原 著
三河湾六条潟におけるアサリ Ruditapes philippinarum 稚貝が
摂取する天然海水中の植物プランクトンの
サイズとその藻類群
栗田 貴代 *1 今尾 和正 *1 橋口 晴穂 *1 森 将人 *1 中田喜三郎 *2 *1 株式会社日本海洋生物研究所中部支店,〒466‒0031 愛知県名古屋市昭和区紅梅町3‒9‒2 *2 名城大学大学院総合学術研究科,〒468‒8502 愛知県名古屋市天白区塩釜口1‒501 2020 年3 月11 日受付,2020 年7 月28 日採録 AbstractIn order to ascertain the size and phytoplankton group of the food that the settled juvenile and the early juvenile of the Manila clam Ruditapes
philippinarum ingest in the sea area, Manila clams of each growth stage were bred in natural seawater collected in Mikawa Bay through
labo-ratory experiments. Changes in size-fractionated chlorophyll a content and phytoplankton composition were investigated. The experimental results show that the settled juvenile ingested cyanobacteria and nanoflagellates such as Cryptophyceae less than 10 µm, the early juvenile ingested diatoms less than 20 µm in addition to nanoflagellates, and that the juvenile clam further ingested the diatoms and dinoflagellates size to 20 µm or more. The food environment is very important for the growth and survival of juvenile Manila clams. It is necessary to understand the size composition of the phytoplankton to evaluate the quality of food environment to these clams.
Keywords: Ruditapes philippinarum, settled juvenile, phytoplankton size, size-fractionated chlorophyll a, laboratory experiments 1. はじめに アサリ Ruditapes philippinarum は,沿岸域において重 要な漁業資源である(蒲原ら,2013).近年,アサリの 漁獲量は全国的に減少しており,三河湾を主漁場とする 愛知県は全国最大の漁獲を誇っているものの,最近では 漁獲量が激減している(鈴木,2017; 蒲原ら,2018). 三河湾東奥部に位置する六条潟は,豊川河口の左岸 に発達した南北に広がる面積3.06 km2の河口干潟で(曽 根ら,2015),アサリ稚貝が毎年大量に発生する(蒲原 ら,2013; 鈴木,2017).このアサリ稚貝は,伊勢・三 河湾の各地先に移植放流され,愛知県のアサリ漁業を 支えている(蒲原ら,2013; 曽根ら,2015).六条潟に おけるアサリ稚貝に関しては,浮遊幼生の供給経路 (市川ら,2010)や,浮遊幼生来遊後の着底場所(蒲原 ら,2013),着底後の稚貝の季節による挙動の相違(蒲 原ら,2013)など,幼生や稚貝の物理的な供給メカニズ ムに関する研究のほか,稚貝の生残に及ぼす貧酸素水塊 の影響(蒲原ら,2014)や,アサリ稚貝によるプランク トン等懸濁態有機物の摂 と物質収支(曽根ら,2015) 等が報告されている.アサリ稚貝が高密度に生息する六 条潟では,その現存量を維持するため 料環境は特に重 要と考えられるが,アサリ稚貝の 料となる植物プラン クトンの種やそのサイズに関する研究は乏しい. 成貝未満のアサリが摂食する 料のサイズに関する知 見には,浮遊幼生では天然海水を用いた懸濁物除去率 (Tezuka et al., 2009)の測定例がある.着底後間もない個 体を対象とした事例では,殻長252‒572 µm の初期稚貝 を対象に,細胞の形態や大きさの異なる複数の培養され た珪藻綱の 料価値を検討した報告(伯耆,2016)があ る.稚貝では,人工 料や培養した植物プランクトンを 用いて摂 可能な 料サイズを検討した報告(Arakawa et al., 1997; Nakamura, 2004)があるが,いずれも殻長 が5 mm 以上の個体を対象にした研究である.従来アサ リの種苗生産の分野では,浮遊幼生や稚貝の 料とし てハプト藻綱の Pavlova lutheri, Isochrysis sp.,珪藻綱の
Chaetoceros gracilis, Ch. calcitrans など,細胞の大きさが
3‒10 µm 程度の植物プランクトンが用いられている(千 葉県水産研究センター,2004).このように人工 料や 培養された植物プランクトンを用いたアサリ稚貝の 料 のサイズに関する情報は散見されるものの,着底後から 稚貝までの成育段階初期のアサリが実際の海域において 摂食する植物プランクトンのサイズや種類については知 見が限定的である.一方曽根ら(2015)は,六条潟では 高密度に生息するアサリ稚貝が周辺海域からの供給や内 部生産により生成される懸濁態有機物を摂 により効率 よく底泥へ転送し,成長に転換すると指摘した.した がって,天然海域におけるアサリ稚貝の植物プランクト ンに対する摂 の実態を明らかにすることは,六条潟が 有する生物生産機構と水質浄化機能を高く維持するため の方策を検討する上で重要である. 本研究では,アサリの着底直後の稚貝から殻長15 mm 程度の稚貝までを対象に,三河湾で採取した海水を用い て室内実験を行い,実験前後のサイズ分画されたクロロ フィル a 量の変化と,植物プランクトンの種組成を調べ たところ,アサリ稚貝が摂取する植物プランクトンの細 胞サイズとその分類群について重要な知見を得たので報
