The role of calcium response in regulation and maintenance of secretory function in salivary glands
(唾液腺の分泌機能の調節と維持におけるカルシウム 応答の役割)
日本大学松戸歯学研究科細胞機能制御学専攻 櫻井 甫
(指導 吉垣純子)
全文
(唾液腺の分泌機能の調節と維持におけるカルシウム 応答の役割)
日本大学松戸歯学研究科細胞機能制御学専攻 櫻井 甫
(指導 吉垣純子)
図
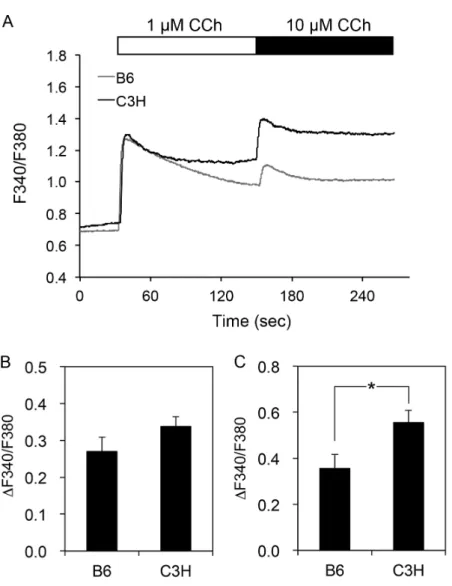
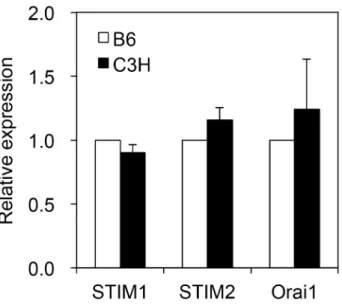
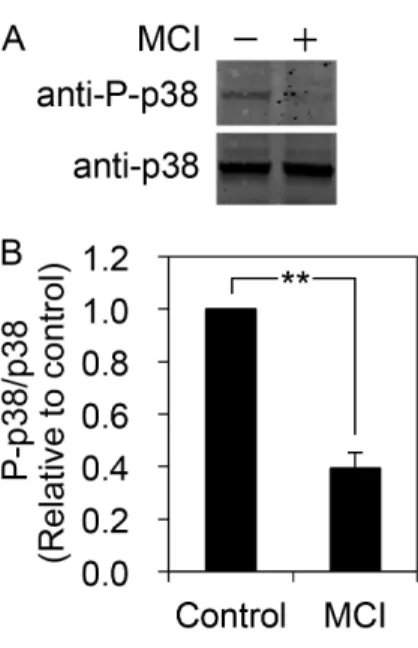
![Fig. 1.3 Effect of thapsigargin on [Ca 2+ ] i in submandibular gland cells](https://thumb-ap.123doks.com/thumbv2/123deta/6027131.2073921/14.892.165.725.111.753/fig-effect-thapsigargin-ca-i-submandibular-gland-cells.webp)
関連したドキュメント
The findings obtained in this study can be summarized as follows: i) CD56(-)CD16(+) NK cells were increased around the small bile ducts and beneath the biliary epithelia of large
Memory CD4+ T cells were able to respond to a pair of anti-CD2 mAbs (anti-T112 and anti-T113) to proliferate even in the absence of monocytes, whereas any proliferative re-
In immunostaining of cytokeratin using monoclonal antibodies, the gold particles were scattered in the cytoplasm of the hepatocytes and biliary epithelial cells
• Transplantation model systems were established in the zebrafish and clonal ginbuna carp to evaluate the activity of hematopoietic cells. • Hematopoietic stem cells
The present results also provide in situ hybridization evidence that at least chief cells and possibly both chief cells and sustentacular cells of the carotid body express NGF mRNA..
During land plant evolution, stem cells diverged in the gametophyte generation to form different types of body parts, including the protonema and rhizoid filaments, leafy-shoot
These findings further suggest that CD45 + /ColI + may contribute to kidney fibrosis by producing MCP-1/CCL2 and TGF-beta, which may be responsible for chronic
Precisely, over a period of 120 months, the total number of new infections that will be generated from the two patches in the absence of optimal control is 1.2037× 10 4 , whereas,