiment 2). In order to test this possibility, the effect of the CS on licking behavior on the first trial of the retardation test was analyzed and shown in Figure 2. If any excitation has been condi-tioned to the CS during the negative-contin-gency conditioning phase, suppression should be observed on the very first trial of the retardation test. The results shown in Figure 2 clearly indicates that, at least in some of the conditions, considerable amount of excitation has been conditioned to the CS during negative contingency treatment.
Figure 2. Mean suppression ratio of each group on the first trial of the retardation test.
tailed t tests separately conducted for each group, revealed that the SRs of Group 50-50 and 00-50TM were significantly smaller than the level of no-suppression (SR=.5) (1 (9)= 5.06 and t (9)=2.27, respectively)'. The result of a simple ANOVA revealed that the groups differed significantly among themselves (F (4,45)=2.89). Subsequent Fischer's LSD tests revealed that Group 50-50 differed signifi-cantly from Groups 00-50NM and 00-50TM, but Groups 25-50TM and 25-50NM did not differ significantly from other groups.
It could be concluded, therefore, that the substantial amount of excitation was condi-tioned to the CS in Group 50-50 and moder-ate but nonsignificant excitation was condi-tioned in Groups 25-50TM and 25-50NM during the negative-contingency conditioning phase. In the 00-50 conditions, however, one cannot conclude from the results of Experiment 1 alone whether the CS was neu-tral or inhibitory, because in either case the outcome was no suppression of responding during the CS. This issue will be further examined in Experiment 2 by using the sum-mation test.
The second problem which needs to be discussed is that a retardation test cannot dis-criminate the retardation of development of conditioned suppression due to conditioned inhibition from that due to so-called "latent
inhibition" (see Lubow, 1973, for a review). In other words, it is conceivable that the CS was not much attended to or simply ignored
' The meaning of this small but significant differ-ence of Group 00-50TM from SR=.5 may be inter-preted as follows. In Group 00-50TM, due presum-ably to the smaller number of USs received (see Table 1), the pre-CS basal rate of licking was near asymptotic in many cases. So any change in licking rate for whatever reason could take place only unidi-rectionally or in the direction of suppression, which could have resulted in the significant result in Group 00-50TM.
210 K. Kitaguchi and H. Imada after repeated presentations of the CS alone
during the negative-contingency conditioning phase. This problem, which cannot be answered by using the retardation test alone, is to be solved in Experiment 2 by using the summation test. In the summation test, a con-ditioned response elicited by a known excitor should be attenuated by an addition of a puta-tive inhibitor, if it is a real inhibitor and is well attended to.
Experiment 2
Experiment 2 was conducted in the same manner as Experiment 1 except that the sum-mation test was used in order to assess condi-tioned inhibition.
Method
Subjects and apparatus. The subjects were 50 experimentally naive male Wistar rats. The mean body weight was 356 g (range: 336-384 g) at the start of the experi-ment. The apparatus was the same as in Experiment 1, except that the second CS in addition to the tone CS was used in Experiment 2. This second CS was a blinking light (170 lx, 1-s on, l-s off) generated by a 90-W lamp fixed in a lamp-shade suspended
1.5 cm above the center of the table on which the drinking boxes were placed.
Procedure. The preliminary training of drinking was conducted in exactly the same manner as in Experiment 1. The mean num-ber of licks on the last day of this preliminary training phase was 1 441.47. The subjects were then divided into Groups 50-50, 25-50TM, 25-50NM, 00-50 TM and 00-50NM of 10 rats each as in Experiment 1. They then underwent conditioning for 30 days under respective contingency condition in exactly the same way as in Experiment 1.
Excitatory conditioning to the light-CS (4 alternate days). After the completion of conditioning with the 20-s tone CS under dif-ferent contingencies, excitatory conditioning
to the 20-s light CS was given to all groups
on alternate days in the white-and-light
con-text for two days. The light CS was presented
four times in a session of 300 s and two of
these CSs, determined randomly in each
ses-sion, were followed immediately by a 0.7-s
140-V (0.5 mA) footshock. A drinking
train-ing was given in the white-and-light context
on the day intervening conditioning days.
Summation test (2 days).
Tests of the
CSs without US were conducted on two
suc-cessive days in the white-and-light context.
