硬骨魚雑種の形態と遺伝について II
24
0
0
全文
(2) Vol. 14. No. 1 Journal of Hokkaiclo Gakugei University (Section II A) Aug. 1963. Morphological and Genetical Observations in Hybrids of Some Teleost Fishes. II. Hiromu KOBAYASI Biological Laboratory, Asahigawa College, Hokkaiclo Gakugei University. w^mw^m ^ ,%?^ -r (n) ^ ft ^. Introduction The studies of hybrids between the funa [Carassws carassitts) or the goldfish (Carassins auratl(S) and the fishes of different species have long since been carried out by many authors for the improvement of the race and from a genetical standpoint. In hybridization investigations with the funa (Carassius carassius) and the carp (Cyprinits carpio) made by Matsui ('31, '33), it was stated that the hybrids were viable, grew rether rapidly, and showed no observable anomalies in the course of their development; they ranked intermediate to the parent species in several characteristics, being male in sterility. Suzuld ('53, '55b) dealing with embryological and morphological studies in hybridization between the loach {M.isgurnu.s angttillica.ttdatlts) and the tuna {Carassius carassius) or the golclfish (Carassius auratus), reported that hybrid eggs showed high mortality at the gastrula and hatching stages of development. The larvae from funa (goldfish) ? X loach $ survived for 24 days after hatching, while the larvae from loach ? X funa(goldfish) S were extremely abnormal, showing edema, and dying within 14 days after hatching. In development the larvae exhibited paternal influence in number of myotomes, melanophores, and the size of nuclei. Furthermore, artificial interspecific, intergeneric, inter-. familiar (intersubfamiliar), interorcler, and interclass hybridizations between the female golclfish (Carassws auratiis) and 12 kinds of male fishes, and the male frog (Rana nigromacitlala) have been performed by Suzuki ('57 b). Almost all ihe e^'gs produced of these combinations, except goldfishxfuna and goldfishxcarp, developed normally, and funaXmedaka {Oryzias latipes) and goldfishxfrog (Rana nigi'omaciilata) not developing completely, disintegrated at the gastrula stage. Only a few deformed larvae appeared, but died within 20 days after hatching. In a preliminary report with artificial hybridization between the funa (Carassws carassius) and the loach (Barbatti.la toni areas) made by Kobayasi and Yamabayashi ('58), it is reported that a part of the eggs from funa? XloachS showed no observable abnormality in the course of their development; the larvae hatched grew normally, with general external features characteristic of the funa.. Moreover, the present author continued artificial intergeneric, interfamiliar (intersubfamiliar), interorder, and interclass crossbreeding experiments between the funa (Caf'assius carcissins). ( 1).
(3) Morphologlcal and Genetical Observations in Hybrids of Some Teleost Fishes. II. and 8 kinds of fishes ; Moroco percnums, Tribolodon hakonensis, Misgiirnns angtiillicandalns, Barbattila toni areas, Leftia nikkonis, Pungitins tymensis, Oryzias lalipes and Gobiiis similis, and between the funa and the river eight-eyes lamprey {Entosphemi-s japonicii.s), the results of which are presented in this paper. Here, the author wishes to express his heartfelt thanks to Professor Tohru Uchida and Professor Sajiro Maldno, Hokkaido University, for their expert guidance with helpful advice and suggestions. The author is also very grateful to Mr. I. Yamabayashi of Fuji Gakuen Asahigawa High School, for his friendly help during the course of this study. Cordial thanks are also offered to some alumni of the Biological Laboratory, Asahigawa College, Hokkaido Gakugei University, for their technical assistance. Financial aid in part from a grant-in-aid from the Hokkaido Fund for Scientific Research is also gratefully acknowledged here.. Materials and Methods 1. Materials. The fishes providing the material for the present investigation are listed in the following : Pisces. Order Cyprinida Cyprinina (Eventognathi). Suborder. Family. Cyprinidae. Genus. Carassius. Carassius carassins (LINN'E). Species Genus. Moroco. Moroco percmirns (PALLAS). Species Genus. Tribolodon Tribolodon hakonensis (GCfNTHER). Species. Family (Subfamily) Genus. Cobitidae (Cobitinae) Misgiirmis. Misgnrmis ang'uillicaitdaitis (CANTOR). Species Genus. Barbattda Barbatu-la toni areas (JORDAN et FOWLER). Species Genus. Lefua Leftia nikkonis JORDAN et FOWLER. Species. Order Gasterosteida (Thoracostei Hemibranchu). Family. Gasterosteidae. Genus. Pnngiiws. Species. Pu.ngiiins tymensis (NIKOLSKY). Order Cyprinodontida Family Cyprinodontidae Genus Oryzias. Species Oryzias laiipes (TEMMINCK et SCHLEGEL) Order Percida. (2).
(4) Hiromu Kobayasi. Suborder Gobiina. Family Gobiidae Genus Gobitis. Species Gobins similis (GILL) Cyclostomata Order Petromyzonida Family Petromyzonidae Genus Eniosphemis. Species Eniosphemis japonicus (MARTENS) C. carassiiis : the fishes collected in the suburbs of Asahigawa and Ebeotsu during the breeding season (the beginning of June to the beginning of August) were material used. M. percnurus: the fishes collected in the suburbs of Asahigawa during the breeding season (the beginning of May to the beginning of July) were material used. T. hakonensis : the fishes collected in the Ishikari river running through the suburbs of Asahigawa and Fukagawa during the breeding season (the middle of May to the beginning of July) were material used. M. an&uillicaiidaius : the fishes collected in the Asahigawa region during the breeding season (the beginning of June to the beginning of August) were material used. B. toni areas : the fishes collected in the suburbs of Asahigawa during the breeding season (the middle of April to the end of June) were material used. L. nikkonis : the male fishes collected in the Sapporo region at the end of the breeding season (the end of June to the beginning of July) were material used. P. tymensis : the male fishes collected in the suburbs of Asahigawa at the end of the breeding season (the end of May to the beginning of June) were material used. 0. lalipes : in this study the fishes cultured in the aquarium of the laboratory were material used. G. similis : the male fishes collected in the suburbs of Asahigawa during the breeding season (the end of May to the end of June) were material used. E. japoniciis : their males collected in the Ishikari river running through the suburbs of Fukagawa during the breeding season (the beginning of June to the beginning of July) were material used. In the tables and figures of the present paper, abbreviations used are: C. c=C. carassws, M. p-=A/I. percmtt'Ks, T. h=T. Jiakonensis, M. a=M. angiiillicandatn.s, B. t.o=B. torn areas,. L. n=L. nikkonis, P. t=P. iymensis, 0. 1=0. laiipes, G. s=G. similis, E. j=E. japonicns. 2. Method of hybridization Uufertilized eggs of the following species; M. percnurus, T. hakonensis and B. toni oreas were taken out of matured females, and those of C. carassilis and M. angtl.illicail.dattis were removed from females which received injection of loach pituitary bodies, and were kept in a water bath regulated at 27°C (Kobayasi and Yamabayashi '57 b). Sperm Wcis pressed out into Ringer's solution from gonacls of a matured male in each case. Insemination was carried out. artificially at room temperatures in all cases. The eggs derived from single specimens were divided into two parts, one of which was inseminatecl with the sperm of different species for the crossbreed, and the other was left without insemination for control. In order to avoid any trouble, insemination with the sperm of the same species was performed with the eggs taken out from the distinct specimens.. (3).