告する. 2. 材料と方法
2.1 実験供試アサリの入手
実験に用いたアサリは,着底稚貝(殻長200‒300 µm, settled juvenile),初期稚貝(殻長300‒1000 µm,early ju-venile)および稚貝(殻長1‒15 mm,juvenile clam)である (英語表記以外のアサリの成育段階区分は,社団法人全 国沿岸漁業振興開発協会編,1997 による).着底稚貝お よび初期稚貝と稚貝では,実験に供するまでの入手方法 が以下のように異なる. 着底稚貝および初期稚貝は,親貝から採卵し飼育に より得た.親貝は2017 年秋季に三河湾で採取された (Table 1).採卵および飼育は種苗生産で用いられる方法 (千葉県水産研究センター,2004)に準拠した.着底稚 貝および初期稚貝の飼育には,容量80 L 程度のポリプ ロピレン(PP)製平底水槽を用いた.平底水槽の底には 濾過海水でかき混ぜ法により10 回程度洗浄された粒径 0.2 mm 未満程度の砂を敷いた.飼育時の 料には,浮 遊幼生から初期稚貝に至るまで,培養したハプト藻綱の P. lutheri を用いた.本種は細胞の大きさ3‒5 µm 程度の 単細胞の鞭毛藻類である.実験前日に網目250 µm 等の ステンレス製ふるいを用いてアサリと砂を分離し,アサ リは濾過海水を満たした水槽に収容して実験開始まで無 給 で馴致した. 稚貝は,三河湾奥部の豊川河口域(Fig. 1,◇印)にお いて潜水により採取された.底泥採取後,冷暗所条件下 で実験室に搬入し,網目1 mm のステンレス製ふるいを 用いて底泥からアサリを分離し,濾過海水を入れた水槽 に収容し実験開始まで数日間飼育した. 料は,培養 した P. lutheri を用いた.実験前日から実験開始までは, 無給 で馴致した. 2.2 実験海水の採取 実験に用いた海水は,三河湾のアサリの産卵期であ る2017 年10 月から11 月にかけて合計3 回採取された (Table 1).採取場所は Fig. 1 に示す3 地点(図中●印)で, 三河湾奥部(St.A, CDL −1.0 m),六条潟の縁辺部(St.B, CDL −2.0 m)および三河湾中央部(St.C, CDL −13.4 m)で ある.採水は上げ潮時に行った.実験に際し,可能な限 り高濃度の 料を確保するため,採水前に多項目水質計 (AAQ-RINKO,JFE アドバンテック株式会社製)を用い Fig. 1 Location of sampling stations in Mikawa Bay. Solid circles
(●) show sampling stations of experimental natural seawa-ter. Open diamond (◇) shows the sampling area of juvenile clams. Numerals in the charts represent ground height (CDL (m))
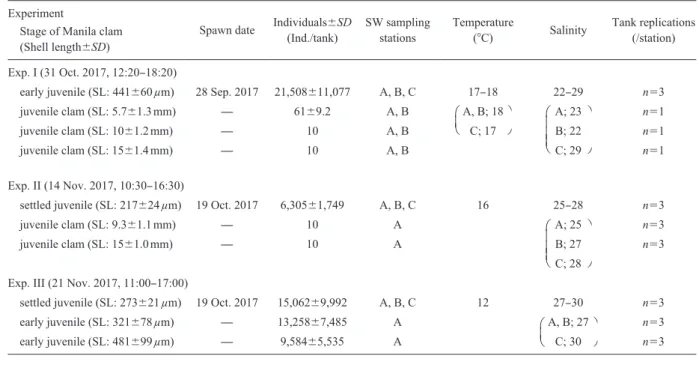
Table 1 Summary of experimental conditions Experiment
Spawn date Individuals±SD (Ind./tank) SW sampling stations Temperature (°C) Salinity Tank replications (/station) Stage of Manila clam
(Shell length±SD)
Exp. I (31 Oct. 2017, 12:20‒18:20)
early juvenile (SL: 441±60 µm) 28 Sep. 2017 21,508±11,077 A, B, C 17‒18 22‒29 n=3
juvenile clam (SL: 5.7±1.3 mm) ̶ 61±9.2 A, B A, B; 18 A; 23 n=1
juvenile clam (SL: 10±1.2 mm) ̶ 10 A, B C; 17 B; 22 n=1
juvenile clam (SL: 15±1.4 mm) ̶ 10 A, B C; 29 n=1
Exp. II (14 Nov. 2017, 10:30‒16:30)
settled juvenile (SL: 217±24 µm) 19 Oct. 2017 6,305±1,749 A, B, C 16 25‒28 n=3
juvenile clam (SL: 9.3±1.1 mm) ̶ 10 A A; 25 n=3
juvenile clam (SL: 15±1.0 mm) ̶ 10 A B; 27 n=3
C; 28 Exp. III (21 Nov. 2017, 11:00‒17:00)
settled juvenile (SL: 273±21 µm) 19 Oct. 2017 15,062±9,992 A, B, C 12 27‒30 n=3
early juvenile (SL: 321±78 µm) ̶ 13,258±7,485 A A, B; 27 n=3 early juvenile (SL: 481±99 µm) ̶ 9,584±5,535 A C; 30 n=3