In each session of 300 s, one tone CS, one
light CS, and one compound CS consisting of
both tone and light were presented. The
dura-tion of each CS was 20 s. For half of the
sub-jects in each group, the light CS was
present-ed on the first trial, and the light plus tone
compound CS was presented on the second
trial. For the other half of the subjects, this
order was reversed. On the third trial, the
tone CS was presented to all subjects. On the
second day of testing, the order of the first
and second trials was reversed from that of
the first day for all subjects. The mean
inter-trial interval was 70 s (Range: 60-80 s).
Results and Discussion
During the excitatory conditioning phase,
there was clear evidence of conditioned
sup-pression to the light CS. In the test sessions,
mean SR of the two light-CS trials, when all
groups were combined, was .06. When the
tone CS, a putative inhibitor, was presented
in compound with the light CS, the mean SR
of the two compound-CS trials, again for all
groups combined, was .07. Thus no
attenua-tion of suppression was observed with the
addition of the tone to the light. It is not
unlikely that the data were subjected to the
`floor effect.'
A closer look at the licking data for the
compound-CS trials revealed, however, that
the licking behavior was suppressed the most
at the beginning of the CS, presumably due
to the orienting response to the novel
corn-CS-US negative contingency 217
pound CS. And the suppression became grad-ually less during the time-course of the CS presentation. The means of the number of licks for successive 5-s periods of the com-pound-CS presentation for all groups com-bined were 1.37, 1.73, 2.38 and 2.47. Assuming that the data of the latter half of the compound-CS presentation were least affected by theorienting response and/or by the floor effect, these data were used for fur-ther analyses.
Figure 3 compares the mean SRs of the last 10 s of the light CS and those of the compound-CS. In calculating SRs, the base-line responses (A) was the mean number of
licks per 10 s calculated for the 20-s period immediately preceding the CS in question. Upper panel shows the results for the NM
Figure 3. Mean suppression ratio to the light-CS alone and to the compound CS as a function of nega-tive contingency. The upper panel is for the NM con-dition, and the lower for the TM concon-dition, .50-.50 condition being common for both panels.
condition and the lower panel shows those for the TM condition, with the data for Group 50-50 appearing on both panels.
The results shown in Figure 3 may be summarized as follows: In Group 50-50, the conditioned suppression to the compound CS is greater than that to the light CS alone. This seems to suggest that the excitation aroused by the tone CS was added to that aroused by the light CS in the compound test. In Groups 00-50, the conditioned suppression to the compound is less than that to the light CS alone. This attenuation of conditioned sup-pression seems to suggest that the inhibition aroused by the tone CS was added to the excitation aroused by the light CS in the compound test. In Groups 25-50. the tone had virtually no effect when it was given in compound with the light CS, suggesting that the tone was neither an excitor nor an inhibitor in these groups.
A 3 x 2 ANOVA conducted for the NM condition revealed that the 'Groups' x 'Types of test-trial' interaction effect was significant (F (2,27)= 4.93). Analyses of the simple effect of types of test-trial conducted for each contingency condition revealed that the mean SR of the compound test was significantly lower than that of the light-CS test in Group 50-50 (F (1,27)=3.79) and it was significant-ly higher in Group 00-50 (F (1,27)=6.20). The results of these statistical tests confirm the summary statement described in the above paragraph. Although the same 3 x 2 ANOVA conducted for the TM condition did not reveal any statistical significance, the general picture of the graph is essentially the same as that of the NM condition.
Figure 4 shows mean SRs for the tone-CS trials on the first test day. In this case, the SRs were calculated based on the B obtained from the entire 20-s tone presentation period. A one-way ANOVA revealed a significant effect of groups (F (4,45) = 4.45). Subsequent Fischer's LSD tests revealed that
218 K.Kitaguchi and H. lmada
Group 50-50 differed significantly from Groups 00-50NM, 00-50TM and 25-50NM. Two-tailed t tests revealed that only the mean SR of Group 50-50 differed significantly from the level of no suppression or from SR=.5 (t (9)=5.56).
Taking the above results and discussions as a whole, following conclusions could be drawn from Experiment 2: (1)The two meth-ods of manipulating contingencies did not produce any difference in the results as in Experiment 1; (2)The excitation was condi-tioned to the tone-CS in Group 50-50, which is consistent with the result of Experiment 1 shown in Figure 2; and (3)The inhibition was conditioned to the tone-CS in Groups 00-50NM and 00-50TM. This last point was inconclusive from Experiment 1 alone.