(5) Morphological and Genetical Observations in Hybrids of Some Teleost Fishes, II. The combinations and numbers of crossbreeds and purebreeds in ihe present experiment are shown in Table 1. Table 1. Combination and numbers of each crossbreed and purebreed ; ( ) number of crosses undeveloped. '56 Cross C. c? xM. ps M. p ? x C. c S. July. June. 2(2). C. c <? x T. /; S. 1(4) 1. '57 May June. 2(2) 1(3). 2(3). T. /;<? xC. c S. July. 2(3). C. c ? xB. t. o S. 2(2) 2. B. t. o° xC, c S. 1(1). C. c 9 x L. n S. 2. 59. 3(2) 2(1) 3(3) 2. 2(2) 2 2 1(1) 2. June July Total. 3(1). 8(8) 5(7) 3(4) 2(3) 2 2(1) 14(5) 9(8) 8(4) 3(1) 3(1) (4). 3(1) 2(1) 2(3) 2(1). (4). C. c ? xP. f.6. (3). C. c ° x 0. / S C. c 9 x G. s S C. c ? xE. jS C. c° x. C.cS. M. po xM. p £ T. li 9 x T. li 3. 1(1) 1(1). 2(1). M. a 9 x M a S B. t. o? xB. t. oS. 1. 2 2. 1. (D. (3). 1(2) 2(1) 2(1). 2(2). 2. '62. July August. 2(4). C. c ? x M. a S M. a 9 x C. c S. '58 June July. 2. 1. 1(2) 2(1). (3) (5) (5) (3) 1 2 14(6) 3(1) 3(3) 5(1) 3(2). The fertilized eggs of each case were kept in large incubator-dishes, with 4 to 8 changes water per day. Here the embryos were detained until a complete disappearance of yolk. During the experiment the water was kept at 20° to 27°C, and PH being at 6. 2. The unfertilizecl eggs for control were kept in the same condition as the fertilized eggs. For the observation of developing eggs, some eggs were taken from the incubator-dishes at necessary intervals. The time needed for the development and mortality of eggs was expressed by average values based on the actual calculation. 3. Rearing of hybrid specimens The larvae from each cross-combination were reared in the incubator until a complete. disappearance of the yolk, and then transferred to glass vats each containing 17 liters of water. Then they were fed, and kept in the glass vats until they had grown into young fish. They were then transferred to large outdoor aquaria each containing 800 liters of water. They were fed with water-fleas and earth worms. In the winter time the larvae were kept in the basement laboratory from the middle of November to the beginning of April. The measurements of bodily characters of the hybrid specimens were made according to the system of Matsubara (55). Results of Observation 1. Development of the eggs from each combination. In the eggs of fishes used in the present experiment, the blastoderm is formed not only in. (4).
(6) I-Iiromu Kobayasi. the fertilized eggs, but also in the unfertilized eggs ; consequently the fertilization is proved for the first time at the 2-cell stage. C. carassins^ xM. percnurus S: The females 600-700 mm. in body length lay about 600 to 1000 eggs at a time. Being an opaque orange and heavier than water, each egg is spherical, measuring about 1.4 to 1.7mm. in diameter. The main bulk of the egg consists of yolk, the cytoplasm proper being an extremely thin peripheral layer around the yolk. The vitelline membrane of the egg is comparatively thick and tough, and possesses a micropyle. Soon after oviposition in water, the egg, due to stimulation by the water it has absorbed, becomes gradually transparent. The vitelline membrane is elevated from the egg proper, and begins to receive the cytoplasm. at the animal pole via a cytoplasmic stream. The vitelline membrane shows a strong adhesive power. The unfertilized eggs from three early litters used in the combination are slightly immature or hypermature, and those from two latter litters are favourably mature. In the crossbred eggs, first segmentation extends from I-;:,- to 2-^- hours after insemination.. The proportion of the unsegmented eggs at the 2-cell stage to the total number of the crossbred eggs is about 30 per cent,. Hatching. about 10 per cent more than. Appear, melano. that of the purebred eggs. The. phores .on body. Heart form.. normally developing eggs repeat. Appear. some somites. segmentation every 30 minutes.. The morula stage is reached in. Small yolli plug. about 7 hours, and the blastula. Gastrula stage. stage about 9-10 hours after. Blastuln stage. insemination. The proportion. Moruia stage. of the dead eggs observed at the blastula stage shows about. 32-ccll stage. 39 per cent of the total number. 2-cell stage. of the crossbred eggs. The gas-. 10 20 30 40 50 60 70 80 90 100 ('/»). trula stage occurs at about 12. Fig. 1 Difference in mortality rate at various stages in. -18 hours after insemination.. development of the crossbred eggs of C. cai'assins ?. During this stage only a few. x M. pe.rcnurus S, and the purebrecl eggs of the maternal line. 0 ...... C. c s? x M. ps, ®...... C. c ? x C ,c $. deaths occur, in spite of a con-. siderable number of irregularly. formed blastodiscs. The proportion of dead eggs at this stage is found to be about 43 per cent of the total number ef the crossbrecl eggs. The formation of the embryonal body of the normally developing eggs is almost identical to that of the purebred eggs. The yolk plug closes at about 27-35 hours ; 6 pairs of somites appear at about 40-45 hours ; Kupffer's vesicle at about 55-60 hours ; and pigments covering the eye cups begin to appear at about 85-90 hours after insemination. By this time the abnormal embryos have either ceased development and died, consequently the mortality rate of the eggs increases gradually during these stages. At about 100-110 hours after insemination the embryos with normal feature show. (5).
(7) Morphological and Genetical Observations in Hybrids of Some Teleost Fishes. II. heart, rudiments of the pectoral fins, and marginal fin. The proportion of the dead eggs at this stage reaches about 68-70 per cent of the total number of the crossbred eggs. By this time almost all the surviving embryos are normal in appearance. At about 10 hours after the development of the heart, asteroid melanophores form on the body, in the same way as observed in the purebred eggs. At about 148-155 hours after insemination all the normal embryos finish hatching. The proportion of the hatched embryos is about 29-31 per cent of the total number of the crossbred eggs, or about 20 per cent less than that of the purebred eggs. But the time of hatching and the process of development observed in the hatched eggs concerned here seems to similate those found in the purebred eggs. About 80 per cent of the total number of the hatched embryos is normal in form. None of the uninseminated eggs for control segmented. M. percnurns ? x C. carassius $ .• Accounts on the general character of the unfertilized eggs of. M. perow'l'iis are given with a chapter of the hybrid between M. percnurns and T. hakonensis in the previous paper (Kobayasi '62). The unfertilized eggs used in the combination are favourably mature, with the exception of slightly immature eggs from two litters in May of '57. In the crossbred eggs, first segmentation begins at 1-,:;-~2 hours after insernination. Then 90. per cent of the crossbred eggs reaches the 2-cell stage. Thereafter the segmentation is repeated every 30-40 minutes. The eggs reach the morula stage about 6 hours after insemination. At about this stage the abnormal development and death begin to increase somewhat. The bastula stage is reached about 2-3 hours after the morula stage. In the blastula stage the number of the dead eggs continues to increase gradually. The blastodiscs of some eggs appear distinctly abnormal in form. The gastrula stage is reached in about 22 hours after insemination. At this stage, the number of. Hatching. dead eggs and irregular blasto-. Appear, melano-. phores on body. discs increase as the time. Heart forri.. progresses, and the proportion. Appear some. of the dead eggs at the end of. somites. this stage shows about 37 per. Small yolk plug. cent of the total number of the. Gastruhi stage. crossbred eggs. Generally, the. Blasitula stage. time for the development of the. Morula stage. crossbred eggs since this stage begins to be slower than that. 32-cell stage. of the purebred eggs. The yolk. 2-cell stage. plug closes at about 35-38 10 20 30 40 50 60. -70 80 90 100 (%). Fig. 2 Difference in mortality rate at various stages in development of the crossbrecl eggs of M. percnurns 9 x C. carassiits 3 , and the purebred eggs of the maternal line. 0 ..... .M. pf xC. c $, ®. .. . . .M. /-)<? xM.jbS. (6). hours, 6 pairs of somites appear at about 40-48 hours after insemination. Through these stages the number of dead eggs increases more and more, and.
