DOI: http://doi.org/10.14947/psychono.38.16
光遺伝学的手法を用いた触知覚とその記憶のメカニズムの解明
村 山 正 宜
理化学研究所 脳神経科学研究センサー 触知覚生理学研究チーム
Understanding the cortical mechanisms of somatosensory perception
and memory consolidation using optogenetics
Masanori Murayama
RIKEN Center for Brain Science, Lab for Haptic Perception and Cognitive Physiology
The neural circuits related to somatosensory perception, the flow of information between the circuits, and the neural activity which represents the content of perception still remains elusive. Recently, by using optogenetic meth-ods, we identified top-down circuits essential for tactile perception. Selective suppression of this circuit resulted in inaccurate tactile perception; mice cannot distinguish between smooth and rough floors. Moreover, we found that this circuit is active during sleep. Suppression of this circuit during non-rapid eye movement (non-REM) sleep blocked memory consolidation of tactile perception. This lecture introduces the outline of these findings.
光遺伝学的手法を用いた皮質間投射制御 ある脳機能は,一つの局所回路(A)の活動だけで表 出されるのか。それとも回路A–B間の相互活動によるの だろうか。仮に回路間活動に脳機能が依存するとした ら,投射 A→Bと,投射B→Aが運ぶ情報の中身や,そ の生理的役割は等しいのだろうか。この疑問を解決する ためには,各投射経路を制御し,その結果としての神経 活動と動物行動とを観察する必要がある。光遺伝学的手 法では,特定の細胞に光駆動型タンパク質を発現させ る。これにより,特定の細胞種を選択的に活性化するこ とが可能である。また発現させるタンパク質を選択すれ ば,細胞を抑制することも可能である。さらに,その細 胞の軸索を活性または抑制すれば,経路選択的な制御が 可能となる。本稿では,マウスの皮質間投射を選択的に 活性化または不活化して動物行動を制御した例を紹介す る。近年,我々の研究グループは,皮膚感覚の知覚(触 知覚)に関わる反響回路を発見した(Figure 1)(Manita et al., 2015; Manita, Miyakawa, Kitamura, & Murayama, 2017)。また我々は,この回路が触知覚の記憶の固定化
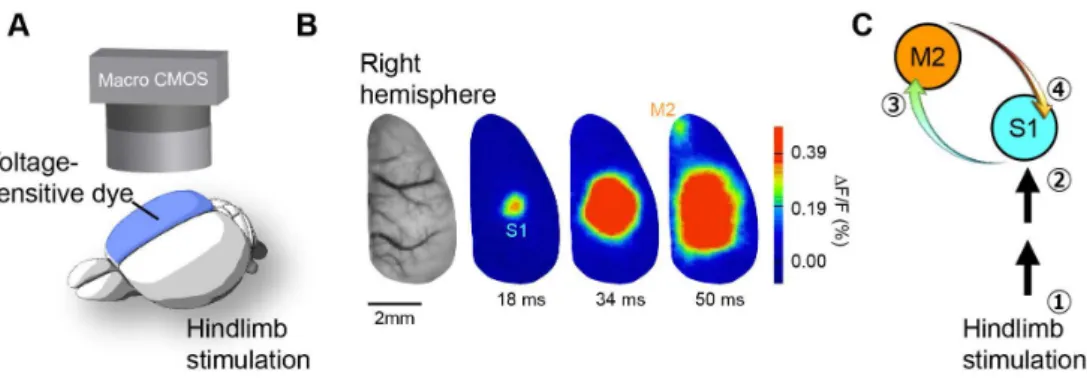
