つくばリポジトリ NC 9 162
全文
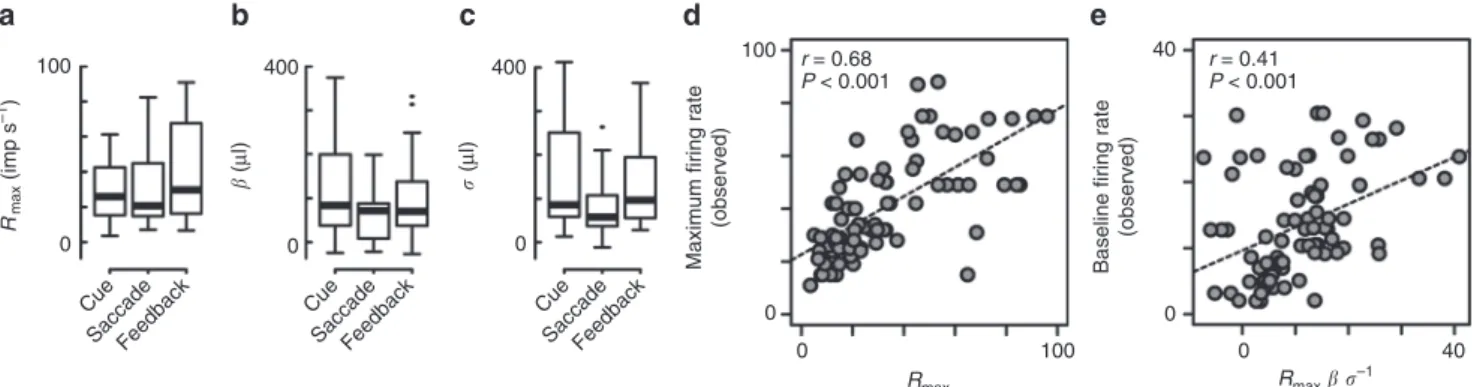
図
関連したドキュメント
An easy-to-use procedure is presented for improving the ε-constraint method for computing the efficient frontier of the portfolio selection problem endowed with additional cardinality
It is suggested by our method that most of the quadratic algebras for all St¨ ackel equivalence classes of 3D second order quantum superintegrable systems on conformally flat
Keywords: continuous time random walk, Brownian motion, collision time, skew Young tableaux, tandem queue.. AMS 2000 Subject Classification: Primary:
We include applications to elliptic operators with Dirichlet, Neumann or Robin type boundary conditions on L p -spaces and on the space of continuous
n , 1) maps the space of all homogeneous elements of degree n of an arbitrary free associative algebra onto its subspace of homogeneous Lie elements of degree n. A second
In this work, we present a new model of thermo-electro-viscoelasticity, we prove the existence and uniqueness of the solution of contact problem with Tresca’s friction law by
the existence of a weak solution for the problem for a viscoelastic material with regularized contact stress and constant friction coefficient has been established, using the
Abstract: The existence and uniqueness of local and global solutions for the Kirchhoff–Carrier nonlinear model for the vibrations of elastic strings in noncylindrical domains