て最大クロロフィル a(Chl.a)量の水深を把握し,その水 深においてバンドーン型採水器により各地点80 L 程度採 水した.採水層は,各地点とも海面下0.5‒2 m がほとんど であった.採取した海水を遮光した密閉容器に収容し, 冷暗所条件下でただちに実験室に搬入した. 2.3 実験条件 実験は上述の実験海水を採取した日と同日に,計3 回 実施された.各実験の設定状況を Table 1 に,実験の模 式図を Fig. 2 に示す.実験は開始から終了まで,採水当 日に終了させた.採取された海水中の懸濁物のばらつ きをなくすため,地点ごとにいったんポリカーボネイ ト(PC)製100 L 円形水槽にまとめ,攪拌しながら各試 験区の PC 製7 L 円形水槽に6 L ずつ分注した.試験区 当たりの試料数は,基本的に3 であるが,稚貝を対象に した実験では,供試稚貝の採集状況に応じて1 もしくは 3 に設定した(Table 1).全試験区の7 L 水槽を水浴にし た平底水槽に並べ,アサリを投入して実験を開始した. 実験室内は遮光し,各水槽には海水循環を促すため微弱 なエアレーションを施した. 料プランクトンが実験中 に沈降しないようエアレーションを強くすると,着底稚 貝および初期稚貝が動きストレスを受けることが懸念さ れた.それを回避するためエアレーションは微弱にし た.実験終了はいずれも6 時間後である.実験前後に試 水中の植物プランクトン現存量,分画 Chl.a 量および動 物プランクトン現存量を測定した. 2.4 試料の採取および測定・計数 サ イ ズ0.2‒2 µm の ピ コ プ ラン クトン(Sieburth et al., 1978)に該当するシアノバクテリアを対象に,7 L 円形水 槽から採水し,試料を50 mL 滅菌済容器に収容して,最 終濃度1% になるよう濾過滅菌したグルタルアルデヒド で固定した.DAPI(最終濃度1 µg·mL−1)染色後,Sudan black で黒暗色に染色したポリカーボネート製フィルター (Whatman ニュークリポアメンブレン,GE Healthcare UK, Ltd.,孔径0.2 µm)上に濾過捕集し,無蛍光イマルジョン オイルで包埋しプレパラートを作成した.落射式蛍光顕微 鏡を用い,B 励起光下で細胞数を計数し,1 細胞当たりの 炭素量(250 fgC·cell−1(Kana & Glibert, 1987))から炭素現
存量を推定した. ナノプランクトン(サイズ2‒20 µm),ミクロプランク トン(サイズ20‒200 µm)およびメソプランクトン(サ イズ200 µm‒20 mm)のサイズ(Sieburth et al., 1978)の植 物プランクトン(シアノバクテリアを除く)では,円形 水槽から採水した試料1 L をポリエチレン製容器に収容 後,最終濃度2% になるようグルタルアルデヒドで固定 し,静置沈殿による試料濃縮を行い,位相差装置付正 立顕微鏡を用いて形態により種の同定および細胞数の 計数を行った.また,細胞の大きさを計測して,宮井 ら(1988)の細胞の長径および短径による体積換算の近 似式に準じて平均細胞体積を算出し,Strathmann(1967), Nakamura(1998)に準じて炭素現存量を推定した.植物 プランクトンの炭素現存量は,後述の分画 Chl.a と比較 するため,出現種の1 細胞当たりの体積から同じ体積の 球に換算した際の直径(等価球径)を算出して4 つに区 分し(0.2‒2 µm, 2‒10 µm, 10‒20 µm, 20 µm<),それぞれ の炭素現存量を推定した.なお,植物プランクトンでは 従属栄養性の種類は炭素現存量推定の対象から除外し た. Chl.a では円形水槽から500 mL 程度採水し,遮光し たポリエチレン製容器に収容して測定に供した.サイ ズ別の Chl.a 量を把握するため,ポリカーボネート製 フィルター(Whatman ニュークリポアメンブレン,GE Healthcare UK, Ltd.,孔径0.2 µm, 2 µm, 10 µm の3 種 )お Fig. 2 The schematic diagram for experimental conditions. The experiment was started by putting natural seawater into three water tank at
よびナイロン製プランクトンネット(網目20 µm)を用い て試料300 mL を濾過し計4 つの分画(0.2‒2 µm, 2‒10 µm, 10‒20 µm, 20 µm<)を得た.Chl.a は,各フィルターとも 蛍光法(Strickland & Parsons, 1968)により測定した.実 験開始時および終了時の各分画の Chl.a 量と,実験に供 したアサリの個体数から,以下の式(1)により Chl.a 変化 量(pg·ind.−1·h−1)を算出した.なおアサリは実験終了後 に全個体をプランクトンネット等で捕集し,5% 中性ホル マリンで固定後,個体数の計数と殻長の計測を行った. 0 Chl.a =(E Et−×)×V t N 変化量
(1) Et:実験終了時の Chl.a(pg·L−1),E0:実験開始時の Chl.a(pg·L−1), V:試水容積(L),t:実験時間(h),N:水槽に投入し たアサリ個体数 動物プランクトンでは,植物プランクトンと同様に試 料の採取および濃縮を行い,正立顕微鏡を用いて形態に より種の同定および計数を行った.出現種ごとに必要な 部位の長さを計測し,重量‒長さの関係式および重量‒炭 素現存量の関係式(弘田,1986 など)を用いて炭素現存 量を推定した. 2.5 入水管口径の測定 アサリ稚貝が物理的にどの程度のサイズの懸濁物を摂 取可能か把握するため,殻長と入水管口径の関係を求め た.アサリは,入水管から懸濁物粒子を取り込むが,成 育段階初期では入水管は形成されておらず,殻長1 mm 以上の稚貝に成長すると入水管が視認できる(千葉県 水産研究センター,2004).豊川河口域で採取した殻長 2‒6 mm の稚貝を対象に,実体顕微鏡下でデジタルカメ ラにより動画を撮影した.動画から,殻から入水管およ び出水管を出している瞬間を静止画として記録し,殻長 および入水管口径を測定した. 3. 結果 3.1 実験に供した試水中の植物プランクトン組成 実験に供した試水中の植物プランクトン種の現存量 を,細胞の等価球径のサイズ区分ごとに,採水日別,地 点別に Fig. 3 に示した.図は実験開始時における試験区 水槽3 個の平均値で示している.等価球径は1 細胞当た りの大きさであり,群体を形成する種ではアサリが実際
Fig. 3 The estimated carbon biomass of dominant phytoplankton species in natural seawater at several stations. Each column shows the aver-age of three experimental tanks
に摂取する大きさを示すものではない.
出現した植物プランクトンはいずれの場合も,浮遊性 の種類が全炭素現存量の90% 以上を占めた.