にも関与していることを見出している(Miyamoto et al., 2016; Miyamoto, Hirai, & Murayama, 2017)。これらの研究 において,我々は回路選択的な神経活動の制御を行いな がら動物の行動を観察した。このような介入実験と行動 観察から,その回路機能を明らかにすることが可能であ る。 触知覚に関わる皮質間投射 まず,マウスの皮膚感覚の知覚を司る神経回路を探る ために,膜電位イメージング法を用いて大脳新皮質の広 範な領域から神経活動を観測した(Figure 1A)。マウス の後肢を極微弱の電流で刺激すると,まず後肢に対応し た第一体性感覚野(S1)の領域が活性化し,次に第二運 動野(M2)が活性化した(Figure 1B)。続いて神経活動 を抑える薬をS1またはM2にそれぞれ投与し,その効果 を観測した。その結果,S1を抑制した場合はM2の活動 が,逆にM2を抑制した場合はS1の遅い活動(遅発性神 経活動)が抑制された。これらの結果は,後肢からの情 報がS1→M2→S1と流れることを示す(Figure 1C)。これ は,皮膚感覚がボトムアップ投射として S1から高次脳 領域であるM2に送られた後,再びS1へ「トップダウン 投射」としてフィードバックされる反響回路が存在する ことを示唆する。
Copyright 2019. The Japanese Psychonomic Society. All rights reserved. Corresponding address: RIKEN Center for Brain Science,
2–1 Hirosawa, Wako, Saitama 351–0198, Japan. E-mail: [email protected]
行動課題時におけるM2トップダウン投射の抑制 エネルギー消費の観点から,生体にとって無駄な神経 投射は存在しないと考えられる。では,マウスの触知覚 におけるトップダウン投射の生理的な役割は何か。これ を解明するため,光遺伝学的手法を用いて,M2神経細 胞が伸長する軸索を S1領域で不活性化させ,動物行動 を観 察 し た。 ま ず, 光 感 受 性 抑 制 性 イ オ ン ポ ン プ (ArchT)(Han et al., 2011)の遺伝子配列が組み込まれた アデノ随伴ウィルス(AAV)を両脳のM2に注入するこ とで,M2神経細胞にArchTを発現させた。その神経細 胞に光(緑色)を照射すると光感受性イオンポンプが活 性化し,細胞内から細胞外にH+が排出され,結果的に 神経細胞の活動は抑制される。そこで M2–S1経路を選 択的に抑制するため,M2神経細胞の投射先であるS1に て神経軸索を光抑制した(Figure 2A)。触知覚を評価す る目的で,マウスに対して皮膚感覚を手掛かりとする自 発性場所選好テストを行った(Figure 2B)。このテスト では,四角い箱の床面に紙やすり(ザラザラ)とそれを 裏返した面(ツルツル)を半分ずつ敷き,その箱の中に マウスを置いた。マウスの脳には小型光照射装置を設置 し,M2からS1へのトップダウン投射を光照射で抑制で きるようにした(Miyamoto & Murayama, 2016)。マウス はもともと,ザラザラ,ツルツルの床面のどちらか一方 を好む傾向があるため,光照射をしないマウス群では, ザラザラまたはツルツル床のどちらかに滞在時間が偏っ た。一方で,光照射をしたマウス群では,その偏りが減 少した(Figure 2C, D)。マウスの触知覚をテストする他 Figure 1. Identification of a reciprocal top-down control circuit.
A: Diagram showing macroscopic recording of cortical activity by using voltage-sensitive dye (VSD) imaging. B: Spatiotem-poral dynamics of VSD response evoked by hindpaw (HP) stimulation (single pulse, 0.1-ms duration, 100 V) C: Diagram showing information flow between primary somatosensory cortex (S1) and secondary motor cortex (M2).
Figure 2. Optogenetic inactivation of top-down input alters sensory perception.