(8) Hiromu Kobayasi. the proportion of the dead eggs at the end of these stages reaches about 73 per cent of the total number of the crossbred eggs. Thereafter Kupffer's vesicle and optic vesicle appear. From now on, the development of all the embryo is strikingly disturbed. On this basis the development of many embryos seems to stop at this stage. About 83-85 hours after insemination, in some embryos, pigment begins to cover the eye cups and to appear on body sides. and revolving of the body and moving of the tail begin inside the vitelline membrane. But the heart differentiation in the embryos is not found. In some embryos, the distortion of the tail or body and occurrence of edema in the yolk sac have been found taking place at this time. The number of dead eggs in this stage decreases for a time. Hatching begins at about 90 hours after insemination and continues for about 10 hours. But the highly abnormal embryos die before complete collapse of the vitelline membrane, causing an increase in the death ratio at the hatching stage. The proportion of the dead eggs at this time is nearly 88 per cent of the total number of the crossbred eggs. All the hatching embryos appear with extreme deformity. The mortality rate of the purebred eggs occurs constantly at every stage a slight increase at about the gastrula stage, and the proportion of the hatching larvae to the total number of spawned eggs is about 60-65 per cent. Each larva has a heart and blood corpuscles, Not all of the uninseminated eggs for control segmented entirely. C. carassius0 xT. hakonensis S; The general character of the unfertilized eggs of C. caras-. sins is described in the foregoing. Almost all the unfertilized eggs from three litters used in this combination are slightly immature or hypermature. In this crossbreeding, the proportion of the unsegmented eggs at the 2-cell stage to the total number of the crossbred eggs is about 50 per cent. The developmental process of the segmented eggs thereafter is. Uatching. almost identical to that of the. Appear, melano-. cross between C. carassms ? and. phorcs on body. Heart form.. M. percmirns $ . The develop-. Appear, some somites. ment of the blastodisc from the morula to the blastula stage is. Small yolk plug. almost normal, with but a few. Gastrula stage. dead eggs appearing. Through. Blastula stage. the gastrula stage the dead. Morula stage. eggs increase little, and blastodiscs of abnormal form appear. 32-ccll stage. in a considerable number. The. 2-cell stage. proportion of dead eggs at this 90 100 1. Fig. 3 Diflference in mortality rate at various stages in development of crossbred eggs of C. carassins ° x T. hakonensis S , and the purebrecl eggs of the maternal. stage to the total number of the crossbred eggs is about 65 per cent. At about the "closure of. yolk plug" stage, the number of. line. 0..:... a. c ? x r. /; s, ....... c. c? xc.cs.. (7 ). dead eggs, increases gradually,.
(9) Morphological and Genetical Observations in Hybrids of Some Teleost Fishes. II. and abnormally contracted and underdeveloped embryos appear. Then the development course of the abnormal embryos slows down greatly, and their bodies become twisted. Almost all these abnormal embryos die until the time when the normal embryos begin to develop hearts. By the " closure of yolk plug " stage the embryos which seemed to be developing normally decrease in number, being about 15-20 per cent of the total number of the crossbred eggs. Afterwards, they continue normal development with the appearing of the heart, and melanophores on the body. Their hatching occurs at a similar time to that found in the purebred eggs. The proportion of the hatched embryos to the total number of the crossbred eggs is about 8-10 per cent. But almost all the hatched embryos are normal in form. None of the uninseminated eggs for control segmented. T. haJconensis^ xC. carasswsS: Accounts on the general character of the unfertilized eggs. of T. hakonensis are given with a chapter of the hybrid between T. hakonensis and M. percmirus in the previous paper (Kobayasi '62). Almost all the unfertilized eggs used in this combination were favourably mature.. In the crossbred eggs, first segmentation begins about 1-1 -^- hours aftor insemination. The proportion of the unsegmented eggs at the 2-cell stage to the total number of the crossbred eggs is observed to be about 20 per cent. Thereafter, the segmentation is repeated every 30-40 minutes. The morula. Hatching. stage is reached in about 6. Appear. 43 somites. hours, the blastula stage in. Appear, tail. about 9 hours, and the gas-. Appear.some somites. trula stage about 19-22 hours after insemination. Though. Small yolk plug. the eggs until the blastula. Gastrulu stage. stage show comparatively. Blastula stage. lower mortality, the blasto-. Morula, stage. discs of many eggs begin to appear distinctly abnormal in. 32-cell stage. form. In the gastrula stage,. Z-cfll staiiu. the number of the dead eggs 90 100. increases gradually, and the. Fig'. 4 Difference in mortality rate at various stages in. time for the development. development of crossbrecl eggs of T. liakonei/sis ? x C. carassitis S , and the purebred eggs of the maternal. becomes irregular. In the. line. 0...... T. /; 9 x C. c 6 ®,..... r, A ? x r. AS.. embryos which seemed to be developing normally, the yolk. plug closes at about 37-40 hours, and optic vesicles appear at about 50-55 hours after insemination. The number of the dead embryos begins to increase suddenly since the gastrula stage, and the mortality rate of the crossbred eggs becomes about 78 per cent at the end of these stages. By this time almost all the surviving embryos show mor or less abnormity in form. At about 65-70 hours after insemination, some embryos project the tail from the embryonic body. A slight body movement can be seem in them, Contortion of the tail or body and. (8).
(10) Hiromu Kobayasi occurrence of edema in the yolk sac of some embryos have occourred at this time. But the. number of the dead eggs at this stage decreases for a time. Hatching begins at about 78 hours after insemination and continues for about 8 hours. Because the highly abnormal embryos die before collapse of the vitelline membrane, the number of dead embryos increases at this stage too. The proportion of dead eggs at the hatching stage shows more than 90 per cent of the total number of the crossbred eggs. Moreover, almost all the hatched embryos show heavy abnormity. The mortality rate of the purebred eggs was found constantly at every stage observed here, the proportion of the dead eggs at the hatching stage being about 40-45 per cent of the total number of spawned eggs. No larva has developed a heart, blood corpuscles, or pigment on the body sides. Not all of the uninseminated eggs for control segmented entirely. C. carassius <? x M. anguillicauclatus S; The general character of the unfertilized eggs of C. carassws is described in the foregoing. Almost all the unfertilized eggs used in this combination are favourably mature, with the exception of slightly immature or hypermature eggs from two litters in July of '59. In the crossbred eggs, first segmentation occurs from 1-:,- to 2-,:,- hours after insemination.. The developmental process of the crossbred eggs by this stage is almost identical to that of the purebred eggs. The proportion of the unsegmented eggs at the 2-cell stage to the total number of the crossbred eggs is about 28 per cent, or about 8-10 per cent more than that of the purebred eggs. Thereafter the segmentation is repeated every 30 minutes, in the same manner as observed in the purebred eggs. Thirty-two-cell stage is reached in about 5 hours, the morula stage in about 7 hours, and the blastula stage about 9-10 hours after insemination. Through these stages, the developmental process of each egg progresses regularly, the dead eggs are only a few in. Hatching. number. The proportion of the. Appear, melano-. phores on body. dead eggs observed at the. Heart form.. blastul stage is about 40 per. Appear.some. cent of the total number of the. snmites. crossbred eggs. The gastrula. Small yolk plug. stage is reached in about 16. Gastrula stage. hours, and the "closure of yolk. Blastula stage. plug " stage about 33-36 hours after insemination. During these. Morula stage. stages the number of dead eggs. 32-cell stage. increases somewhat, and irregu-. 2-cell stage. larly developed embryos appear. 90 100 (%). in a considerable number.. Fig'. 5 Difference in mortality rate at various stages in. Almost all these irregulaly de-. development of crossbrecl eggs of C. carassius ? x M. (ingwllicaudatus $, and the purebrecl eggs of the. veloped embryos die gradually. maternal line. 0 ... . . . C. c ? x M aS, ...... . C. c ? x C. c S .. (9). until the time when the normal embryos begin to hatch. The.