2017 年10 月31 日(Table 1, Exp. I)の 植 物 プ ラ ン ク トン炭素現存量は,いずれの地点,サイズ区分ともに 20 μgC·L−1以下と他の採水日と比べ少なかった.湾奥
の St.A ではサイズ区分10‒20 µm の炭素現存量が最も多 く,珪藻綱の Thalassiosira spp., Melosira varians, Navicula spp.が優占した.前二者は群体性種である.St.B および St.C では2‒10 µm の現存量が最も多く,群体性珪藻の Skeletonema costatum,単細胞性のクリプト藻綱(Cryp-tophyceae)が 優 占 し た. そ の ほ か 湾 央 部 の St.C で は 0.2‒2 µm でシアノバクテリアの Synechococcus sp.が多 かった.
2017 年11 月14 日(Table 1, Exp. II)で は, 湾 奥 か ら 沖に向かうに従い炭素現存量が多くなり,いずれの地 点 で も20 µm<が 最 も 多 く, 湾 央 部 の St.C の20 µm< では500 μgC·L−1以上と特に多かった.20 µm<の優占
種はいずれの地点でも群体性珪藻の Thalassiosira spp.,
Eucampia zodiacus であった.
2017 年11 月21 日(Table 1, Exp. III)では,炭素現存 量はいずれの地点,サイズ区分とも80 μgC·L−1以下
で,いずれのサイズ区分でも湾奥の St.A で他の地点と 比べ多かった.St.A では20 µm<が比較的多く,群体性 珪藻の Cerataulina pelagica, E. zodiacus や渦鞭毛藻綱の
Ceratium furca が優占した.St.A では次いで2‒10 µm お
よび10‒20 µm も多かった.2‒10 µm ではいずれの地点 ともに,単細胞性のクリプト藻綱(Cryptophyceae)およ びラフィド藻綱(Raphidophyceae)が優占した. 3.2 実験に供した試水中のサイズ分画クロロフィル a 実験に供した試水中のサイズ分画 Chl.a を,採水日 別,地点別に Fig. 4 に示した.
2017 年10 月31 日(Table 1, Exp. I)に採水した試水中 の Chl.a は,分画合計で1.3‒1.9 µg·L−1で,いずれの地 点,サイズ分画ともに1 µg·L−1以下と比較的少なかっ た.サイズ分画 Chl.a は,St.A および St.C では20 µm< で最も多かったが,植物プランクトン炭素現存量は St.A では10‒20 µm,St.C では2‒10 µm で最も多く,両者 の傾向は一致しなかった.これは植物プランクトンの うち,St.A の10‒20 µm では M. varians, Thalassiosira spp. が,St.C の2‒10 µm では S. costatum 等が比較的多く出現 し,これらは群体性の珪藻綱であるため結果的に粒子径 が大きくなったことによる.St.B では2‒10 µm で最も多 く,植物プランクトン現存量の出現傾向と一致した.
2017 年11 月14 日(Table 1, Exp. II)で は,Chl.a は 分 画合計で2.3‒15.1 µg·L−1で,湾奥から沖に向かうに従い
多く,いずれの地点ともに20 µm<で最も多い傾向がみ られ,植物プランクトン炭素現存量の出現傾向とよく 一致した.サイズ分画 Chl.a は,特に多かった St.C の 20 µm<の約12 µg·L−1を除き,約4 µg·L−1以下であった.
2017 年11 月21 日(Table 1, Exp. III)では,Chl.a は分 画合計で4.9‒8.2 µg·L−1で,ほとんどのサイズ分画とも に湾奥の St.A で他の地点と比べ多く,植物プランクト ン現存量の出現傾向とよく一致した.サイズ分画 Chl.a はいずれの地点ともに20 µm<が3 µg·L−1前後で最も多 く,次いで2‒10 µm で多かった. 3.3 実験に供した試水中の動物プランクトン 図には示さなかったが,実験に供した動物プランク トンの優占種は,繊毛虫門,節足動物門に属した.繊 毛虫門の優占種は,Exp. I では Ciliophora, Codonellopsis
morchella および Oligotrichida で,全 動物プランクトン
炭素現存量の4‒37% を占めた.Exp. II および III では,
Tiarina fusus, Amphorellopsis acuta および Oligotrichida が優
占し,全炭素現存量の2‒35%,Exp. III では29‒72% を占 めた.節足動物門では,Copepoda(nauplius), Paracalanus sp.(copepodite), Oithona spp.(copepodite)が優占し,Exp. I では全動物プランクトン炭素現存量の11‒90% を占 め,Exp. II で14‒88%,Exp. III で は11‒34% を 占 め た. 全動物プランクトン炭素現存量は,Exp. I では10.1‒ 17.8 mgC·m−3,Exp. II では6.6‒17.8 mgC·m−3,Exp. III で
は13.4‒35.8 mgC·m−3であり,各実験の水槽に投入され
たアサリの炭素現存量に対する動物プランクトンの割合 Fig. 4 The concentration of chlorophyll a on four size range.
はいずれの実験でも1% 未満であった. なお,動物プランクトンの炭素現存量では,実験前後 に一定の変化は認められなかった. 3.4 稚貝が摂取可能な粒子サイズの推定 殻長2 mm 以上の稚貝を対象とした測定値から,殻 長と入水管径の関係を Fig. 5 に示した.この関係式 から,アサリ稚貝が摂取可能な粒子サイズは,殻長 5 mm で<503 µm, 殻 長 10 mm で<839 µm, 殻 長 15 mm で<1170 µm と推定される. 3.5 実験により得られた Chl.a 変化量 実験前後の Chl.a 変化量を,サイズ分画ごとに各試験 区の平均値で示した(Fig. 6).グラフはアサリの殻長の 大きさ順に配置した.Chl.a 変化量の負の値が大きいほ ど,より Chl.a が減少したことを示している.
着底稚貝(Fig. 6a,Exp. II殻長217 µm,Exp. III 殻長 273 µm)のうち,殻長217 µm ではサイズ分画20 µm< の Chl.a が 大 き く 減 少 し た. 次 い で0.2‒2 µm お よ び 2‒10 µm で減少した.殻長273 µm では,2‒10 µm が大き く減少し,次いで0.2‒2 µm, 20 µm<が減少した.