A: Experimental diagram of a miniature wireless LED device attached to the S1 in the both hemispheres. AAV-ArchT or GFP were also injected to M2 in the both hemispheres. B: Diagram showing the mouse behavioral setup for the spontane-ous place preference test (SPPT). C: Place preference location plots from two representative mice showing the animal’s posi-tion over the course of the 4-min session with and without LED illuminaposi-tion. D: Summary of the preference in the SPPT Two-way ANOVA revealed a significant interaction effect of injection substrate (GFP or ArchT) and LED (on or off) (F(1, 27)=5.29, p<0.05) and a post-hoc test revealed a significant of LED effect on ArchT group (LED-off : 71.2±3.3% versus LED-on: 55.7±1.8%, F (1, 27)=12.94, * p<0.01)
の2課題でも同様に行動異常が観察された。一方,運動 機能や短期記憶,視覚機能や不安行動等をテストする行 動課題では成績はコントロール群と比べて優位な差は観 察されなかった。以上の結果から,M2からS1へのトッ プダウン投射が,正常な触知覚に必須であることがわ かった。 触知覚記憶の固定化を制御 知覚学習によりトップダウン方向の情報の連絡が強化 さ れ る こ と が霊 長 類(Miyashita, 2014) や げ っ 歯 類 (Makino & Komiyama, 2015)において知られており,記 憶を思い出す想起への関与が示唆されてきた。では,知
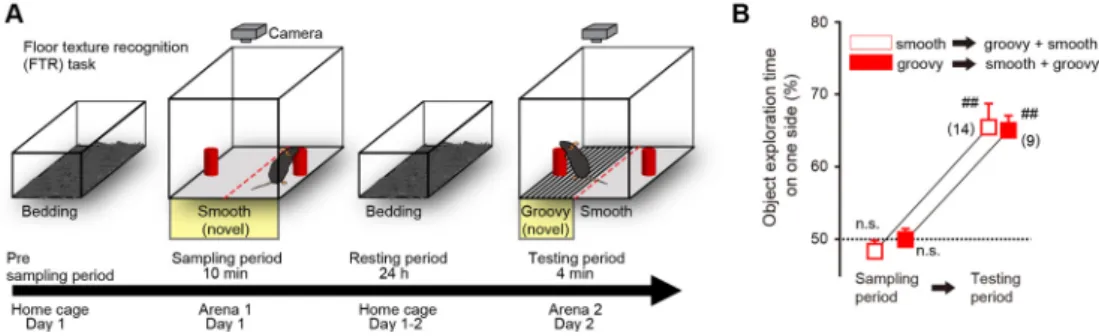
覚体験を想起の可能な記憶として固定化する過程におい てもトップダウン投射は関与するのか(Tononi & Cirelli, 2014)。逆に,S1 から M2 へのボトムアップ投射は記憶 の固定化に関与するのだろうか。そこで我々は,上述し たM2–S1回路をモデルとして,経路選択的な光遺伝学 的手法を用いてこの仮説を検証した。触覚に関する知覚 記憶の固定化を調べるためのマウスの行動課題として床 面認識課題を確立した(Figure 3A)。1日目,マウスを 10分間,床がツルツルの材質である箱の内部を自由に 探索させた。床の左右には探索用のオブジェクトが設置 してある。これを学習期間とする。そののち,マウスを ホームケージにもどし,24時間おく。これを休息期間 Figure 3. Floor texture recognition task.
A: Behavioral paradigm. During the sampling period, mice housed in a home cage were transferred to an arena with floor areas with a smooth- (or groovy-) -texture. After 10 min of free movement in the arena the mice were returned to the home cage for a 24 h resting period. Twenty-four hours after the sampling period, the mice were transferred to the arena with two textured floor areas for the testing period where they were permitted to move freely in the arena for 4 min. B: Object explo-ration time on a texture in the sampling period and on a novel texture in the testing period. Data are means±SEM; statisti-cal significance from 50% chance level (##p<0.01) was assessed by one-sample t-test.
Figure 4. Optogenetic inactivation of M2 axons impairs memory consolidation.
A: Examples of electroencephalography (EEG) and electromyography (EMG) recordings during the resting period. Brain states for awake, NREM, and REM sleep were identified with EEG recordings made through a screw in the skull and EMG re-cordings from a neck muscle. B: Diagram of the sleep-state specific optogenetics. Inactivation occurring during the resting period immediately after (first hour) or long after (6–7 h) the sample period was performed in a state-specific manner ac-cording to the detection system. The cumulative illumination time was 30 min in each state. Note that, in some mice, the end times of the illumination in NREM sleep were greater than the first hour because the cumulative 30 min of NREM sleep did not fall within the first 1 h. C: Summary for the task in (B) when M2 fibers were inactivated at S1 during the three periods.