(11) Morphological and Genetical Observations in Hybrids of Some Teleost Fishes. II. proportion of the dead eggs at the end of these stages is found to be about 55 per cent of the total number of the crossbred eggs. The formation of the embryonal body of the normally developing eggs is completely identical to that of the purebred eggs. Kupffer's vesicle appears at about 50-52 hours, pigmentp covering the eye cups beginning to appear at about 83-85 hours, and the heart and rudiment of the pectoral fins appearing at about 100-110 hours after insemination. The proportion of the dead eggs at the end of these stages is found to be about 62 per cent of the total number of the crossbred eggs. Afterwards, melanophores on the body appear, and revolving of the body and moving of the tail are repeated inside the vitelline membrane. Hatching begins about 130 hours after insemination and continues for about 20 hours. The proportion of the hatched embryos is about 35-40 per cent of the total number of the crossbred eggs, or about 15 per cent less than that of the purebred eggs. About 75 per cent of the total number of the hatched embryos is normal in borm. None of the uninseminated eggs for control developed since formation of the blastoderm. M. anguillicaudatua ? xC. caraasius S: Accounts on the general character of the unfertillized. eggs of M. angti.illicaudatii.s are given with a chapter of the hybrid between M. angn.illicaiidatns and B. toni areas in the previous paper (Kobayasi '62). The unfertilized eggs used in the combination are of favourable maturity, with the exception of slightly hypermature eggs from one litter in July of '58 and '59. In the crossbred eggs, the blastoderm is formed in about 30 minutes, and begins first segmentation about 50-60 minutes after insemination. The proportion of the unsegmented eggs at the 2-cell stage to the total number of the crossbred eggs is about 12-15 per cent. Thereafter, the segmentation follows every 15 minutes. The eggs arrive at the morula stage in about 3-77 hours, and at the blastula stage about 6 hours after insemination. During these stages, the number of the dead. Hatching i-. eggs continues to increase gra-. Appear, melano-. dually, and the proportion of the. phores on body. dead eggs at the blastula stage. Heart form. \-. to the total number of the. Appear, some somites. crossbred eggs is about 35 per. Small yolk plug. cent. The gastrula stage is. Gastruia stage. reached in about 8 hours, and. Blaslula stage. the " closure of yolk plug " stage about 13-16 hours after. Morula stage. insemination. In these stages. 32-ccll stage. the mortality rate of the eggs. 2-celt stage. increases suddenly, and the time To20io40506070" 80 90 100 (%). for the development becomes. Fig. 6 Defference in motality rate at various stages in. irregular. The proportion of the. development of crossbred eggs of M. angwUicaudattis ? x C. ciircissius S , and the purebred eggs of. dead eggs at the end of these. the maternal line. 0 ..... .M. a <? x C.c S, ® ...... M. c/ ? x M. a S .. 00). stages reaches about 73 per cent of the total number of the.
(12) Hiiomu Kobayasi. crossbred eggs. Afterwards, the majority of the remaining eggs assumes an irregular development, giving rise to undergrowth and abnormality. In the embryos which seemed to be developing normally, kupffer's vesicle appears at about 24-26 hours, pigments covering the eye cups beginning to appear at about 30-33 hours after insemination. Soon after appearance of the pigments in the eyes, the tubular heart becomes visible on the anterior side of the yolk sac. Almost all the seriously abnormal embryos have died by this stage. The proportion of the dead eggs at this stage becomes about 83 per cent of the total number of the crossbred eggs. At about 38-40 hours after insemination, the rudiments of pectoral fins appear behind the otic capsules, and the body and tail moving faintly in the vitelline membrane. Though the number of dead eggs at this stage decrease for a time, all the surviving embryos show more or less abnormity in form. Hatching begins at about 46 hours after insemination and continues for about 24 hours. At about the hatching stage, the number of the dead embryos increases again, and the proportion of the hatched embryos is about 6-8 per cent of the total number of the crossbred eggs. Furthermore, almost all the hatched embryos show extreme abnormity.. The mortality rate of the purebred eggs is constant at each stage, the proportion of the hatched embryos being about 65-70 per cent of the total number of spawned eggs. Just after hatching, each larva has a heart, blood corpuscles, and blacldsh eyes covered by pigments. None of the. uninseminated eggs for control developed. C. cwassiiis0 xB. toni oreasS; The general character of the unfertilized eggs of C. carassms. is described in the fore going. Almost all the unfertilized eggs used in this combination were favourably mature, except somewhat immature or hypermature eggs from two litters in July of '56 and one litter in June of '57. In this crossbreeding, the pro-. Hatching. portion of the unsegmented eggs. Appear, melano-. phores oh body. at the 2-cell stage is about 30. Heart form.. -35 per cent of the total number of the crossbred eggs. The. Appear, some somites. developmental process of the. Small yolk plug. segmented eggs after this stage. Gastrula stage. is almost identical to that of. Blastula stage. the crossbred eggs between. C.. Morula stage. carassuts ? and M. anguillicaitdal'its S . The development of. 32-cell stage. the eggs in early stages is al-. 2-cell stage. most normal. At the blastula .10 2.0. 30 40 50 60 70 80 90 .100 ( %). Fig. 7 Difference in mortality rate as various stages in development of crossbrecl eggs of C. ca"assins° xB. tnni. areas S , and the purebrecl eggs of the maternal line. 0...... Cc? xB. t. o 6, d ...... C. c ? x C.c S,. (.11'). stage, a few of the dead eggs and a considerable number of irregularly formed blastodiscs occur. From the gastrula stage. to the " closure of yolk plug.
(13) Morphological and Genetical Observations in Hybrids of Some Teleost Fishes. II. stage the number of the dead eggs increases, and the proportion of the dead eggs at the end of these stages becomes about 63 per cent of the total number of the crossbred eggs. Since the number of dead eggs decreases gradually, and almost all the surviving embryos by the " occurrence of heart " stage are normal in form and developmental process. Their hatching. begins at a similar time to that found in the purebred eggs, and continues longer than that of the purebred eggs. The proportion of the hatched embryos is about 30-35 per cent of the total number of ihe crossbred egg. About 70 per cent of the total number of the hatched embryos is normal in form.. None of the uninseminated eggs for control segmented. B. toni oreas ? x C. carassius 3; Accounts on the general character of the unfertilized eggs. of B.toni areas are given with a chapter of the hybrid between B. torn areas and M. angttillicaiidatits in the previous paper (Kobayasi '62). All the unfertilized eggs used in this combination are favourably mature.. In the crossbred eggs, first segmentation begins at about 2-^- hours after insemination. The proportion of the unsegmented eggs at the 2-cell stage to the total number of the crossbred eggs is about 8 per cent. Thereafter the segmentation is repeated every 40-50 minutes The eggs reach the morula stage in about 6-^- hours, the blastula stage in about 10 hours, and the gastrula stage about 22-25 hours after insemination. From the morula stage to the blastula stage a considerable dead eggs and irregularly fanned blastodiscs are found. At about the gastrula stage the mortality rate of the eggs begins to increase suddenly; the proportion of the dead eggs at this stage reached about 70 per cent of the total number of the crossbred eggs. In the surviving embryos, the " closure of yolk plug " stage arrives at about 48-53 hours, and optic vesicles appear at about 67-70 hours after insemination. Daring these stages the number of dead eggs in-. Hatcliing i-. creases more and more, and. AppCcir. mel'ano*. phores on body. almost all the surviving embryos. Heart form. ^. appearing more or less abnormal. in form. Shortly thereafter. Appear.some somjtes. some embryos project the tail. Small yolk plug. from the embryonic body. The. Gastrula stage. heart develops about 110-130. Blastula stage. hours after insemination. Con-. Manila stage. tortion of the body and edema in the yolk sac of the many. 32-cell stage. embryos have already occurred. 2-cell stage. before hatching. Hatching be60 70 80 90 100 (;'„). gins at about 200 hours after. Fig. 8 Defference in mortality rate at various stages in. insemination and continues for. development of crossbred eggs of B. toni areas ? x. about 24 hours. The proportion. C. carassiiis 3 , and the purebrecl eggs of the maternal line. ® ..... B.t.oS xC. c <?, ® ..... B.t.of xB.t.o 3.. (72). of the hatching embryos is about 4 per cent of the total.