初期稚貝では(Fig. 6a,Exp. III 殻長321 µm,Exp. I 殻 長441 µm,Exp. III 殻長481 µm),2‒10 µm が他の分画に 比べ大きく減少した.次いで大きく減少したのは,殻長 321 µm では0.2‒2 µm,殻長441 µm では10‒20 µm,殻長 481 µm では0.2‒2 µm および10‒20 µm であった.
稚 貝 で は(Fig. 6b,Exp. I 殻 長 5.7 mm,Exp. II 殻 長 9.3 mm,Exp. I 殻長10 mm,Exp. II 殻長15 mm,Exp. I 殻 長15 mm),すべての分画で Chl.a は減少し,稚貝サイズ が大きくなるにつれて変化量が大きくなる傾向がみられ た.
分画 Chl.a のうち最もサイズが大きい20 µm<の Chl.a が他の分画と比べ大きく減少したケースは Exp. II のみ で(Fig. 6a,Exp. II 殻 長 217 µm,Fig. 6b,Exp. II 殻 長 9.3 mm,Exp. II 殻長15 mm),それ以外の Exp. I および Exp. III では20 µm<の Chl.a 減少はそれほど大きくはな かった. 4. 考察 本研究では,実験前後のサイズ分画 Chl.a の変化量か ら,アサリ稚貝が摂取したと考えられる植物プランク トンの細胞サイズとその分類群を検討した.これには, 分画 Chl.a と植物プランクトンの細胞サイズとの間に 一定の関係が見られることが前提となる.今回は Chl.a と細胞サイズの関係を直接比較するデータを得なかっ た.そこで分画 Chl.a と,これと同じサイズ区分の等価 球径ごとの植物プランクトン炭素現存量との関係を調 べたところ,両者には統計的に有意な相関が認められ た(Fig. 7).したがって,サイズ分画 Chl.a の変化から アサリが摂取する植物プランクトンのサイズを検討して おおむね差し支えないと判断した.しかし,植物プラン クトンの細胞サイズから炭素現存量を推定する方法に は課題があることが指摘されており(Verity et al., 1992; Montagnes et al., 1994; Menden-Deuer & Lessard, 2000), Chl.a 量と植物プランクトンの炭素現存量との関係はさ まざまな要因で変化することが知られている(Furuya, 1990; Cloern et al., 1995).したがって,今回の実験のよ うにアサリが摂 したと考えられる天然海水中の植物プ ランクトンの種類とサイズを同時に測定する手法の精度 向上は今後の課題である. 実験後には実験前と比べ,実験水槽中の Chl.a が減少 した(Fig. 6).この減少の要因としては,植食者による 摂食が考えられる.植食者は,実験に供したアサリ稚貝 のほか,本実験では天然海水を用いたことから,動物プ ランクトンが挙げられる.動物プランクトンの炭素現存 Fig. 5 The relationship between shell length of juvenile clam (SL)
and the diameter of inhalant siphon (IS)
Fig. 6 Variation of size-fractionated chlorophyl a concentra-tion at different growth stages of Manila clams at several experiments. Error bar shows the standard deviation. The value on the x-axis shows the experimental number and the average shell length of Manila clam. a) settled juvenile and early juvenile, b) juvenile clam
量は実験の各水槽に投入したアサリ稚貝の現存量のいず れも1% 未満であり,動物プランクトンが Chl.a の減少 に及ぼす影響は極めて限定的であったと推測される.し たがって,実験後に Chl.a が減少した要因のほとんどは アサリ稚貝の摂食によるものと考えられた. 実験に用いた天然海水の全 Chl.a に占める各サイズ分 画の割合は,いずれの実験でも最も大きいサイズ分画の 20 µm<で最も多く,Exp. I では平均32%(15‒44%),Exp. II で 平 均68%(60‒80%),Exp. III で 平 均47%(36‒62%) を占め,特に Exp. II で割合が高く Chl.a 量も多かった (Fig. 4).しかしながら,Chl.a 変化量は Exp. II を除き
20 µm<に 比 べ2‒10 µm や10‒20 µm の 減 少 が 大 き か っ た(Fig. 6a, Exp. I, Exp. III).つまり,海水中には20 µm< の Chl.a 量が多いにもかかわらず,殻長273 µm および 441 µm のアサリが摂取する20 µm<の量は相対的に少な いことが伺える.Exp. II 殻長217 µm は,本研究で最も供 試個体の殻長が小さく,ここでみられた20 µm<の大きい サイズの Chl.a の減少は,アサリの摂食以外の別の要因 による可能性が考えられた. 本実験では,人工 料や培養した単一種の植物プラ ンクトンではなく六条潟でのアサリ稚貝の 料環境を 把握するため天然海水を用いたことから,さまざまな細 胞サイズの多種多様な植物プランクトンが含まれてい た(Fig. 3).前述のように,Exp. II および Exp. III では ともに20 µm<が多くを占めており,特に Exp. II で顕著 であった(Fig. 3).Exp. II における20 µm<の優占種は
Thalassiosira spp.(等価球径:約39 µm),E. zodiacus(等
価球径:約31 µm)で,いずれも群体性珪藻であった. Exp. III の20 µm<の優占種は同じく E. zodiacus の他,群 体性珪藻の Cerataulina pelagica(等価球径:約23 µm)で あった.植物プランクトンの細胞体積が5×103 µm3(等
価球径:約20 µm)を超える種類では,沈降速度は種類 に も よ る が1.0 m·d−1以 上 に な り(Bienfang & Harrison,