とする。2日目,床の半分がツルツルの材質,半分がデ コボコの材質の箱を用意し,前日と同じオブジェクトを 設置して,マウスを自由に探索させた。これを試験期間 とする。すると,マウスはなじみのない新しい環境を好 んで探索する性質をもつので,ツルツルの床面に置かれ たオブジェクトよりも,はじめて体験するデコボコの床 面に置かれたオブジェクトをより長い時間にわたり探索 した(Figure 3B)。2日目にみられた,一方の質感の床面 への滞在時間のかたよりは,前日に経験した床面の質感 を記憶し,その記憶にもとづく新しい質感の床面への選 好性を示すと考えられるので,この偏りを知覚記憶の指 標として用いた。 マウスの脳状態は少なくとも3つに分類できる。覚醒 状態,深い眠りであるノンレム睡眠,浅い眠りであるレ ム睡眠,である(Figure 4A)。そこで,マウスの脳波お よび筋電位から脳の状態をリアルタイムで判定し,特定 の脳の状態にあるときだけ,M2からS1へのトップダウ ン投射を光遺伝学的な手法により抑制した(Figure 4B) (Tsunematsu et al., 2011)。学習の直後(0–1時間)のノン レム睡眠において光の照射によりトップダウン投射を抑 制すると知覚記憶の固定化は阻害されたが,学習の6∼ 7時間後のノンレム睡眠におけるトップダウン投射の抑 制は記憶の固定化に影響しなかった(Figure 4C)。一方, 学習の直後の覚醒においてトップダウン投射を抑制して も記憶の固定化には影響しなかった。これより,学習の 直後のノンレム睡眠におけるトップダウン投射が記憶の 固定化に必要であることがわかった。興味深いことに, 学習直後のノンレム睡眠時に,逆の経路,すなわちS1 からM2へのボトムアップ投射を同様の手法で抑制した が,この操作は記憶固定化を阻害しなかった。この結果 は,S1とM2は広域回路を形成しているにもかかわら ず,学習直後のノンレム睡眠時においては,S1からの 投射と,M2からの投射とでは,記憶の固定化に対する 生理的な役割が異なることを示す。 ノンレム睡眠時における皮質間神経活動 記憶の固定化がどのような神経活動によって担われて いるかを調べるために,M2とS1に電極を刺入し,各領 域の局所フィールド電位を記録した。局所フィールド電 位は,記録電極周囲の神経細胞集団全体の活動を捉えて おり,集団活動のオンオフのリズムと関連している。ノ ンレム睡眠時では,デルタ帯域(0.5–4 Hz)の低周波の 局所フィールド電位がM2とS1とで観察され,同期して いた。この 2領域での脳波をGranger因果性解析した。 Granger 因果性解析は,経時変化する 2 つの変数 X と Y (例: 気温とビールの消費量)において,変数X (例: 気 温)から変数Y (例: ビールの消費量)への予測を試み る解析方法である(Brovelli et al., 2004)。解析の結果,実 際にM2からS1へのトップダウン方向に情報が流れてい ることを見出した。光照射でM2からS1へのトップダウ ン投射を抑制すると,この情報の流れは抑制された。 次に,それぞれの領域でのユニット記録により単一神 経細胞レベルの神経活動を記録した。M2とS1の両方に おいて,学習時に活性化した神経細胞は,学習後のノン レム睡眠時においても再活性化していた。光照射によっ てM2からS1へのトップダウン投射を抑制すると,M2に おける再活性化は阻害されなかったが,S1における再活 性化が阻害された。これまでの研究で,記憶の固定化に は神経活動の再活性化が重要だと考えられてきたが (Yang et al., 2014),実際にどの脳領域が再活性化を誘発し ているのか,全くわかっていなかった。これらの結果は, 感覚記憶の固定化に不可欠な感覚野における再活性化に は,皮質間トップダウン投射が必要であることを示す。 2領域の同期/非同期活性による記憶固定化の制御 次に,トップダウン投射の活性化が記憶の固定化にお よぼす影響について検討した。大脳新皮質を刺激するた め,大脳新皮質の興奮性神経細胞にチャネルロドプシン (ChR2)を発現した遺伝子改変マウスを用いた(Beltra-mo et al., 2013)。光感受性のイオンチャネルであるチャ ネルロドプシンを発現した神経細胞に光を照射すると, 神経細胞の活動は活性化される。そこで学習ののちのノ ンレム睡眠において,M2およびS1をデルタ帯域(2 Hz) のリズムで同期させた光を用いて刺激した。光刺激して いない通常のマウスにおいては,学習の4日後に床面認 識課題を行うと,1日目の知覚記憶を忘れており床面の 選好性を示さなかった。ところが,同期して光刺激した マウスは学習から4日たっても記憶を保持していた。一 方で,光を同期させずに位相をずらして光刺激すると, 翌日には記憶を忘れていた。この結果から,大脳新皮質 に対する特定の刺激により記憶の固定化を制御すること が可能であることが示された。 通常,断眠されたマウスは正常な知覚記憶を形成でき ない。そこで,断眠されたマウスにおいても同期した光 刺激により記憶の固定化が可能かどうかを調べた。断眠 させたマウスのM2とS1に同期光刺激を与えた場合,こ のマウスは学習から1日後においても4日後においても 知覚記憶を保持していた。このことから,睡眠不足の場 合でも,大脳新皮質を適切に刺激すれば記憶を維持ある いは向上できることがわかった。
む す び 光遺伝学的手法を用いた投射経路選択的な活動制御 は,生理的意義が不明な回路の役割を探究する研究に役 立つ。これまでの回路研究は,ある脳部位の神経活動と 脳機能との関連を観察するだけの相関関係探求型の研究 であった。因果関係探求型の研究には,直接,神経活動 に介入することが可能な光学遺伝学的手法による投射選 択的な制御が有用である。今回はスペースの関係上割愛 したが,化学遺伝学的手法(DREADD, Designer Recep-tors Exclusively Activated by Designer Drugs)も同様に強力 な研究ツールである。この手法は,光遺伝学的手法に比 べると回路制御の時間精度は落ちるが,2光子顕微鏡な どを用いた光学測定法との相性がよく,生体内で電気記 録が極めて困難な樹状突起活動や軸索活動を光学的に捉 えながら,シナプス前細胞またはそれらの活動を制御で きる点は特筆すべきである。研究目的と手法に合わせた 光・化学遺伝学的手法の組み合わせにより生理学的な広 域回路研究が今後ますます加速するだろう。 引用文献