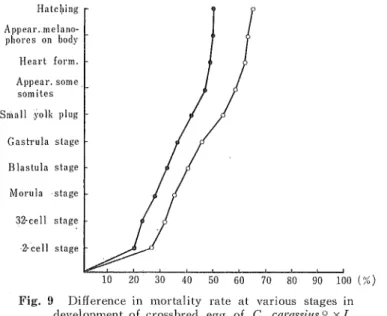
(14) Hiromu Kobavasi. number of the crossbred eggs. The majority of the surviving embryos cannot hatch by their own power, and die with a part of the body protruding from the egg into the water through the dissolution of the vitelline membrane, or die before complete collapse of the vitelline membrane. Furthermore, all hatched embryos show extreme deformity.. The mortality rate of the purebred eggs is constant at each stage except a slight increase at about the gastrula stage, and the proportion of the hatched embryos is about 65-70 per cent of the total number of spawned eggs. Each larva has developed a heart and pigment on the body sides. None of the uninseminated eggs for control developed. C. cwassius0 xL. nikkoniaS: The general character of the unfertilized eggs of C. caraswis is described in the foregoing, Almost all the unfertilized eggs used in this combination were favourably mature, with the exception of slightly immature or hypermature eggs from a litter in July of '56. The developmental process and mortality rate of these crossbred eggs bear a striking resemblance to that of the crossbred eggs between C. carassiltS 9 and M. augltilicaitdatns S . The proportion of the unsegmented eggs at the 2-cell stage is about 25-28 per cent of the total number of the crossbred Hatching. |-. T. F. _. ... ... Appear..melano-. ^ni. eggs. the developmei. phores on body. eggs in the early stages is nearly. Heart form.. normal, and the dead eggs are. Appear. some somites. only a few in number. From the gastrula stage to the " closure. Small yolk plug. of yolk plug " stage the number. Gastrula stage. of dead eggs increases little, and. Blastula stage. some embryos show abnormity. Manila. in form. The proportion of the. stage. dead eggs at the end of these. 32-cell stage. stages is about 50 per cent. 2-cell stage. of the total number of the. 90 100 (%) Fig. 9 Difference in mortality rate at various stages in development of crossbrecl egg of C. carassiiis ° x L. nikkois S, and the purebred eggs of the maternal line. 0 ...... C. c ? xL. nS, ....... C. c? xC. cS.. crossbred eggs. Thereafter the abnormal embryos die gradually. Almost all the surviving embryos when hatch are normal. in form, moving the body in the egg shell. Their hatching begins and finishes in a similar time to that found in the purebred eggs, except in a few abnormal embros. The proportion of hatched embnyos is about3 5-38 per cent of the total number of the crossbred eggs. About 87 per cent of the total number of the hatched embryos is normal in form. None of the uninseminated eggs for control segmented. C. caraasius ? x P. tymensis S, xO. latipeaS, xG, similisS, x £'. japonicus $; The eggs of the. funa (C. carassitts) were insemlnated with sperm of 3 kinds of fishes, P. tymensis, 0. laiipes,. (73).
(15) Morphological and Genetical Observations in Hybrids of Some Teleost Fishes. II. and G. si'milis, of different order, while that of the river eight-eyes lamprey (E. japonicus) of different class. But the eggs produced in these combinations did not develop at all. 2. Growth of the larvae derived from each combination Larvae derived from C. carassius ?, x M. percnurus 6, xT. hakonenais S, x M. anguillicaudatus 3,. xB. toni areas S mid xL. nikkonis S (Figs. 10-17): The larvae derived from each combination are identical in form and vitality. Just after hatching, all the larvae (except a few undergrown individuals in each combination) are normal in form, and measure about 3.2-4.0 mm. in total length. The larva hangs down. the large yolk sac from its ventral side. The eye cups become black due to the development of the melanophores. The body is straight, and 32-34 myotomes are recognized. The tail possesses a marginal fin extending straight behind. The melanophores appear scattered on the dorsal side of the body on the neighbouring region of the yolk sac. The heart beats vigorously and the red blood corpuscles are found running in the dorsal aorta and cardial vein. Because of the heavy yolk, the larva lies on its side. By this time these larvae are identical to the larva of C. carassiiis in form.. Hatching of a few abnormal larvae occurs more slowly than that of the normal ones ; the bodies are undergrown, and contorted. The abnormal larvae showed in extreme cases edema. in their yolk sac, and lack the heart. Four days after hatching, the larvae with normal appearance has absorbed one-half of its yolk, and swims around normally, settling down again at another spot. Eight days after hatching, the larvae have completed absorbing the yolk. The body becomes slender; he gets up and swims about freely. The pectoral fins grow, and begin to develop fin rays. By this time the highly abnormal larvae have gradually died. Twelve days after hatching the marginal fin is still continuous, but its caudal part is pointed and has on its ventral side mesenchymal rudiments of the caudal fin rays. The jaws develop and the mouth can now be opened and closed. At this time almost all the larvae except the abnormal ones begin to eat.. Sixteen days after hatching, the larvae generally show the dorsal lobe of caudal fin. The caudal fin assumes a circular shape. Shortly thereafter, the marginal fin shows signs of division into dorsal and anal fins. The gill-covers develop. By this time almost all the abnormal larvae have died. Twenty days after hatching, the marginal fin of the larvae begins to degenerate. Fin rays occur in the dorsal and anal fins, and the caudal fin assumes a triangular shape, the apex pointing towards the anterior part. The larvae at 25-28 days after hatching become opaque because of the pigment covering the body surface. Almost all the marginal fin disappears. The pelvic fin occurs in the remaining ventral marginal fin. The caudal, dorsal and anal fins are almost completely formed. About 30 days after hatching the larvae grow further, and all the fins completely develop. The scales appear, and the body-colour is close to that of the adult fish of C. carassms. From just after hatching to this time, in the larvae hatched with normal appearance there is almost no death. At this stage the larvae were transferred from a small indoor aquarium to large. U4^.
(16) Hiromu Kobayasi. Fig. 10-17 Growth of larvae between female funa and two kinds of male fishes 10) Normal embryos just before hatching from C. c ? xM.6S. 11) Deformed larva at 3 clays after hatching from C. c ? x M. p S . 12) Deformed larva at 6 clays after hatching from C. <7 ? xM. p6. 13) Normal larva at 1 clay after hatching from C.c ? xM. p6. 14) Normal larva at 4 clays after hatching from C. c° x M. pS. 15) Normal postlarval fishes at 20-23 clays after hatching from C. c ? x M. ff S . 16) Normal young fishes at 30-35 clays after hatcing from C. c° xM a S. 17) Normal young fish at 60 clays after hatching from C. c ¥ xM. a'6 . outdoor aquarium. No death was observed thereafter. Further development continued normally in a similar manner to that observed in pure C. cCD'assittS. Larved derivae from M. percnwus ° x, T. hakonensis ? x, M. angullicaudatus ? x, and B. toni orens ? x C. carassius S (Figs. 18-23): Just after hatching, all the larvae from each combination. are highly abnormal in form, consequently their myotomes cannot be counted distinctly. The larvae from M. psrowrus ? XC. carassius $ measure about 2.0-3.5 mm., from 7'. hakonensis ? XC. carassittsS about 3.5-5.5 mm., from M. angnillicaiidatits ? XC. carassius S about 1.02.0 mm. and from B. toni areas ? XC. carassius S about 1.8-3.0 mm. in body length. The. larvae from each combination are almost identical in form and vitality. Their body and tail are contracted and twisted to the lateral or clorsoventral side. Edema is found in a considerable area of the yolk sac. Almost all of these larvae except ones from T. hakanensis ? XC. (75-).
(17) Morphological and Genetical Observations in Hybrids of Some Teleost Fishes. II. carassizis S , melanophores appear in the eyes and on the dorsal side of the head. The development of the circulatory organ is impeded, and the heart is found at anterior part of tlie yolk sac as a slender tube or cannot be recognized at all. All the larvae lie on their side. Though they move occasionally, they do not swim.. m. a. Figs. 18-23 Deformed larvae at 3-5 days after hatching between four kinds of female fishes and male funa. 18) M.p^ xC. cS. 19) M'.pf xC. cS. 20) T./;° xC. c S. 21) M. a 9 xC.c S. 22) T. I!Q xC.c'6. 23) B.t.o? xC.c-6.. Two to three days after hatching, further differentiation and growth of the body can hardly be recognized, except for an increase of melanophores and the appearance of xantophor. The. eclema extends through the whole area of the yolk sac and into a part of the head. The twist of the trunk and tail increases more and more. The majority of hatched larvae dies by this time.. Thereafter a few surviving larvae completely stop their development and movement, and their bodyies rather begin to collapse. All die by the 7th days after hatching, u In their growth, the differentiation of the pigments proceeds most favourably, and yet it is defficult to evidence whether these carried the paternal influence or not.. (^).
