The role of autophagy in Arabidopsis plants
exposed to ultraviolet-B or high temperature
stress
著者
Gonul Dundar
号
18
学位授与機関
Tohoku University
学位授与番号
生博第413号
URL
http://hdl.handle.net/10097/00129521
氏 名 ( 本 籍 地 )
学 位 の 種 類
学 位 記 番 号
学 位 授 与 年 月 日
学 位 授 与 の 要 件
研 究 科 , 専 攻
論
文
題
目
博士論文審査委員
グニュル ドュンダルGonul Dundar
博士(生命科学)
生博第 413 号
令和2年9月25日
学位規則第4条第1項該当
東北大学大学院生命科学研究科
(博士課程)生態システム生命科学専攻
The role of autophagy in Arabidopsis plants exposed to
ultraviolet-B or high temperature stress (UVB または高
温ストレスに曝されたシロイヌナズナ植物におけるオート
ファジーの役割 )
(主査)准教授 日出間 純
教 授 東谷 篤志
教 授 佐藤 修正
教 授 永田 裕二
論文内容の要旨
Autophagy is one of the catabolic processes important for the survival of the cells by ensuring homeostasis under various conditions. Its functions are known both in a bulk degradation process where cytoplasmic components are unselectively degraded for nutrient recycling and in a selective degradation for the elimination of damaged organelles by vacuolar hydrolyses enzymes. Autophagy plays a central role in programmed cell death (PCD), which is an essential feature of normal plant development, and also in some of the environmental stresses. Recently, there are few reports related to autophagy role in the mechanisms of ultraviolet-B (UV-B) or high temperature (HT) tolerance. However, the knowledge about its role in UV-B responses is not known in plants and even controversy in the case of HT stress.
Chapter I In addition to the substantial damage to plant macromolecules,1 UV-B induces lesions in nuclear, mitochondrial, and chloroplast genome by specifically inducing photoproducts between adjacent pyrimidines on the same DNA strand in plants.2 The cyclobutane pyrimidine dimers (CPDs), which make up the largest component (~75%) of photoproducts,4 can retard growth and development in
Arabidopsis.3,4 The CPDs, are the principal cause of UV-B-induced growth inhibition in plants, thus often used as a direct indicator of UV-B damage.5, Photoreactivation by CPD photolyase is the primary mechanism of CPD repair in rice plants5 and has previously demonstrated that CPD photolyase is crucial for determining the resistance of rice to UV-B, and that elevated CPD photolyase activity can significantly alleviate UV-B-induced growth inhibition.5,7 In Arabidopsis, deficiency of CPD photolyase (PHR1/ UV RESISTANCE 2 [UVR2]; AT1G12370) induces UV-B sensitivity,3 indicating that UV-B-induced CPD accumulation is a principal cause of UV-B-induced cell death and growth inhibition in plants. Additionally, previous study that mutants of the autophagy-related genes (ATG2, 5, 7) exhibited UV-B-sensitive phenotypes like that of PHR1-deficient Arabidopsis thaliana mutants (uvr2).8 These results indicated that autophagy also plays an important role in the plant response to UV-B-induced damage.
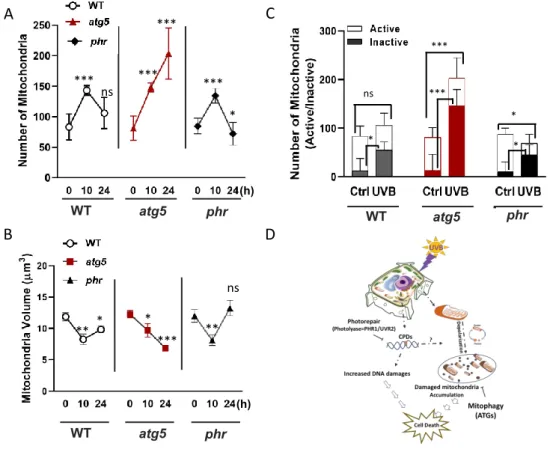
In yeast and mammals, damaged organelles, especially mitochondria are removed by a selective autophagic process known as mitophagy.9 Mitophagy is linked to several physiological functions and human pathologies, such as neurodegenerative disease.10 However, little is known about the relationships between UV-B-induced CPD accumulation and mitochondrial dynamics or between mitochondrial dynamics and autophagy in UV-B damaged plants. Here, we analysed the mitochondrial dynamics and UV-B-induced CPD accumulation in WT, atg5, phr, atg5 phr, and AtPHRox plants in order to understand the role of mitochondria removal by autophagy in UV-B protection in plant cells. The severe UV-B-sensitive phenotype of the atg5 phr double mutant compared with the atg5 and phr single mutants, which were already more sensitive than WT, strongly suggest that observed phenotype is resulted from deficiency in two independent machineries in response to UV-B, i.e., autophagy and CPD photorepair. Furthermore, UV-B induced CPD level was similar between WT and atg5 plants indicating UV-B hypersensitivity of atg5 is not due to high CPDs accumulation. Here, I found that mitochondria, one of the reproductive organelle having its own genome, were fragmented and the number was significantly increased at 10 h after UV-B irradiation in all genotypes, but decreased in WT and phr1
plants at 24 h after UV-B irradiation while massively accumulated in atg mutants. In order to analyse activity of these accumulated mitochondria, tetramethylrhodamine, ethyl ester (TMRE) staining method was preformed and revealed that higher portion of the remaining mitochondria in UV-B treated plants were dysfunctional. Southern blot results for checking CPD levels on mtDNA were no different between WT and atg5 plants indicating that UV-B-induced mitochondrial fragmentation occurred in regardless of CPD accumulation on mtDNA. Taken together here we show that UV-B damages mitochondrial function and causes mitochondrial fragmentation in Arabidopsis, and the damaged dysfunctional mitochondria are removed by autophagy.