1984; Smayda, 1970),他の小型種と比べ沈降速度が大き いと考えられる.実験中は,着底稚貝等の巻き上がりを 防ぐためエアレーションを微弱にした.そのため,細 胞のサイズが比較的大型で群体を形成する珪藻が実験 水槽中で沈降し,見かけ上 Chl.a 量が減少した可能性が 考えられる.実際,Exp. II では,実験終了時に実験水 槽の底に植物プランクトン等と思われる懸濁物が沈殿 している様子が観察された.Exp. III 殻長321 µm では 20 µm<が減少していないことを考慮すると,Exp. II 殻 長217 µm および Exp. III 殻長273 µm での20 µm<の減少 は沈降によるものであり,アサリ稚貝に摂取されたもの ではないと判断するのが妥当であろう.したがって,殻 長300 µm までの着底稚貝は10 µm 未満の植物プランク トンを摂 すると考えられる.実験中の植物プランクト ン細胞の沈降による結果への影響の除去は今後の課題で ある. 伯耆(2016)は,着底稚貝および初期稚貝(殻長252‒ 572 µm)に対する4 種類の珪藻(Ch. gracilis, Navicula sp.,
Ch. affinis, Cylindrotheca closterium)の培養株の 料価値
を調べ,最も成長速度が高く好適な種類は Ch. gracilis であったと報告している.これら珪藻のサイズは明示さ れていないが,Ch. gracilis の細胞の大きさは4‒8 µm と 考えられ(千葉県水産研究センター,2004),本研究の 着底稚貝(殻長200‒300 µm)で2‒10 µm の Chl.a が他の 分画に比べ大きく減少した結果とよく一致した. 一方,稚貝(殻長5.7‒15 mm)では,すべての分画で Chl.a が減少した(Fig. 6b).推定された入水管径(Fig. 5) から判断して,これらの稚貝は0.2 µm から20 µm<のす べてのサイズを摂取可能であったと考えられる. 以上の分画 Chl.a の変化および植物プランクトン種組 成から判断した,アサリが摂取可能な植物プランクト ンのサイズと,それに対応した植物プランクトン藻類群 を Fig. 8 に整理した.プランクトンのサイズカテゴリー は Sieburth et al.(1978)に準じ,本実験で用いた天然海 水中に含まれる植物プランクトン藻類群のサイズ範囲 をそれぞれ示した.加えて,アサリの各成長段階にお ける摂取可能な 料サイズの範囲を示した.着底稚貝 (殻長200‒300 µm)が摂取可能な 料サイズは0.2‒2 µm と2‒10 µm で,これらのサイズの植物プランクトン分 類群は,0.2‒2 µm はシアノバクテリア,2‒10 µm はク リプト藻綱,ラフィド藻綱等の微小鞭毛藻類が該当し た.初期稚貝(殻長300‒1000 µm)のうち殻長500 µm 程 度までは,0.2‒2 µm および2‒10 µm に10‒20 µm が加わ り,これら微小鞭毛藻類に加え珪藻綱が含まれる.殻長 500 µm より大きい初期稚貝と稚貝では,上記サイズに 加え20 µm<が摂取可能となり,これに該当する植物プ ランクトンは,珪藻綱と渦鞭毛藻綱である.このほか, 今回は優占しなかったが,一般には20 µm 未満には渦鞭 毛藻綱が含まれる.ここでは 料のサイズ下限値を明ら かにすることはできなかったが,アサリ浮遊幼生を対象 に天然海水中の懸濁粒子の取り込み効率を調べた例で は,1.4‒2.0 µm のサイズの率が高かったという報告があ る(Tezuka et al., 2009). 一般に微細藻類は二枚貝のすべての生育段階の食物と Fig. 7 The relationship between the size-fractionated chlorophyll
a concentration and the estimated phytoplankton biomass
して重要で(Brown et al., 1989),栄養価の指標として藻 類に含まれる多価不飽和脂肪酸が着目されている(Volk-man et al., 1989; 鳥羽ら,1994; Caers et al., 1999; Delaporte
et al., 2005; 丸尾ら,2012 など).六条潟においては着底
稚貝および初期稚貝が摂取し比較的現存量が多くサイ ズが20 µm 以下の藻類は,シアノバクテリア,クリプト 藻綱,珪藻綱,ラフィド藻綱に属する種類であった.こ れらのうちシアノバクテリアは多価不飽和脂肪酸を含 まず(齋藤,1996; Patil et al., 2007; Jónasdóttir, 2019),淡 水性のシアノバクテリアである Synechocystis spp.はヤマ トシジミの 料として不適であるとの指摘があり(嵯峨 ら,2014),アサリにとってもシアノバクテリアは 料 価値が低い可能性がある.シアノバクテリア以外のクリ プト藻綱,珪藻綱,ラフィド藻綱に属する種では,ω-3 不飽和脂肪酸が比較的豊富に含まれる(Patil et al., 2007; Mitani et al., 2017; Jónasdóttir, 2019 な ど ). 六 条 潟 周 辺 海域では豊川の流量が増大した際に20 µm 以下の Chl.a が多く,クリプト藻綱の組成比が大きくなるという森 ら(2020)の報告がある.六条潟のアサリ稚貝はこれら 微細藻類を有効に利用しており,微細藻類の発生が六条 潟でのアサリ稚貝の生残,成長を可能にする重要な条件 のひとつになっていると考えられる. 本研究では,アサリ稚貝が摂 する 料を把握するた め,六条潟周辺海域において採水された海水中に一定 時間着底稚貝等を投入し,実験前後の差から 料のサイ ズと植物プランクトン種を調べた.得られた Chl.a 変化 量の結果には,ばらつきが見られた(Fig. 6).その要因 としては,採取地点による Chl.a 量の多少,出現した植 物プランクトンの種とサイズの相違が挙げられる.任意 の植物プランクトン現存量を与えることが可能な培養さ れた 料と異なり,天然海水では実験開始時の Chl.a 量 が少ない場合は実験前後で差が検出されにくい場合が あり,また多種多様な植物プランクトンが対象となるた め,得られる結果の分散が大きくなることがあると考え られる.今後これらの課題を回避するためには,実験を 実施する試験区数や各試験区での試料数を増やすなど, 分散を低減する工夫が必要である. アサリに関する最近の研究によると,三河湾では 秋季の 料不足がその後の減耗を招いており(蒲原 ら,2018),秋季から冬季の減耗を乗り越えるためには, アサリ個体群の維持に必要な 料環境の解明が重要と指 摘されている(曽根ら,2019).芝ら(2020)は,アサリ の個体成長モデルを用いた解析の結果,近年の伊勢湾東 部沿岸域におけるアサリの減耗は, 料環境の悪化が関 与している可能性があることから,アサリのエネルギー 収支において同化量を増やす 料環境の改善が必要と指 摘している.本研究により,着底初期のアサリの 料と して10 µm 未満の植物プランクトンが重要であると考え られたことから,今後は 料のサイズを考慮して 料環 境を把握するとともに,数値モデルによりアサリ稚貝の 生残,成長を定量的に解析する際にはアサリ稚貝,植物 プランクトンともにサイズを考慮したモデル化が求めら れる. 謝 辞 本研究成果は,国土交通省中部地方整備局三河港湾事 務所により三河湾で実施された調査の結果の一部であ り,有益な指導・助言をいただいた大阪大学大学院工学 研究科青木伸一教授,名城大学大学院総合学術研究科鈴 木輝明特任教授,横浜国立大学大学院都市イノベーショ ン学府・研究院中村由行教授に深く感謝申し上げます. 実験結果の解釈にあたっては,三重大学大学院生物資源 学研究科伯耆匠二助教に有益な助言をいただきました. 深く感謝申し上げます.最後に,調査にご協力いただい Fig. 8 Available food size for several growth stages of Manila clams and phytoplankton groups. Size classification of phytoplankton based