(18) Hiromu Kobayasi 3. Some morphological observation of the fishes derived from C. carassins ? X M. percmir'us S , XT. hakonensis S and x5. <om" areas S. The fishes derived from the combinations between the eggs of C. carassws with the sperms of the five different kinds of fishes grew to adulthood 2 or 3 years after hatching. Thus the fishes from each combination are completely identical in external form, and the results of the morphological observations of the fishes from C. carassius ? X M. perciwriis $ , T. /W konensisS and X-B. <ow' areas 6 are here described by way of example. Table 2. Frequency of some bodily characters in hybrids of C. carassitis ? xM. percnnrus S, x T. liakonensis S and x B. font areas $ . Number of spines and soft rays of Number of Vertebral Dental formula each fin scales along ^..lf of pharyngeal D. P. V. A. C. lateral line l-uu"L "teeth. ModCc.c/o xM.pS 111,16 13 10 HI, 5 19 31 30 4-4 MoclC c. (?° xT. liS 111,16 14 9 III, 5 19 29 29 4-4. ModC c.o? xB. t. 06 111,16 15 9 III, 5 19 30 30 4-4 Table 3. Average clemension of some bodily characters in the hybrids, C. carassins ? X M. Fercnui'tis S , x T. hakonensis S and x 5. tow o<'e(?sS; except for body length, the value is expressed by the proportion of. length of each part.. Body Body Body Head Height of Diameter Lengtth. S? hei^ht breadth leneth pe^l of orbit ofsnout Average C. c° xM. ^) 3 68 3.1 5.2 3.0 7.3 12.8 12.5 AverageC.c$xT./;o 72 3.0 5.5 3.1 7.1 13.0 11.9 Average C. (; ? xB. t. o o 80 3.1 5.0 3.2 6.9 12.0 12.8. These fishes are identical in external and internal form to that of C. carassiua, as shown in Figures 24-28, and Tables 2 and 3. The bocly-height is rather pronounced. The head is relatively large but rather short in proportion to its height, while the caudal peduncle is deep. The rather small mouth is bordered by rather thick lips, and lower jaw is slightly shorter. There are no barbels. The mode of spines and rays of each fin are in the following, D. Ill, 16; A. Ill, 5; P. 14; V. 9; C. 19. The dorsal fin is inserted nearer to the caudal base than to the tip of the snout; the short anal fin is inserted below the posterior end of the dorsal fin ; the pelvic fins are inserted below the anterior end of the dorsal fin ; the pectoral fin is of symmetrical form, with bluntly rounded posterior margin ; the caudal fin is deeply emarginated. The scales are large cycloid (Fig. 27), and the number of scales along the lateral line is 28 to 34. The body colour is dark grey, with a wash of yellowish white on the sides and abdomen.. So far as certain anatomic features such as the pharyngeal teeth, the gill-rakers, the vertebral count, the liver and the intestine are concerned, the fishes from the above three combinations were identical to the funa (C. carass'his) of the maternal line. The dental formula of pharyngeal teeth is 4-4 being very close to that of the funa (Fig 26). The gill-rakers is completely identical to that of the funa (Fig. 25). The vertebral count is 28-32 exactly the same as that of the funa, and less than that of M. psrcnill'lis, T. ha'wnsnsis and B. torn areas. The shape,. size, and colour of the liver coincide with those of the funa. The coil of the intestine closely (.17-).
(19) Morphological and Gcnutical Observations in Hybrids of Som'.; Teleost Fishes. II. KsfiSF%ii ,r..7 ••. 'SwS^S. Fig. 24 Adult fish from C. carassins ? x M. percnnrus S. Figs. 25-28 Some body characters of adult fish from C. Ctiras.ii/is ? x M. percnurus S. 25) The first right gill each showing the gill rake. 26) The pharyngeal teeth. 27) A scale. 28) Coil of the intestine. resembles that of the funa, but somewhat complicated in comparison with that of M. pe>'cnn-rus and T. ha^onensis, and more complicated in comparison with that of B. toni oreas. So far as some ecological observations have gone thus far, the fishss from these combi-. ilations are identical in behavior to the funa (C. carassitts) of the maternal line. Discussion Newman ('15) performed the interfamiliar and interorder (intersuborder) hybrizations between (7.S).
(20) Hiromu Kobayasi Fimdtl-lus and some foreign teleost fishes, and reported that the considerable eggs in these combinations developed, and a few deformed larva hatched, but died presently after hatching. Suzuki ('57 b, c) observed that the interorder and interclass crossbred eggs between the female goldfish {Carassius anraius) and the male medaka (Oryzias lalipss), and the male frog (Rana nig'l'omacnlaid), and between the female carp (Cyprimis carpid) and the above males, did not develop at all. In the present study, the artificial interorder and interclass crossbreedings were made between Carassins carassms ? and Pimgiiius tymensi.s s , Oryzias laiipes 8 , Gobius similis $ and Entosphemis japomcns S . But all the eggs produced in these combination were undeveloped identical with the results of Suzuki ('57 b, c). It is probable that the fertilization in these combinations was not carried out.. In the intergeneric hybridization studies with the funa (Carassilts carassitts) and the carp {Cyprinits carpio), Matsui ('31, '33) observed that the hybrids were viable, grow rather rapidly, and ranked intermediate to the parent species in several characteristics, The intergeneric and interfamiliar (intersubfamiliar) crossbreedings between the fishes of different genera of the Cyprinidae, and between them and some fishes of the Cobitidae wer investigated by Suzuki ('53 '55 b, '56, '57 b, c). Almost all the eggs produced in these combinations, except carp (Cyprifws carpio} xgoldfish (Ca/'assiits aiiratus) which developed and grow normally, incurred breakdown at about the gastrula stage. Only a few deformed larvae hatched, but died within 24 days after hatching. The hybrid larvae from goldfish (Ca/'assittS anratns) X loach (Misgirrniis angllillicaudatns} exhibited paternal influence in number of myotomes, melanophores and the size of nuclei.. Lookimg through the results of the intergeneric and interfamiliar (intersubfamiliar) hybridizations between the funa (Ca-l'assiltS carassws) and the five kinds of fishes by the present author, a special difference between the intergeneric and interfamiliar (intersubfamiliar) hybridizations was not shown.. In the eggs from Carassiits carassius 9 X Moroco percmirus 8 , X Tribolodon hakonensis S, X Misgurnits angnillicandains S , y.Barbaiula toni oreasS, xLefua nikkonis s, each crossbreeding carried almost identical results. The fertilization rate averaged 65-70 per cent, and about 10 per cent less than that of the purebred eggs of the maternal line. But their development proceeded favorably, except the " closure of yolk plug " stage, at which appeared a rather higher rate of mortality, an average of 20-30 per cent of the total number of the crossbred eggs hatching. Almost all the larvae from these crossbreedings continued normal growth with strong resistance to disease. Their features completely resembled the funa (Carassius carassins) of the maternal line; while in the eggs from those reciprocal combinations, except Lefiia nikkonis ? xCarassitts carassinsS which was not made, the results from each crossbreeding bore a resemblance. The fertilization rate was average 80-90 per cent, and. little less than that of the purebred eggs of the maternal line. But the great number of these eggs incurred breakdown from the gastrula stage to the "appearance of some somites" stage and in the "just before hatching" stage. All but a few hatched larvae from these crossbreedings appeared in an extreme deformation, and died at about 7 days after hatching. Consequently, (.19-).