General Discussion
Specific questions asked in the present study are: (1)Would conditioned inhibition take place with negative CS-US contingen-cy? (2)Would conditioned inhibition become greater as the negative CS-US contingency becomes stronger? (3)Would zero contin-gency produce any conditioning? (4)Would two methods of manipulating contingency used in the present study bring about any
dif-Figure 4. Mean suppression ratio to the tone CS alone for each group.
ference in conditioning?
To start with Question 4, the main pur-pose of the present study, the methods of manipulating contingencies did not affect conditioning differentially. It could, there-fore, be concluded that the contingency dif-ferences rather than the different number of USs accompanying contingency manipula-tion is the main factor affecting classical con-ditioning.
Conditioned inhibition observed with ..50 contingency (Groups 50NM and 00-50TM) appears to suggest the positive answer to Question 1. One may, however, argue that this evidence of inhibitory condi-tioning does not necessarily support the con-tingency view, because it could simply be an indication of inhibition conditioned to the CS- in a differential conditioning. For this reason, Witcher and Ayres (1980) placed special emphasis upon the fact that inhibition was conditioned in their .1-.8 group, where P (USICS) is higher than 0. The condition com-parable in the present study to .1-.8 group of Witcher and Ayres (1980) was .25-.50 condi-tion (Groups 25-50TM and 25-50NM). In this condition, however, no evidence of inhibitory conditioning was observed. One may argue that the present study failed to dis-tinguish the conditioned inhibition due to the differential conditioning from that due to negative contingency.
With regard to Question 3, the present experiments clearly showed that zero contin-gency (Group 50-50) yielded evidence of excitatory conditioning. This is consistent with the results from other studies (e.g., Ayres, Benedict, & Witcher, 1975 ; Benedict & Ayres, 1972 ; Keller, Ayres, & Mahoney, 1977 ; Kremer, 1971 ; Kremer & Kamin, 1974 ; Quinsey, 1971).
There seem to be three possible explana-tions for this excitatory conditioning with zero contingency. The first is a possibility of procedural artifacts. Quinsey (1971), for
CS-US negative contingency 219 example, noted that "even though one might
program a zero within-session correlation, a positive between-session correlation might
still exist. This positive correlation would arise whenever an experiment involved ses-sions without CSs and USs, together with sessions containing random and independent presentations of CSs and USs" (Witcher & Ayres, 1980, p. 73). In order to eliminate this confounding factor, we conducted drinking training in the white-and-light context, while we did the negative-contingency condition-ing in the black-and-dark context. One may still argue, however, that the difference of the two contexts was not distinctive enough. This having been so. drinking training interpolated between conditioning sessions may have led to decreased overall P (USInoCS) and thus increased between-session positive contin-gency.
The second possible explanation for con-ditioned excitation with zero contingency is an application of the theories which include some parameters other than CS-US contin-gency. As Papini and Bitterman (1990) noted, Rescorla himself substituted the Rescorla-Wagner model for his contingency view (Rescorla & Wagner, 1972). According to this model, the CS trained with zero con-tingency acquires excitatory property during the early trials, and then becomes gradually neutral. The model would hold, therefore, that the excitation observed in our Group 50- 50 was a preasymptotic effect. During the negative-contingency conditioning, the sub-jects of Group 50-50 was exposed to 30 ses-sions with zero contingency. And this num-ber of sessions might not have been large enough to neutralize the CS. Keller et al. (1977) demonstrated that some amount of excitatory conditioning to the CS occurred by a small number of training sessions, and that the CS was neutralized only after 36 sessions with zero contingency.
The third possibility that might account
for the excitatory conditioning developed to CS under zero contingency is to attribute it to accidental CS-US pairings in the early stage of training. Benedict and Ayres (1972) demonstrated, for example, that excitation is acquired when a number of CS-US pairings happened to occur early in the truly random sequence of CS and US before many non-pairings occurred. In the present experiments, however, the zero contingency condition for Group 50-50 was achieved early by the end of the first session of 640 s. Thus a possibili-ty of deflection to a positive contingency early in the training may be eliminated from the present study.