(h) ns *** *** *** * *** WT atg5 phr ** ** *** * * ns WT atg5 phr (h) WT atg5 phr * * *** ns *** * WT atg5 phr A B C D
Figure 1 Changes in number and volume of mitochondria in autophagy- and photorepair-deficient Arabidopsis mutants
exposed to UV-B. (A) and (B) Number and the volume of each mitochondria is counted with IMARIS software by MT-GFP dots quantification within each cell. Images from three different replicates are used for calculation (n ≥ 40 cells for mitochondria number, n ≥ 600 mitochondria for volume) ± SE. (C) Ratio of the active mitochondria are calculated by dividing TMRE-signal dots number to the GFP-dots number within each cell. Images from three different replicates are used for calculation (n ≥ 20) ± SE. Error bars denote standard deviation and asterisk indicates significant difference (P < 0.05) according to the two‐way ANOVA analysis. (D) Graphical figure of UV-B induced mitophagy.
Chapter II Microsporogenesis is one of the most sensitive process affected by HT.11 Anther development follows well-regulated programs and amidst them PCD is crucial to break down anther wall cells, such as tapetum and middle layer cells for maturing pollen grains and anther dehiscence. The proper timing of tapetal degradation by PCD is necessary for the production of viable pollen.11 MYB80 is a key transcription factor that controls tapetal PCD by regulating several genes, including directly upregulating UNDEAD.12 Autophagy is thought dispensable for regular reproductive development in
Arabidopsis,13 intriguingly, a transgenic tobacco line overexpressing Arabidopsis AtATG6/BECLIN1 in the tapetum cells showed male sterility14 due to accelerated tapetum cell death therefore microsporogenesis abortion. Nevertheless, the rice autophagy null mutant OsATG7 showed a defect in tapetum cell degradation and sporophytic male sterility under normal culture conditions.15 These results indicate autophagy plays an important role in the tapetal degeneration, but the biological significance and its specific function in development are still controversial. Here, to check whether elevated temperatures directly induce autophagy, and if so, how autophagy is involved in upon HT stress, the contribution of autophagy to pollen and anther development under moderate HT stress is analysed in
Arabidopsis plants.
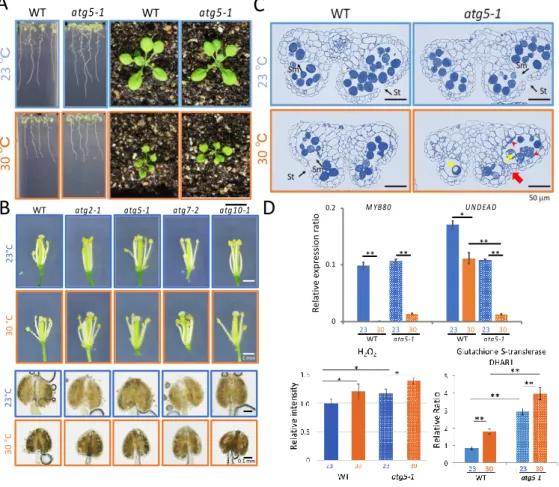
Firstly, the physiologic parameters of plants grown at 30 °C were measured at different developmental stages in WT and atg mutant plants and found that most of the atg plants showed no difference than WT plants at the vegetative stage but in the reproductive stage. Furthermore, Arabidopsis atg mutants became almost completely male-sterile at 30 ℃ whilst WT plants still set some seeds indicating that autophagy is essential to for mitigating HT injury during pollen development. Meanwhile, ovules developed normal in both WT and atg5 under either temperature conditions. In the histologic observation of the anthers cross-sections of stage 13 anthers at 30 °C, the pollen size and number decreased in WT anthers, but there was still septa and stomium breakage. Whilst, septum and stomium breakage did not occur in atg5 anthers. These results indicate that autophagy is essential for septum and stomium breakage at 30 °C, that is necessary for the maturation and release of the pollens through degeneration of anther wall cells. Gene expression results of MYB80 and UNDEAD indicated that HT stress alters the timing of tapetal PCD through repression of MYB80 transcriptional regulation, which then sequentially activates autophagy to compensate reduction of tapetal PCD by HT. In addition to this hypothesis, here I also found that HT induces oxidative damage in developing anthers, much more prominently in the atg5 mutant. Altogether, in addition to HT-induced ROS, over-accumulation of ROS in autophagy deficiency may strongly affect the spatiotemporal ROS signalling for tapetal PCD. In either hypothesis, the proper timing of pollen development including tapetal PCD is strictly controlled, and autophagy seems to be important to minimize the effects of HT and to maintain this pollen specific development.