on Sieburth et al. (1978). For phytoplankton, the size range of phytoplankton algae contained in the natural seawater used in this experiment was shown
た愛知県漁業協同組合連合会の皆様に厚く御礼申し上げ ます.
参考文献
Arakawa, H., Yaoita, T., Koike, T. & T. Morinaga, 1997. Size of suspended particles caught by Manila clam, Ruditapes
philippinarum. Mer (Paris), 35, 149‒156.
Bienfang, P. K. & P. J. Harrison, 1984. Co-variation of sinking rate and cell quota among nutrient replete marine phytoplankton. Mar. Ecol. Prog. Ser., 14, 297‒300.
Brown, M. R., Jeffrey, S. W. & C. D. Garland, 1989. Nutritional aspects of microalgae used in magniculture; a literature review. CSIRO Mar. Lab. Rep., 205, 44 pp.
Caers, M., Coutteau, P. & P. Sorgeloos, 1999. Dietary impact of al-gal and artificial diets, fed at different feeding rations, on the growth and fatty acid composition of Tapes philippinarum (L.) spat. Aquaculture, 170, 307‒322.
千葉県水産研究センター,2004.アサリ種苗生産の現場基 礎技術̶富津研究所の経験̶.千葉県水産研究セン ター,千葉県.98 pp.
Cloern, J. E., Grenz, C. & L. Vidergar-Lucas, 1995. An empirical model of the phytoplankton chlorophyll: Carbon ratio̶The conversion factor between productivity and growth rate. Limnol. Oceanogr., 40, 1313‒1321.
Delaporte, M., Soudant, P., Moal, J., Kraffe, E., Marty, Y. & J. Sa-main, 2005. Incorporation and modification of dietary fatty acids in gill polar lipids by two bivalve species Crassostrea
gigas and Ruditapes philippinarum. Comp. Biochem.
Physi-ol. A MPhysi-ol. Integr. PhysiPhysi-ol., 140, 460‒470.
Furuya, K., 1990. Subsurface chlorophyll maximum in the tropical and subtropical western Pacific Ocean: Vertical profiles of phytoplankton biomass and its relationship with chlorophyll
a and particle organic carbon. Mar. Biol., 107, 529‒539.
弘田禮一郎,1986.動物プランクトン調査:現存量の測定. 「沿岸環境調査マニュアル(底質・生物編)」(日本海洋 学会編).東京:恒星社厚生閣,pp. 184‒191. 伯耆匠二,2016.アサリの摂 生態と消化機構に関する研究. 博士論文,東京大学. 市川哲也,桃井幹夫,鈴木輝明,石田基雄,2010.リセプ ターモードモデルを利用した造成適地選定手法.水産 工学,46, 217‒222.
Jónasdóttir, S., 2019. Fatty acid profiles and production in marine phytoplankton. Mar. Drugs, 17, 151.
蒲原聡,芝修一,市川哲也,鈴木輝明,2018.伊勢・三河湾 のアサリ増殖環境.月刊海洋,50, 406‒414. 蒲原聡,山田智,曽根亮太,青木伸一,2014.三河湾六条潟 におけるアサリ稚貝の生産機構.愛知県水産試験場研 究報告,19, 1‒9. 蒲原聡,山田智,和久光靖,曽根亮太,岩田靖宏,2013.三 河湾六条潟におけるアサリ着底初期稚貝の動態.愛知 県水産試験場研究報告,18, 13‒20.
Kana, T. & P. M. Glibert, 1987. Effect of irradiances up to 2000 μEm−2 s−1 on marine Synechococcus WH7803-II.
Pho-tosynthetic responses and mechanisms. Deep-Sea Res., 34, 497‒516.
丸尾知佳子,藤林恵,愛川良雄,西村修,2012.イソシジ ミ(Nuttallina olivacea)の生育状況に影響を与える 料源の検討.土木学会論文集 G(環境),68, III_277‒
III_283.
Menden-Deuer, S. & E. J. Lessard, 2000. Carbon to volume relation-ships for dinoflagellates, diatoms, and other protist plankton. Limnol. Oceanogr., 45, 569‒579.