A clear conclusion of the present study is that two methods of manipulating CS-US contingency yielded the same results. As to the adequacy of the contingency view, how-ever, one cannot draw a definitive conclusion from the present study. Although Papini and Bitterman (1990) maintained that the concept of contingency might be neither necessary nor sufficient in the analysis of classical con-ditioning, further experiments should he awaited in which the difference between con-ditioning and drinking-training contexts is entirely made different, and/or in which the conditioning with zero contingency is well extended.
References
Ayres, J. J. B., Benedict, J. 0., & Witcher, E. S. 1975 Systematic manipulation of individual events in a truly random control in rats. Journal of Comparative and Physiological Psychology, 88, 97-103.
Bando, S. 1981 The effects of CS-US contingency upon licking conditioned suppression in rats. Unpublished bachelor's thesis, School of Humanities, Kwansei Gakuin University. Benedict, J. 0., & Ayres, J. J. B. 1972 Factors
affecting conditioning in the truly random control procedure in the rat. Journal of Comparative and Physiological Psychology, 78, 323-330. Hearst, E., Bottjer, S. W., & Walker, E. 1980
220 K Kitaguchi and H, Imada
Conditioned approach-withdrawal behavior and some signal-food relations in pigeons: Performance and positive vs. negative "associa-tive strength" . Bulletin of the Psychonomic Society, 16, 183-186.
Imada, H., Shuku, H., & Moriya, M. 1983. Can a rat count? Animal Learning and Behavior, 11, 396-400.
Keller, R. J., Ayres, J. J. B., & Mahoney, W. J. 1977 Brief versus extended exposure to truly random control procedure. Journal of Experimental Psychology: Animal Behavior Process, 3, 53-65. Kremer, E. F. 1971 Truly random and traditional
control procedures in CER conditioning in the rat. Journal of Comparative and Physiological Psychology, 76, 441-448.
Kremer, E. F. & Karnin, L. J. 1974 The truly random control procedure: Associative or nonassociative effects in rats. Journal of Comparative and Physiological Psychology, 74, 203-210. Lubow, R. E. 1973 Latent inhibition. Psychological
Bulletin, 79, 398-407.
Nageishi, Y., & Imada, H. 1974 Suppression of lick-ing behavior in rats as a function of predictability of shock and probability of conditioned-stimulus-shock pairings. Journal of Comparative and Physiological Psychology, 87, 1165-1173. Papini, M. R., & Bitterman, M. E. 1990 The role of
contingency in classical conditioning. Psychological Review, 97, 396-403.
Quinsey, V. L. 1971 Conditioned suppression with no CS-US contingency in the rat. Canadian Journal of Psychology, 25, 69-82.
Rescorla, R. A. 1966 Predictability and number of
pairings in Pavlovian fear conditioning, Psychonomic Science, 4, 383-384.
Rescorla, R. A. 1967 Pavlovian conditioning and its proper control procedures. Psychological Review, 74, 71-80.
Rescorla, R. A. 1968 Probability of shock in the presence and absence of CS in fear conditioning. Journal of Comparative and Physiological Psychology, 66, 1-5.
Rescorla, R. A. 1969a Conditioned inhibition of fear resulting from negative CS-US contingencies. Journal of Comparative and Physiological Psychology, 67, 504-509.
Rescorla, R. A. 1969b Pavlovian conditioned inhibi-tion. Psychological Bulletin, 72, 77-94. Rescorla, R. A. 1972 Informational variables in
Pavlovian conditioning. In G. H. Bower (Ed.), The psychology of learning and motivation. Vol. 6. New York: Academic Press. Pp.1-46. Rescorla, R. A., & LoLordo, V. M. 1965 Inhibition
of avoidance behavior. Journal of Comparative and Physiological Psychology, 59, 406-412. Rescorla, R. A., & Wagner, A. R. 1972 A theory of
Pavlovian conditioning: Variation in the effec-tiveness of reinforcement and nonreinforcement. In A. H. Black & W. F. Prokasy (Eds.), Classical conditioning II: Current research and theory. New York: Appleton-Century-Crofts. Pp.64-99. Witcher, E. S., & Ayres, J. J. B. 1980 Systematic
manipulation of US pairings in negative CS-US correlation procedures in rats. Animal Learning and Behavior, 8, 67-74.