atg2-1 atg5-1 atg7-2 atg10-1 WT 1 mm 0.1 mm 2 3° C 3 0 ° C 2 3 °C 3 0 ° C 2 3 °C 3 0 ° C
atg2-1 atg5-1 atg7-2 atg10-1
WT 1 mm 0.1 mm 2 3° C 3 0 ° C 2 3 °C 3 0 ° C 2 3 °C 3 0 ° C W T 23 a tg 5 -1 23 30 (10 days) 23 (30 days) 30 (22 days) 30 (4 days) 35
A
B
C
W T 23 a tg 5-1 23 30 (10 days) 23 (30 days) 30 (22 days) 30 (4 days) 35 A B C W T 23 at g5 -1 23 30 (10 days) 23 (30 days) 30 (22 days) 30 (4 days) 35 A B C W T 23 a tg 5-1 23 30 (10 days) 23 (30 days) 30 (22 days) 30 (4 days) 35A
WT atg5-1B
C
3 0 ℃ 2 3 ℃ WT atg5-1 0 0.1 0.2 MYB80 UNDEAD R e la ti ve e xp re ss io n r at io 23 30 23 30 WT atg5-1 23 30 23 30 WT atg5-1 M YB80 UN DEAD ** ** ** * ** 3 0 ℃ 2 3 ℃A
C
B
D
Figure 2 Autophagy contribution on HT responses. (A) Ten day-old seedlings of WT Col-0 and atg5 mutant were grown at
23 °C (blue), 30 °C (orange) on soil. (B) Pistil and anther morphology at flowering stage at 23 and 30 °C. Scale bars (SB), 1 mm. Iodine staining of pollen grains (down panel) in anthers. SB, 0.1 mm. (C) Horizontal cross-section of anther stained with 0.05% toluidine blue. Red arrows and T show undegenerated tapetum cells in atg5 mutant at 30 ℃. SB, 100 µm. (D) Hydrogen peroxide levels were detected by DAB staining at stage 12 anthers. GST/DHAR1 protein from MALDI-TOF analysis. **P<0.01. Altered genes expression of MYB80 and UNDEAD in stage 9 anthers of WT and atg5 mutant grown at 23 or 30 ℃ for 3 days. Bars show ±SD. * P<0.05. **P<0.01.
Conclusion In this study, I showed that autophagy plays an important role in both UV-B and HT
responses thorough involving in the cell death process, and therefore it can be used as a modulator mechanism to alleviate the decrease in plant growth and yield. atg mutants of Arabidopsis exhibited UV-B-hypersensitive phenotypes during the vegetative stage, whilst HT of 30 ℃ resulted in complete abortion of pollen development during the developmental stage. These results suggest that autophagy exerts an essential role in the cell death pathway in plants exposed to UV-B and HT, and the degree of the importance for cells varies depending on the type of stress and growth stages during stress exposure.
Bibliography
1. Tevini, M. et al. Effects of UV-B Radiation on Plants During Mild Water Stress II. Effects on Growth, Protein and Flavonoid Content. Zeitschrift für Pflanzenphysiologie 110, 459–467 (1983).
2. Britt, A. B. et al. UV-sensitive mutant of Arabidopsis defective in the repair of pyrimidine-pyrimidinone(6-4) dimers. Science 261, 1571–4 (1993).
3. Landry, L. G. et al. An Arabidopsis photolyase mutant is hypersensitive to ultraviolet-B radiation.
Proc. Natl. Acad. Sci. U. S. A. 94, 328–32 (1997).