Mitani, E., Nakayama, F., Matsuwaki, I., Ichi, I., Kawabata, A., Kawachi, M. & M. Kato, 2017. Fatty acid composition pro-files of 235 strains of three microalgal divisions within the NIES microbial culture collection. Microb. Resour. Syst., 33, 19‒29.
宮井博,松崎加奈恵,小川数也,杉原拓郎,1988.植物プラ ンクトンの形態に基づく種類別生物量の簡易推定法. 日本プランクトン学会報,35, 121‒126.
Montagnes, D. J. S., Berges, J. A., Harrison, P. J. & F. J. R. Taylor, 1994. Estimating carbon, nitrogen, protein, and chlorophyll
a from volume in marine phytoplankton. Limnol.
Ocean-ogr., 39, 1044‒1060.
森将人,今尾和正,古賀あかね,橋口晴穂,中田喜三郎, 2020.三河湾六条潟周辺海域におけるアサリ Ruditapes
philippinarum 稚貝の 料環境.J. Adv. Mar. Sci. Tech.
Soci., 26, 11‒23.
Nakamura, Y., 1998. Growth and grazing of a large heterotrophic dinoflagellate Noctiluca scintillans laboratory cultures. J. Plankton Res., 20, 1711‒1720.
Nakamura, Y., 2004. Suspension feeding and growth of juvenile Manila clam Ruditapes philippinarum reared in the labora-tory. Fish. Sci., 70, 215‒222.
Patil, V., Kallqvist, T., Olsen, E., Vogt, G. & H. R. Gislerod, 2007. Fatty acid composition of 12 microalgae for possible use in aquaculture feed. Aquacult. Int., 15, 1‒9.
嵯峨友樹,宮廻隆洋,小山維尊,野尻由香里,神谷宏,勢村 均,谷幸則,2014.宍道湖における植物プランクトン の含有するカロテノイドとω-3 不飽和脂肪酸の測定. 陸水学会要旨集,107. 齋藤洋昭,1996.海洋生物と n-3 高度不飽和脂肪酸.化学と 生物,34, 107‒113. 芝修一,姫野天領,吉田司,蒲原聡,田中義人,鈴木輝明, 2020.個体成長モデルを用いた伊勢湾東部沿岸域にお けるアサリ Ruditapes philippinarum 資源の減耗要因の 検討.水産海洋研究,84, 11‒26.
Sieburth, J. M., Smetacek, V. & J. Lenz, 1978. Pelagic ecosystem structure: Heterotrophic compartments of the plankton and their relationship to plankton size fractions. Limnol. Ocean-ogr., 23, 1256‒1263.
Smayda, T. J., 1970. The suspensions and sinking of phytoplankton in the sea. Oceanogr. Mar. Biol. Rev., 8, 353‒414.
曽根亮太,蒲原聡,山田智,鈴木輝明,高倍昭洋,2015.ア サリ Ruditapes philippinarum 稚貝が高密度に発生する 河口干潟における水質浄化機能̶三河湾豊川河口六 条潟における6 月の観測事例̶.水産海洋研究,79, 117‒129. 曽根亮太,和久光靖,石田俊朗,宮脇大,山田智,2019.六 条潟におけるアサリ Ruditapes philippinarum の秋季減 耗要因について.水産海洋研究,83, 252‒259.
Strathmann, R. R., 1967. Estimating the organic carbon content of phytoplankton from cell volume or plasma volume. Limnol. Oceanogr., 12, 411‒418.
Strickland, J. D. H. & T. R. Parsons, 1968. A practical handbook of seawater analysis. Fish Res. Board Can., 167, 185‒194. 鈴木輝明,2017.沿岸環境の再生・創出と豊かな漁業生産
̶伊勢・三河湾を例として̶.海洋と生物,39, 554‒ 563. 社団法人全国沿岸漁業振興開発協会編,1997.第3 編アサリ 増殖場造成計画.沿岸漁場整備開発事業,増殖場造成 計画指針ヒラメ・アサリ編,平成8 年度版.東京:社 団法人全国沿岸漁業振興開発協会,pp. 123‒316. Tezuka, N., Ichisaki, E., Kanematsu, M., Usuki, H., Hamaguchi, M.
& K. Iseki, 2009. Particle retention efficiency of asari clam
Ruditapes philippinarum larvae. Aquat. Biol., 6, 281‒287.
鳥羽光晴,深山義文,酒井美恵,1994.イソクリシス・タヒ チ株の大量培養̶IV 単一種藻類給 でのアサリ稚貝
に対する 料価値.栽培技研,22, 75‒81.
Verity, P. G., Robertson, C. Y., Tronzo, C. R., Andrews, M. G., Nelson, J. R. & M. E. Sieracki, 1992. Relationships between cell volume and the carbon and nitrogen content of marine photosynthetic nanoplankton. Limnol. Oceanogr., 37, 1434‒ 1446.
Volkman, J. K., Jeffry, S. W., Nichols, P. D., Rogers, G. I. & C. D. Garland, 1989. Fatty acid and lipid composition of 10 spe-cies of microalgae used in mariculture. J. Exp. Mar. Biol. Ecol., 128, 219‒240.
* * * * * * *
Phytoplankton Size and Its Algal Group in Natural Seawater Consumed by Juvenile
Manila Clam Ruditapes philippinarum in Rokujo Tidal Flat of Mikawa Bay, Japan
Kiyo Kurita*1, Kazumasa Imao*1, Seiho Hashiguchi*1, Masato Mori*1, Kisaburo Nakata*2
*1 Chubu Branch, Marine Biological Research Institute of Japan Co., Ltd., 3‒9‒2 Koubai-cho, Showa-ku, Nagoya, Aichi 466‒0031, Japan *2 Graduate School of Environmental and Human Sciences, Meijo University, 1‒501 Shiogamaguchi, Tenpaku-ku, Nagoya, Aichi
468‒8502, Japan