(21) Morphological and Genetical Observations in Hyclricls of Some Teleost Fishes. II. it was not evident whether they carried the paternal characteristics or not. The result from the latter combinations resembles that furnished in the intergeneric and interfamiliar (intersubfamiliar) crossbreedings made by Suzuki ('53, '55 b '57 b), in the intergeneric, interfamiliar and intersuborder hybridizations by Newman ('10, '14, '15), etc. But the evidence presented from. the former combinations does not agree with that furnished in the hybrids between the funa (Carassms carassius) and the carp {Cyprimis carpio) by Matsui ('31> '33), and between the goldfish {Carassius auratns) and the loach {Misgiirmis angiiillicaudatiis} by Suzuki ('53, '55 b). Also. a similar situation was not found in hybridization experiments made by other previous authors, so far as the present author knows.. In the hybridization between Ftindlthis heteroclitzis ? XScomber scombnis S , Newman ('18) observed that a few nearly normal embryos and many abnormal embryos developed from the crossbred eggs, the former completely lacking the paternal pigment, and the latter possessing both the parental pigments in varying degrees. It was concluded that the lack of paternal pigmeat in the former is caused by the failure to conjugate of the female with the male nucleus in insemination, or by the eviction of the male nucleus soon after conjugation, leaving only the femal nucleus; and also that the possession of both parental pigments by the latter is caused by the conjugation of the female nucleus and the male nucleus. Loeb ('12), in discussion of the inheritance of omitted paternal characteristics in hybrids between Ft.mdubis ? and Menidea 3 , Ctenolabrus S and Stenotomns $, without taking up the Cytological study of those eggs, stated that the sperm in these cases acted as parthenogenetic agent. In eggs from the cross between Functubis and Ctenolabriis, Morris ('14) observed cytologically that some chromosomes of male origin lagged at anaphase of the first and late cleavage. From the cytological study of the hybrid eggs from Fundul'us X Ctenolabrus, Stenotott.mus X CtenolahritS, Ftinchilus'xP'rionotits and Menidia x Ctenolabms, Pinney ('18, '22, '28) announced that lag and • elimination of some chromosomes of paternal origin occurred at anaphase of the first and late cleavages.. Based on the above facts, it is supposed that the occurrence of normal larvae from the crosses, Carassins carassiua ? X Moroco percmti'ns S , X Triboloclon hakonensis S , X Misgu.rmis angitillicaudat'us $ , xBarbats.da toni oreas s and xLefua nikfsoms S, may be due to nonconjugation of the female nucleus with the male nucleus in insemination, or to activation by male nucleus. The occurrence of deformed larvae from those reciprocal combinations except. Lefiia nikkonis ? X Carassws ca/'assuis S, is caused by the procrastination and elimination of some chromosomes of male origin at anaphase of the first and last cleavage after the conjugation ofthe female nucleus and the male nucleus. The cytological mechanism of these hybrids isa subject of future investigation. Summary In the artificial intergeneric and interfamiliar (intersubfamiliar) crossbreeding from Carassius carassizis ? xMoroco peromrus s , X Tribolodon hakonensis s, X Misgu'rmis an&uillicaudatzis $, xBarhahila toni areas S and XLefiia nikkonis S, an average of 60-70 per cent of the total number of these crossbred eggs began to develop, and thereafter continued developing under. (20).
(22) Hiromu Kobayasi. favorable conditions with the exception of " closure of yolk plug " stage, at which appeared a rather higher mortality rate. The percentage of eggs hatched on an average is 30 per cent, about 20 per cent less than that of the purebrecl eggs of Carassius caf'ass'MS. Almost all the larvae hatched from these combinations were apparently normal in form and viability. They possessed considerable strong resistance to disease, and resembled completely Carassws carassin's of the maternal line in form. In those reciprocal combinations (except that of Lefna nikkonis ? X Carassws carassi'us S which was not made), an average of 80-90 per cent of the total number of these crossbred eggs began to develop, but they showed a high rate of mortality, especially from the gastrula stage to the " appearance of some somites " stage and in the hatching stage. The percentage. of eggs hatched averaged 3-10 per cent. All the hatched larvae appeared in extreme deformation, and died about 7 days after hatching. It was not evident whether they carried the paternal characteristics or not.. Looking through tlie above results, a special difference between the intergeneric and interfamiliar (intersubfamiliar) hybridizations is not shown. In the interorder and mtercl&ss crossbreed'mg trom Carassius carassius 9 X Pimgitws tymens<s $ , Os'yzias laiipes S , xGobws similis S and X Entosphenits japonicns S, the eggs produced in these combinations did not develop at all. Literature BALON, K. E., DUYVENE DE WIT, J. J. & J. HOLL, 1961. The early stages of aevelopment of. the intersulfamlliar hybrid pp^^ao^&is (CyPrinic'ae). Zool. Anz., 168: 102-112. BUSS, K. & J. E. WRIGHE, 1957. Appearance and fertility of trout hybrid. Transact. Amer. Fisheries Soc., 87 172-181. DUYVENE DE WIT, J. J., 1961. Hybriclization experiments in Rhodeine fishes (Cypriniclae, Teleostei). An intergeneric hybrid between female Rhodens ocellatus and male Acanttwiiwclens atremins. Zoologica : New York Zool. Soc., 46 : 25-26. ., 1962 a. Hybridization experiments in Acheilognathine fishes (Cyprinidae, Teleostei). A comparison of the intergeneric hybrids between Tanakia tanago and Rhodens spinalis and Rlwdens oceilatns from Korea and Japan. Zoologica : New York Zool. Soc., 47 : 117-120. ., 1962 b. Characteristics of Rliodens ocellafus and Aclieilogndtlins lanceolnins hybrids. Copeia, (2): 377-390. ., 1962 c. Hybriclization experiments in Acheilognathine fishes (Cypriniclae, Teleostei). The interspecific hybrids between Acheilognatlins lauceolalns and Acheilognathns tabira. Can. Jour. ZooL, 40: 421-424. ., 1962 d. Hybriclization experiments in Acheilognathine fishes (Cypriniclae, Teleostei). Fertile intrageneric hybrids between female Acheilognathits laiiccotattis and male Aclieilognafhns linibattis from Japan. Can. Jour. Zool., 40 : 425-430. ., 1962 e. Hybridization experiments in Acheilognathine fishes (Cypriniclal, Teleostei). The hybrids between female Acheilogiuitlms /.aiiceolat.ns and Rhodens ainams and Acheilognaflins hiinanfegns. Can. Jour. Zool., 40: 255-259. HOLCIK, J & J. J. DUYVENE DE WIT, 1962. The taxonomic characteristics of hybrid Rhodeus. Copeia,. (4): 777-788.. (27).