4. Mitchell, D. L. et al. The biology of the (6-4) photoproduct. Photochemistry and photobiology 49, 805–19 (1989).
5. Hidema, J. et al. Increase in CPD photolyase activity functions effectively to prevent growth inhibition caused by UVB radiation. Plant J. 50, 70–9 (2007).
6. Takahashi, M. et al. Cyclobutane pyrimidine dimer (CPD) photolyase repairs ultraviolet-B-induced CPDs in rice chloroplast and mitochondrial DNA. Plant J. 66, 433–42 (2011).
7. Hidema, J. et al. Spontaneously occurring mutations in the cyclobutane pyrimidine dimer photolyase gene cause different sensitivities to ultraviolet-B in rice. Plant J. 43, 57–67 (2005). 8. Izumi, M. et al. Entire Photodamaged Chloroplasts Are Transported to the Central Vacuole by
Autophagy. Plant Cell tpc.00637.2016 (2017). doi:10.1105/tpc.16.00637
9. Youle, R. J. & Narendra, D. P. Mechanisms of mitophagy. Nat. Rev. Mol. Cell Biol. 12, 9–14 (2011).
10. Kanki et al. Mitophagy in yeast: Molecular mechanisms and physiological role. Biochimica et
Biophysica Acta - Molecular Cell Research 1853, 2756–65 (2015).
11. Varnier, A. et al. Programmed cell death progressively models the development of anther sporophytic tissues from the tapetum and is triggered in pollen grains during. Elsevier (2005) 12. Anh Phan, H. et al. The MYB80 Transcription Factor Is Required for Pollen Development and
the Regulation of Tapetal Programmed Cell Death in Arabidopsis thaliana W OA. The Plant Cell,
Vol. 23: 2209–24, (2011)
13. Yoshimoto K.et al. Beginning to understand autophagy, an intracellular self-degradation system in plants.Plant and Cell Physiology, 1355–65 (2012)
14. Singh, S. P. et al. BECLIN1 from Arabidopsis thaliana under the generic control of regulated expression systems, a strategy for developing male sterile plants. Plant Biotechnol. J. 8, 1005–22 (2010).
15. Hanamata, S.et al. Roles of autophagy in male reproductive development in plants. Front. Plant
論文審査結果の要旨 老化、飢餓の間に細胞成分を分解する細胞プロセスの 1 つであるオートファジーは、植物の環境ストレス に対する抵抗性の獲得に重要な役割を演じていることが、近年様々なストレス耐性とオートファジーに関す る研究から示唆されている。当研究室においても、高温(HT)ストレス時の植物の花粉発生障害や紫外線 UVB 防御に対してオートファジー機能が関わっている可能性を示唆する結果を得ている。しかし、それら の現象が具体的にどのような局面においてオートファジーが関与し、ストレス耐性に関わっているかなどは 不明であった。Gonul Dundar さんは、オートファジーの一連のプロセスに関わる遺伝子を欠失し、オートフ ァジー機能を失った種々の変異体(atg 変異体)を材料に、以下の点を明らかにした。 (1) 高温(HT)ストレスに対する植物の花粉発生の感受性はよく知られていますが、HT 傷害におけるオート ファジーの関与は不明であった。そこで高温障害を引き起こす30℃でシロイヌナズナの野生株と atg 変 異体を栽培すると、atg 変異体は野生株と比較して明らかに花粉の発達障害と葯の裂開障害が顕著で あった。その要因に関してさらに解析を進めた結果、 通常は花粉の発達に重要なタペート細胞のプロ グラム細胞死が、HT 条件下で抑制される。そのような条件下ではオートファジーが誘導され、タペート 細胞の退化、そして葯の発達に重要な役割を演じていることを見出した。 (2) atg 変異体が UVB 感受性の表現型を示す。その要因を探るために、ミトコンドリア動態とオートファジー に着目し解析を行ったところ、UVB はミトコンドリアの断片化を誘導し、断片化され不活化されたミトコン ドリアは、選択的にオートファジー(マイトファジー)により除去されることを植物細胞を用いて初めて見 出し、この障害を受けたミトコンドリアのマイトファジーの欠失が、UVB 感受性の主要因であることを示し た。また、この UVB 障害時におけるオートファジーは、DNA 損傷による障害応答経路とは独立した経 路で活性化されることも示した。 これらの研究成果は、植物のストレス防御におけるオートファジーの具体的な局面での重要性を実 証したものであり、今後のストレス耐性育種に向けた重要な知見を提供できたと評価する。 本研究成果は、自立して研究活動を行うに必要な高度の研究能力と学識を有することを示している。した がって、Gonul Dundar 女史提出の論文は、博士(生命科学)の博士論文として合格と認める。