Beltramo, R., D’Urso, G., Dal Maschio, M., Farisello, P., Bovet-ti, S., Clovis, Y., ... Fellin, T. (2013). Layer-specific excitatory circuits differentially control recurrent network dynamics in the neocortex. Nature Neuroscience, 16, 227–234.
Brovelli, A., Ding, M., Ledberg, A., Chen, Y., Nakamura, R., & Bressler, S. L. (2004). Beta oscillations in a large-scale senso-rimotor cortical network: Directional influences revealed by Granger causality. Proceeding of the National Academy of
Sciences of the United States of America, 101, 9849–9854.
Han., X., Chow, B. Y., Zhou, H., Klapoetke, N. C., Chuong, A., Rajimehr, R., ... Boyden, E. S. (2011). A high-light
sensitivi-ty optical neural silencer: Development and application to optogenetic control of non-human primate cortex. Frontiers
in Systems Neuroscience, 5, 18.
Makino, H., & Komiyama, T. (2015). Learning enhances the relative impact of top-down processing in the visual cortex.
Nature Neuroscience, 18, 1116–1122.
Manita, S., Suzuki, T., Homma, C., Matsumoto, T., Odagawa, M., Yamada, K., ... Murayama, M. (2015). A top-down corti-cal circuit for accurate sensory perception. Neuron, 86, 1304–1316.
Manita, S., Miyakawa, H., Kitamura, K., & Murayama, M. (2017). Dendritic spikes in sensory perception. Frontiers in
Cellular Neuroscience, 11, 29.
Miyamoto., D., Hirai, D., Fung, C. C. A., Inutsuka, A., Odaga-wa, M., Suzuki, T., ... Murayama, M. (2016). Top-down cor-tical input during NREM sleep consolidates perceptual memory. Science, 352, 1315–1318.
Miyamoto, D., Hirai, D., & Murayama, M. (2017). The roles of cortical slow waves in synaptic plasticity and memory con-solidation. Frontiers in Neural Circuits, 11, 92.
Miyamoto, D., & Murayama, M. (2016). The fiber-optic imag-ing and manipulation of neural activity durimag-ing animal be-havior. Neuroscience Research, 103, 1–109.
Miyashita., Y. (2014). Cognitive memory: Cellular and net-work machineries and their top-down control. Science, 306, 435–440.
Tononi, G., & Cirelli, C. (2014). Sleep and the price of plastici-ty: from synaptic and cellular homeostasis to memory con-solidation and integration. Neuron, 81, 12–34.
Tsunematsu, T., Kilduff, T. S., Boyden, E. S., Takahashi, S., Tominaga, M., & Yamanaka, A. (2011). Acute optogenetic silencing of orexin/hypocretin neurons induces slow-wave sleep in mice. Journal of Neuroscience, 31, 10529–10539. Yang, G., Lai, C. S., Cichon, J., Ma, L., Li, W., & Gan, W. B.
(2014). Sleep promotes branch-specific formation of den-dritic spines after learning. Science, 344, 1173–1178.