(23) Morphological and Genetical Observations in Hybrids of Some Teleost Fishes. II HONMA, Y., 1961. Teratologic observations in Rhodeine species and hybrids (Cypriniclae) 1. Three specimens with abnormal heads. Annot.. Zool. Japan., 34: 153-157, ., & J. J. DUYVENE DE WIT, 1962. Teratologic observations in Acheilognathine specis and hybrids (Cyprinidae) II. Five cases of uncommon deformiteis in larval hybrids of bitterling. Bull. Japan. Soc. Sci. Fisheries, 28: 1056-1063. & _, 1962. Unusual ovipositor development in a female hybrid between female Acheilognathns tabira and male Acheilognathiis limbata (Cyprinidae, Teleostei). Can. Jour. ZooL,. 40: 1013-1017. HUBBS, C., & G. E. DREWRY, 1959. Survival of F. hybrids between Cyprinodont fishes, with a discussion of the correlation between hybriclization and phylogenetic relationship. Jnstitute of Marine Science, 6: 81-91. INABA, D., 1953. On the breeding and hybridization among the salmonidfishes. I. Jour. Tokyo Univ. Fisheries, 39 : 215-222. LOEB, J., 1912. Heredity in heterogeneous hybrids. Jour. Morph., 23: 1. KASANSKY, W. J., 1930 a. Zur Morphologie der hybriden von Carassms carass/'ns L. x Tinca tinca L. Zool. Anz., 75 : 235-240. ., 1930 b. Vererbung der Korpersegmentierung, Pigmentanordnung unt Strahlenzahl der Plossen bei den Hybriden der Cyprinidae. Zool, Anz., 90 : 273-284. KOBAYASI, HIR., 1957. Studies on the intergeneric crossings between 3 kind of loach in Hokkaido,. Misgmwis anguiUicaudatus (CANTOR), Barbaiula toni areas (JORDAN et FOWLER) and Lefua mkkonis JORDAN et FOWLER. (In Japanese) Jap. Jour. Ichthyology, 6 : 160-169. ., 1962. Morphological and genetical observations in hybrids of some teleost fishes. I. Jour. Hokkaido Gakugei, Univ. Sect. II B, 13 (Supplement): 1-111, _, & M. HIROSE, 1957. Ecological study on Mroco percnitftis (PALLAS), of Hokkaiclo. (In Japanese) Jour. Hokkaido Gakugei, Univ. Sect. B, 8 : 126-132. _, & S. MORIYAMA, 1957. On the development of a loach, Barbulnla foiu oreas. (JORDAN et FOWLER). (In Japanese) Jap. Jour. Ichthyology, 6 : 177-183. ., & I. YAMABAYASHI, 1957 a. On the development of the egg inseminated reciprocally. between Misgw'nus anguillicaitdatus (CANTOR) and Moroco percnw'us (PALLAS). (In Japanese) Zool. Mag., 66 : 24-28. & _, 1957 b. On the acceleration of spawing in a loach, Misgttnms. anguiilicaiulatus (CANTOR), by injection of the hypohyses of the same species. (In Japanese) Jap. Jour. Ichthyology, 6 : 170-175. _ & _, 1958. On the reciprocal inseminations between Carassiiis aiiratiis. (L.) and Burbattila tout areas (JORDAN et FOWLER). (In Japanese) Jour. Hokkaiclo Gakugei, Univ. Sect. B, 9: 137-143. KOBAYASI, HIS., & T. MAEUA, 1962. On the pharyngeal bones and their teeth in Japanese Leuciscine and Cyprinine fishes (Cyprinidae). Zool. Mag., 71 : 307-312. MAKING, S., 1939. The chromosomes of the Carp, Cyprinus carpio, including those of some related species of Cyprinidae for comparison. Cytologia, 9: 430-439. ., 1941 a. The chromosomal relation lietween two allied species of the loach (Cobitidae, Pisces). Cytologia, 12: 79-82. __, 1941 b. A karyological study of gold-fish of Japan. Cytologia, 12: 96-111. MATSUBARA, K., 1955. Fish morphology and hierarchy. Part I-III, (In Japanese) Tokyo Japan. MATSUI, Y., 1931. Genetical studies of fresh water fish. 2. On the hybrid of Cyprinus carpio L. and. (22).
(24) Hiromu Kobayasi Carassitis carass/.us (L.) (In Japanese) Jour. Imp. Fish. Exp. Station, 2: 129-137. ., 1933. Genetical studies of fresh water fish. 3. On the male sterility of hybrids of Cypri.nns carpi.o L. and Carassi.us carassins (L.). (In Japanese) Jour. Imp. Fish. Exp. Station, 5 : 52-57. _, MAKING, S. & Y. OJINA, 1956. Some cytological observations on male sterility in the carp-funa hybrids. (In Japanese) Jap. Jour. Ichthology, 5 : 52-58. MORRIS, M., 1914. The behavior of chromatin in hybrids between Ftnidnlns and Ctenolabrus. Jour. Exp. ZooL, 16: 501-521. NEWMAN, H. H., 1910. Further studies of the process of hereclity in I'nndnlns hybrids. .Tour. Exp. Zool., 8: 143-162. ., 1914 Modes of inheritance in teleost hybrids. .Tour. Exp. Zoo)., 16 : 447-499. ., 1915. Development and hereclity in heterogenic teleost hybrids. .Tour. Exp. Zoo}., 18: 511-576. _, 1918. Hybrids between Fmidnlns and mackerel. Jour. Exp. Zool., 26: 391-421. PHILLIP, J. W., et., 1923. Hybriclism of salmon iiicfens and salmon fario in Australia. Salmon and Trout mag., (31): 100-103. PINNEY, E., 1918. A study of the relation of the bahavior of the chromatin to development and hereclity in teleost hybrids. Jour. Morph., 31: 225-261. ., 1922. The initial bloach to normal development in cross fertilized egg. Jour. Morph., 36 : 401-415. ., 1928. Development al factors in teleost hybrids. Jour. Morph., 45: 579-598. SUZUKI, R., 1953. Studies on the interfamiliar crossing between loach (Misgarnas anguillicaudatus} and goldfish (Casassins aitl'atns). (In Japanese) Jap. Jour. Ichthyology, 3: 7-14. ., 1955. Not on the interfamiliar hybrid larvae between mu^l loach (Mtsgtirnus angni.lltcaudatit)s and crus ciancarp (Carassins ccn'assins} or goldfish (Carassiiis am'atns). (In Japanese) Jap. Jour. Ichthyology, 4: 50-58. ., 1956. A study on the interfamiliar crossing between minnow {Gnatlwpogon elongatiis elongains) and mud loach (Misgtirmis angnillicandatns). Sci. Rep. Aichi Gakugei Univ. Bull., 4 : 29-34. ., 1957 a. Hybriclization experiments in Cobitid fishes. Jap. Jour. Genet, 32 : 8-14. ., 1957 b. Development of golclfish eggs insemination with some foreign sperms (a preliminary report). (In Japanese) Zool. Mag., 66: 43-47. ., 1957 c. Development of carp eggs inseminatecl with some foreign sperms (a preliminary report). (In Japanese) Jap. Jour. Ichthyology, 5: 114-117. ., 1961 a. External characters of artificial intergeneric hybrids among Japan bitterlings (Cyprinicl fish). Bull. Japan. Soc. Sci. fisheries, 27 : 418-424. ., 1961 b. Sex and sterility of artificial intergeneric hybrids among bitterlings (Cyprinid fishes) Bull. Japan. Soc. Sci fisheries, 27 : 831-834. ., 1962 a. Hybridization experiment in Cyprinicl fishes I. Gaaflwf'ogoit elongatns elongatns 9 x Psendorasbora parna '6 and the reciprocal. Bull. Japan. Soc. Sci. fisheries, 28: 992-996. ,, 1962 b. On the behavior of carp-goldfise Hybrids. .Tap. Jour. Jchityology, 10: 13-15. SCHWARTZ, J. F., 1962. Artificial pike Hybrids, Bsox americanus vermiculatus x E. lacins. Transact. Amer. Fisheries Soc., 91 ••229-230.. TERAO, T. & N. HAYASHINAKA, 1961. On the artificial hybriclization among the salmonicl fishes, I. (In Japanese) Sci. Rep. Hokkaiclo Fish Hatchery, (16)51-62.. (23).
(25)
図



関連したドキュメント
The inclusion of the cell shedding mechanism leads to modification of the boundary conditions employed in the model of Ward and King (199910) and it will be
(Construction of the strand of in- variants through enlargements (modifications ) of an idealistic filtration, and without using restriction to a hypersurface of maximal contact.) At
It is suggested by our method that most of the quadratic algebras for all St¨ ackel equivalence classes of 3D second order quantum superintegrable systems on conformally flat
[11] Karsai J., On the asymptotic behaviour of solution of second order linear differential equations with small damping, Acta Math. 61
Keywords: continuous time random walk, Brownian motion, collision time, skew Young tableaux, tandem queue.. AMS 2000 Subject Classification: Primary:
This paper develops a recursion formula for the conditional moments of the area under the absolute value of Brownian bridge given the local time at 0.. The method of power series
Answering a question of de la Harpe and Bridson in the Kourovka Notebook, we build the explicit embeddings of the additive group of rational numbers Q in a finitely generated group
Then it follows immediately from a suitable version of “Hensel’s Lemma” [cf., e.g., the argument of [4], Lemma 2.1] that S may be obtained, as the notation suggests, as the m A
